
Assessing the Involvement of Selected Phenotypes of Pseudomonas simiae PICF7 in Olive Root Colonization and Biological Control of Verticillium dahliae


 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Comparative Genomics Supports the Taxonomic Assignment of Pseudomonas simiae PICF7
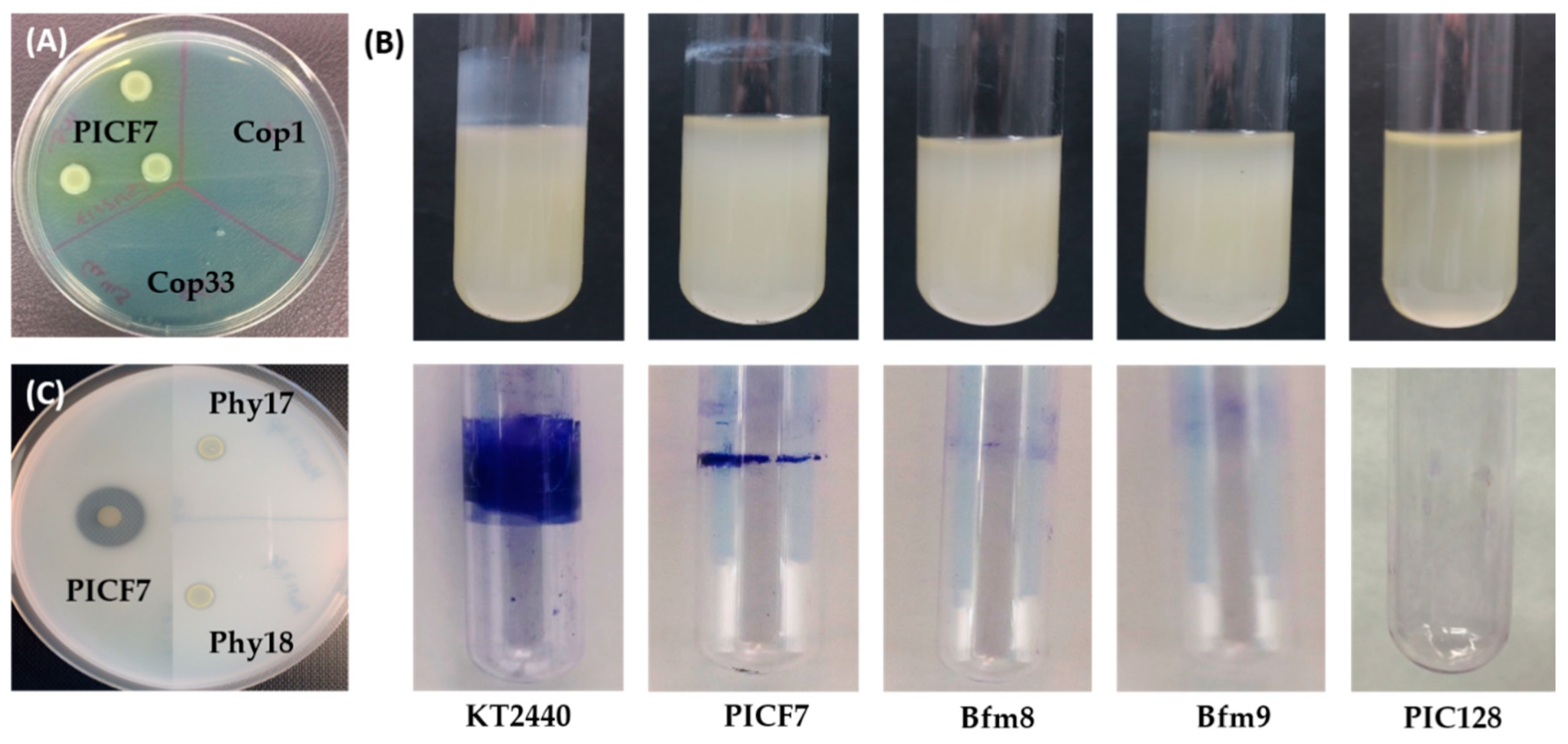
2.2. Screening of a PICF7 Mutant Bank to Identify Altered Target Phenotypes
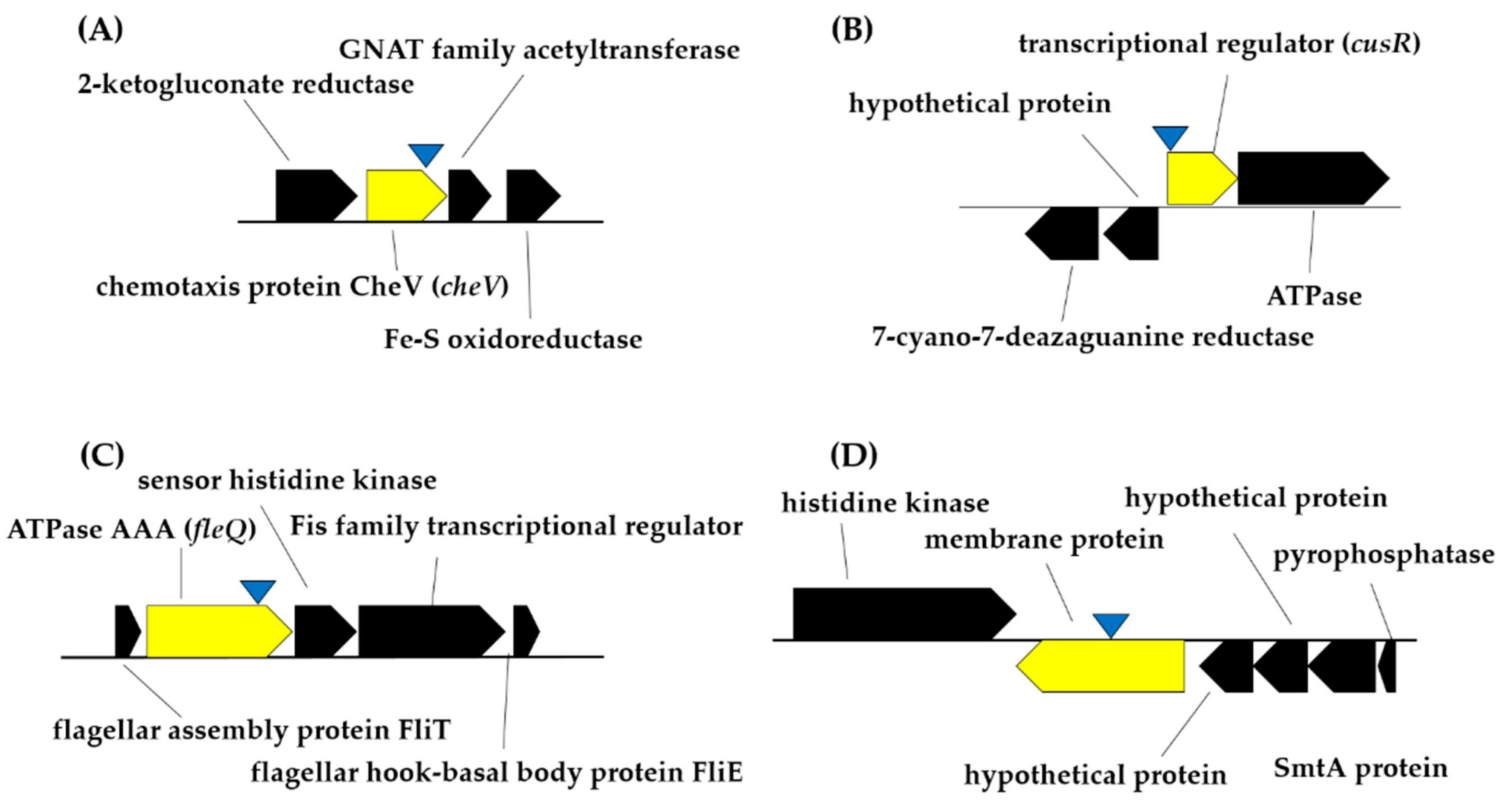
2.3. Identification of the Disrupted Gene in the Genomes of Selected Mutants of Pseudomonas simiae PICF7
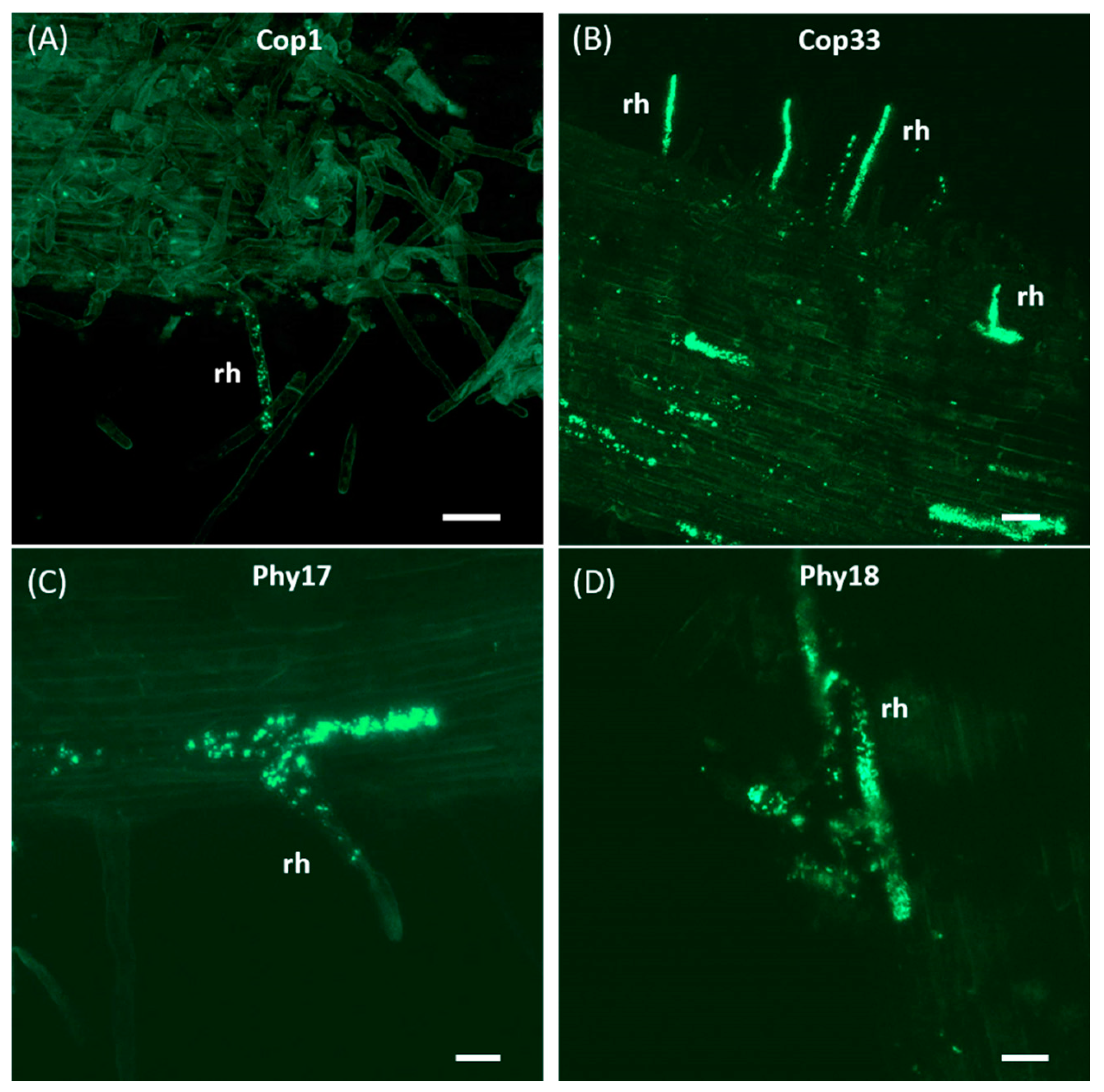
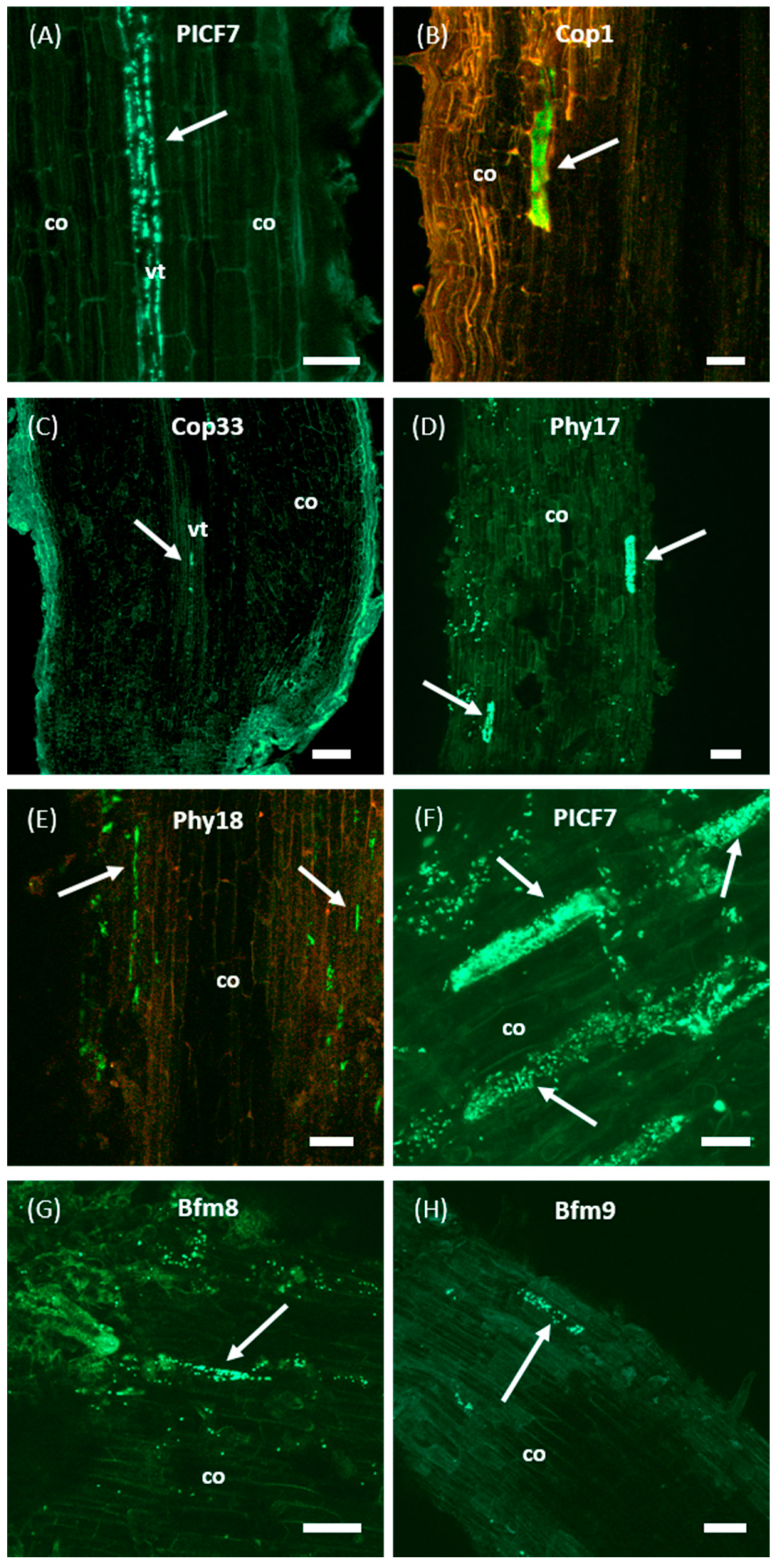
2.4. Only PICF7 Mutants Impaired in Biofilm Formation Are Unable to Internally Colonize Olive Roots
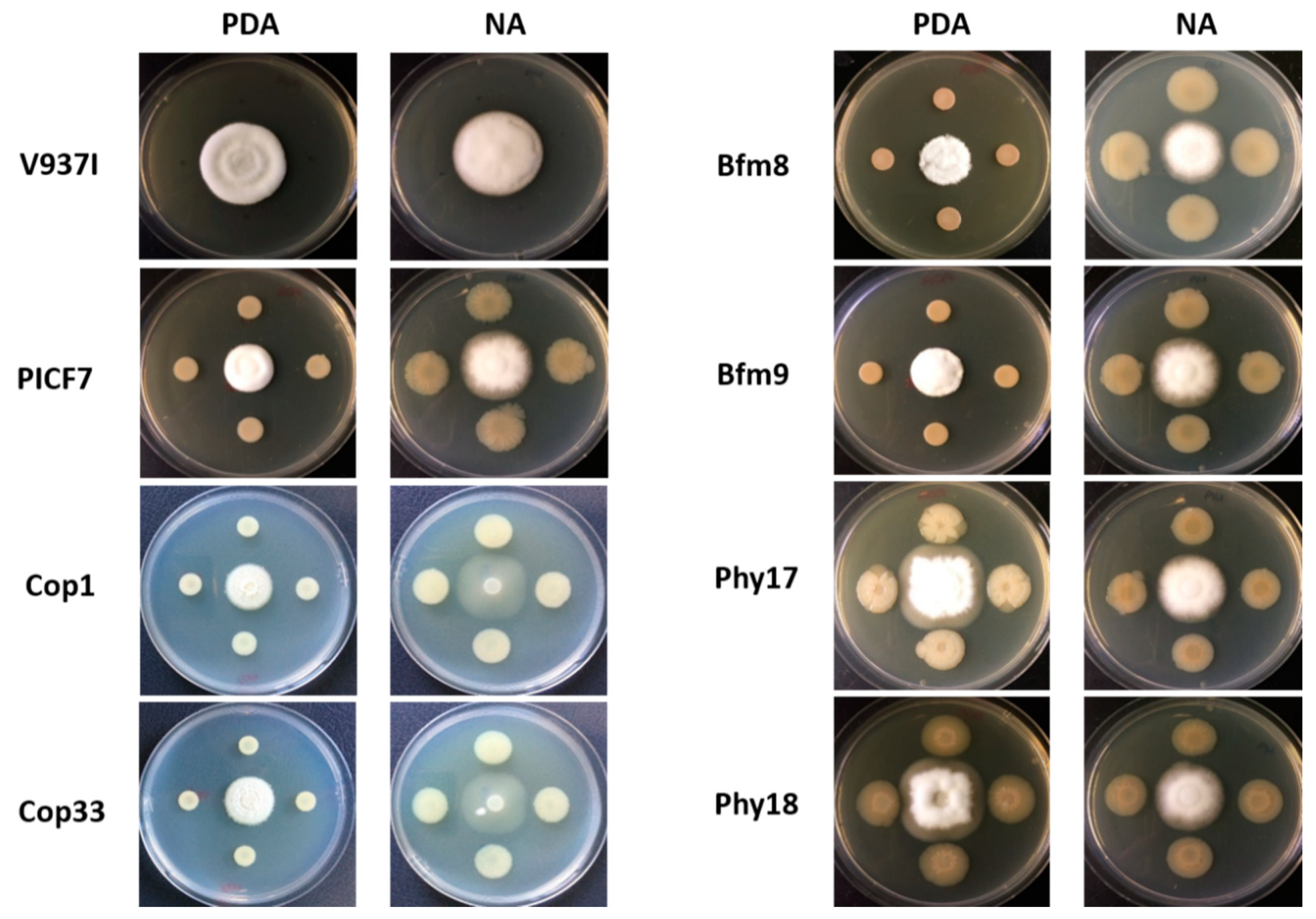
2.5. PICF7 Mutants Defective in Phytase Activity Show a Significant Reduction of In Vitro Antagonism against Verticillium dahliae
2.6. PICF7 Mutants Are Not Affected in Biological Control Ability against Verticillium Wilt of Olive
3. Discussion
4. Materials and Methods
4.1. Culturing Media and Conditions, Microorganisms and Plasmid
4.2. Comparative Analysis of Pseudomonas simiae Genomes
4.3. Screening of Pseudomonas simiae PICF7 Mutants
4.3.1. Identification and Selection of Mutants Impaired in Copper Tolerance
4.3.2. Identification and Selection of Mutants Impaired in Biofilm Formation
4.3.3. Identification and Selection of Mutants Defective in Phytase Activity
4.4. Identification of Transposon Insertion Sites in Selected PICF7 Mutants
4.5. Evaluation of the Olive Root Colonization Ability by Pseudomonas simiae PICF7 Mutants
4.6. In Vitro Antagonism Assays
4.7. Biocontrol Effectiveness of Pseudomonas simiae PICF7 Mutants against Verticillium Wilt of Olive
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- International Olive Council. Available online: www.internationaloliveoil.org/ (accessed on 17 February 2020).
- Montes-Osuna, N.; Mercado-Blanco, J. Verticillium Wilt of Olive and its Control: What Did We Learn during the Last Decade? Plants 2020, 9, 735. [Google Scholar] [CrossRef] [PubMed]
- López-Escudero, F.J.; Mercado-Blanco, J. Verticillium wilt of olive: A case study to implement an integrated strategy to control a soil-borne pathogen. Plant Soil 2011, 344, 1–50. [Google Scholar] [CrossRef] [Green Version]
- Bizos, G.; Papatheodorou, E.M.; Chatzistathis, T.; Ntalli, N.; Aschonitis, V.G.; Monokrousos, N. The Role of Microbial Inoculants on Plant Protection, Growth Stimulation, and Crop Productivity of the Olive Tree (Olea europea L.). Plants 2020, 9, 743. [Google Scholar] [CrossRef]
- Deketelaere, S.; Tyvaert, L.; França, S.D.C.; Höfte, M. Desirable Traits of a Good Biocontrol Agent against Verticillium Wilt. Front. Microbiol. 2017, 8, 1186. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Lama Cabanás, C.; Legarda, G.; Ruano-Rosa, D.; Pizarro-Tobías, P.; Valverde-Corredor, A.; Niqui, J.L.; Triviño, J.C.; Roca, A.; Mercado-Blanco, J. Indigenous Pseudomonas spp. Strains from the Olive (Olea europaea L.) Rhizosphere as Effective Biocontrol Agents against Verticillium dahliae: From the Host Roots to the Bacterial Genomes. Front. Microbiol. 2018, 9, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Lama Cabanás, C.; Ruano-Rosa, D.; Legarda, G.; Pizarro-Tobías, P.; Valverde-Corredor, A.; Triviño, J.C.; Roca, A.; Mercado-Blanco, J. Bacillales Members from the Olive Rhizosphere Are Effective Biological Control Agents against the Defoliating Pathotype of Verticillium dahliae. Agriculture 2018, 8, 90. [Google Scholar] [CrossRef] [Green Version]
- Mulero-Aparicio, A.; Agustí-Brisach, C.; Varo, Á.; López-Escudero, F.J.; Trapero, A. A non-pathogenic strain of Fusarium oxysporum as a potential biocontrol agent against Verticillium wilt of olive. Biol. Control. 2019, 139, 104045. [Google Scholar] [CrossRef]
- Lozano-Tovar, M.; Garrido-Jurado, I.; Quesada-Moraga, E.; Raya-Ortega, M.; Trapero-Casas, A. Metarhizium brunneum and Beauveria bassiana release secondary metabolites with antagonistic activity against Verticillium dahliae and Phytophthora megasperma olive pathogens. Crop. Prot. 2017, 100, 186–195. [Google Scholar] [CrossRef]
- Boutaj, H.; Chakhchar, A.; Meddich, A.; Wahbi, S.; El Alaoui-Talibi, Z.; Douira, A.; Filali-Maltouf, A.; El Modafar, C. Bioprotection of olive tree from Verticillium wilt by autochthonous endomycorrhizal fungi. J. Plant Dis. Prot. 2020, 127, 349–357. [Google Scholar] [CrossRef]
- Aranda, S.; Montes-Borrego, M.; Jiménez-Díaz, R.M.; Landa, B.B. Microbial communities associated with the root system of wild olives (Olea europaea L. subsp. europaea var. sylvestris) are good reservoirs of bacteria with antagonistic potential against Verticillium dahliae. Plant Soil 2011, 343, 329–345. [Google Scholar] [CrossRef] [Green Version]
- Emueller, H.; Eberg, C.; Landa, B.B.; Eauerbach, A.; Emoissl-Eichinger, C.; Eberg, G. Plant genotype-specific archaeal and bacterial endophytes but similar Bacillus antagonists colonize Mediterranean olive trees. Front. Microbiol. 2015, 6, 138. [Google Scholar] [CrossRef] [Green Version]
- Fernández-González, A.J.; Cardoni, M.; Gómez-Lama Cabanás, C.; Valverde-Corredor, A.; Villadas, P.J.; Fernández-López, M.; Mercado-Blanco, J. Linking belowground microbial network changes to different tolerance level towards Verticillium wilt of olive. Microbiome 2020, 8, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Martínez-García, P.M.; Ruano-Rosa, D.; Schilirò, E.; Prieto, P.; Ramos, C.; Rodríguez-Palenzuela, P.; Mercado-Blanco, J. Complete genome sequence of Pseudomonas fluorescens strain PICF7, an indigenous root endophyte from olive (Olea europaea L.) and effective biocontrol agent against Verticillium dahliae. Stand. Genom. Sci. 2015, 10, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercado-Blanco, J.; Rodrı́guez-Jurado, D.; Hervás, A.B.; Jiménez-Dı́az, R.M. Suppression of Verticillium wilt in olive planting stocks by root-associated fluorescent Pseudomonas spp. Biol. Control. 2004, 30, 474–486. [Google Scholar] [CrossRef]
- Prieto, P.; Mercado-Blanco, J. Endophytic colonization of olive roots by the biocontrol strain Pseudomonas fluorescens PICF7. FEMS Microbiol. Ecol. 2008, 64, 297–306. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Lama Cabanás, C.; Schilirò, E.; Valverde-Corredor, A.; Mercado-Blanco, J. The biocontrol endophytic bacterium Pseudomonas fluorescens PICF7 induces systemic defense responses in aerial tissues upon colonization of olive roots. Front. Microbiol. 2014, 5, 427. [Google Scholar] [CrossRef]
- Gómez-Lama Cabanás, C.; Sesmero, R.; Valverde-Corredor, A.; López-Escudero, F.J.; Mercado-Blanco, J. A split-root system to assess biocontrol effectiveness and defense-related genetic responses in above-ground tissues during the tripartite interaction Verticillium dahliae-olive-Pseudomonas fluorescens PICF7 in roots. Plant Soil 2017, 417, 433–452. [Google Scholar] [CrossRef]
- Maldonado-González, M.M.; Schilirò, E.; Prieto, P.; Mercado-Blanco, J. Endophytic colonization and biocontrol performance of Pseudomonas fluorescens PICF7 in olive (Olea europaea L.) are determined neither by pyoverdine production nor swimming motility. Environ. Microbiol. 2014, 17, 3139–3153. [Google Scholar] [CrossRef] [PubMed]
- Maldonado-González, M.M.; Bakker, P.A.H.M.; Prieto, P.; Mercado-Blanco, J. Arabidopsis thaliana as a tool to identify traits involved in Verticillium dahliae biocontrol by the olive root endophyte Pseudomonas fluorescens PICF7. Front. Microbiol. 2015, 6, 266. [Google Scholar] [CrossRef]
- Maldonado-González, M.M.; Prieto, P.; Ramos, C.; Mercado-Blanco, J. From the root to the stem: Interaction between the biocontrol root endophyte Pseudomonas fluorescens PICF7 and the pathogen Pseudomonas savastanoi NCPPB 3335 in olive knots. Microb. Biotechnol. 2013, 6, 275–287. [Google Scholar] [CrossRef] [Green Version]
- Schilirò, E.; Ferrara, M.; Nigro, F.; Mercado-Blanco, J. Genetic Responses Induced in Olive Roots upon Colonization by the Biocontrol Endophytic Bacterium Pseudomonas fluorescens PICF7. PLoS ONE 2012, 7, e48646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Outten, F.W.; Huffman, D.L.; Hale, J.A.; O’Halloran, T.V. The Independent cue and cusSystems Confer Copper Tolerance during Aerobic and Anaerobic Growth in Escherichia coli. J. Biol. Chem. 2001, 276, 30670–30677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rensing, C.; Franke, S. Copper Homeostasis in Escherichia coli and Other Enterobacteriaceae. EcoSal Plus 2007, 2. [Google Scholar] [CrossRef]
- Castro, D.; Torres, M.; Sampedro, I.; Martínez-Checa, F.; Torres, B.; Béjar, V. Biological Control of Verticillium Wilt on Olive Trees by the Salt-Tolerant Strain Bacillus velezensis XT1. Microorganisms 2020, 8, 1080. [Google Scholar] [CrossRef]
- Chong, T.M.; Yin, W.-F.; Chen, J.-W.; Mondy, S.; Grandclément, C.; Faure, D.; Dessaux, Y.; Chan, K.-G. Comprehensive genomic and phenotypic metal resistance profile of Pseudomonas putida strain S13.1.2 isolated from a vineyard soil. AMB Express 2016, 6, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Mercado-Blanco, J.; Alós, E.; Rey, M.D.; Prieto, P. Pseudomonas fluorescensPICF7 displays an endophytic lifestyle in cultivated cereals and enhances yield in barley. FEMS Microbiol. Ecol. 2016, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Lama Cabanás, C.; Mercado-Blanco, J. What determines successful colonization and expression of biocontrol traits at the belowground level? In How Research can Stimulate the Development of Commercial Biological Control against Plant Diseases; De Cal, A., Melgarejo, P., Magan, N., Eds.; Progress in Biological Control; Springer International Publishing: Berlin/Heidelberg, Germany, 2020; pp. 31–46. ISBN 978-3-030-53238-3. [Google Scholar]
- Singh, P.; Kumar, V.; Agrawal, S. Evaluation of Phytase Producing Bacteria for Their Plant Growth Promoting Activities. Int. J. Microbiol. 2014, 2014, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Chen, R.; Zuo, Z.; Lv, Z.; Yang, Z.; Mao, W.; Liu, Y.; Zhou, Y.; Huang, J.; Song, Z. Evaluation and improvement of phosphate solubilization by an isolated bacterium Pantoea agglomerans ZB. World J. Microbiol. Biotechnol. 2020, 36, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Alori, E.T.; Glick, B.R.; Babalola, O.O. Microbial Phosphorus Solubilization and Its Potential for Use in Sustainable Agriculture. Front. Microbiol. 2017, 8, 971. [Google Scholar] [CrossRef] [Green Version]
- Compant, S.; Clément, C.; Sessitsch, A. Plant growth-promoting bacteria in the rhizo- and endosphere of plants: Their role, colonization, mechanisms involved and prospects for utilization. Soil Biol. Biochem. 2010, 42, 669–678. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, A.; Schikora, A. Quorum Sensing of Bacteria and Trans-Kingdom Interactions of N-Acyl Homoserine Lactones with Eukaryotes. J. Chem. Ecol. 2012, 38, 704–713. [Google Scholar] [CrossRef] [PubMed]
- Mercado-Blanco, J.; Bakker, P.A.H.M. Interactions between plants and beneficial Pseudomonas spp.: Exploiting bacterial traits for crop protection. Antonie van Leeuwenhoek 2007, 92, 367–389. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Zhang, H.; Sun, X.; Liu, Y.; Yan, W.; Xun, W.; Shen, Q.; Zhang, R. Bacillus velezensis Wall Teichoic Acids Are Required for Biofilm Formation and Root Colonization. Appl. Environ. Microbiol. 2018, 85, 02116–02118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandin, C.; Le Coq, D.; Canette, A.; Aymerich, S.; Briandet, R. Should the biofilm mode of life be taken into consideration for microbial biocontrol agents? Microb. Biotechnol. 2017, 10, 719–734. [Google Scholar] [CrossRef] [Green Version]
- Tovi, N.; Frenk, S.; Hadar, Y.; Minz, D. Host Specificity and Spatial Distribution Preference of Three Pseudomonas Isolates. Front. Microbiol. 2019, 9, 3263. [Google Scholar] [CrossRef]
- Calderón, C.E.; Tienda, S.; Heredia-Ponce, Z.; Arrebola, E.; Cárcamo-Oyarce, G.; Eberl, L.; Cazorla, F.M. The Compound 2-Hexyl, 5-Propyl Resorcinol Has a Key Role in Biofilm Formation by the Biocontrol Rhizobacterium Pseudomonas chlororaphis PCL1606. Front. Microbiol. 2019, 10, 396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colvin, K.M.; Alnabelseya, N.; Baker, P.; Whitney, J.C.; Howell, P.L.; Parsek, M.R. PelA Deacetylase Activity Is Required for Pel Polysaccharide Synthesis in Pseudomonas aeruginosa. J. Bacteriol. 2013, 195, 2329–2339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pham, T.H.; Webb, J.S.; Rehm, B.H.A. The role of polyhydroxyalkanoate biosynthesis by Pseudomonas aeruginosa in rhamnolipid and alginate production as well as stress tolerance and biofilm formation. Microbiology 2004, 150, 3405–3413. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wilks, J.C.; Danhorn, T.; Ramos, I.; Croal, L.; Newman, D.K. Phenazine-1-Carboxylic Acid Promotes Bacterial Biofilm Development via Ferrous Iron Acquisition. J. Bacteriol. 2011, 193, 3606–3617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velmourougane, K.; Prasanna, R.; Saxena, A.K. Agriculturally important microbial biofilms: Present status and future prospects. J. Basic Microbiol. 2017, 57, 548–573. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Van Verk, M.C.; Stringlis, I.A.; Zamioudis, C.; Tommassen, J.; Pieterse, C.M.J.; Bakker, P.A.H.M. Unearthing the genomes of plant-beneficial Pseudomonas model strains WCS358, WCS374 and WCS417. BMC Genom. 2015, 16, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Vela, A.I.; Gutiérrez, M.C.; Falsen, E.; Rollán, E.; Simarro, I.; García, P.; Domínguez, L.; Ventosa, A.; Fernández-Garayzábal, J.F. Pseudomonas simiae sp. nov., isolated from clinical specimens from monkeys (Callithrix geoffroyi). Int. J. Syst. Evol. Microbiol. 2006, 56, 2671–2676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prieto, P.; Schilirò, E.; Maldonado-González, M.M.; Valderrama, R.; Barroso-Albarracín, J.B.; Mercado-Blanco, J. Root Hairs Play a Key Role in the Endophytic Colonization of Olive Roots by Pseudomonas spp. with Biocontrol Activity. Microb. Ecol. 2011, 62, 435–445. [Google Scholar] [CrossRef] [Green Version]
- Eljounaidi, K.; Lee, S.K.; Bae, H. Bacterial endophytes as potential biocontrol agents of vascular wilt diseases—Review and future prospects. Biol. Control. 2016, 103, 62–68. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Berendsen, R.L.; De Jonge, R.; Stringlis, I.A.; Van Dijken, A.J.H.; Van Pelt, J.A.; Van Wees, S.C.M.; Yu, K.; Zamioudis, C.; Bakker, P.A.H.M. Pseudomonas simiae WCS417: Star track of a model beneficial rhizobacterium. Plant Soil 2020, 1–19. [Google Scholar] [CrossRef]
- Khoshru, B.; Mitra, D.; Khoshmanzar, E.; Myo, E.M.; Uniyal, N.; Mahakur, B.; Das Mohapatra, P.K.; Panneerselvam, P.; Boutaj, H.; Alizadeh, M.; et al. Current scenario and future prospects of plant growth-promoting rhizobacteria: An economic valuable resource for the agriculture revival under stressful conditions. J. Plant Nutr. 2020, 43, 3062–3092. [Google Scholar] [CrossRef]
- Shulse, C.N.; Chovatia, M.; Agosto, C.; Wang, G.; Hamilton, M.; Deutsch, S.; Yoshikuni, Y.; Blow, M.J. Engineered Root Bacteria Release Plant-Available Phosphate from Phytate. Appl. Environ. Microbiol. 2019, 85, e01210-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanif, M.K.; Hameed, S.; Imran, A.; Naqqash, T.; Shahid, M.; Van Elsas, J.D. Isolation and characterization of a β-propeller gene containing phosphobacterium Bacillus subtilis strain KPS-11 for growth promotion of potato (Solanum tuberosum L.). Front. Microbiol. 2015, 6, 583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Illakkiam, D.; Shankar, M.; Ponraj, P.; Rajendhran, J.; Gunasekaran, P. Genome Sequencing of a Mung Bean Plant Growth Promoting Strain of P. aeruginosa with Biocontrol Ability. Int. J. Genom. 2014, 2014, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Jog, R.; Pandya, M.; Nareshkumar, G.; Rajkumar, S. Mechanism of phosphate solubilization and antifungal activity of Streptomyces spp. isolated from wheat roots and rhizosphere and their application in improving plant growth. Microbiology 2014, 160, 778–788. [Google Scholar] [CrossRef] [Green Version]
- Trivedi, P.; Pandey, A.; Palni, L.M.S. In vitro evaluation of antagonistic properties of Pseudomonas corrugata. Microbiol. Res. 2008, 163, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yan, F.; Chai, Y.; Liu, H.; Kolter, R.; Losick, R.; Guo, J.-H. Biocontrol of tomato wilt disease by Bacillus subtilis isolates from natural environments depends on conserved genes mediating biofilm formation. Environ. Microbiol. 2013, 15, 848–864. [Google Scholar] [CrossRef] [Green Version]
- Sang, M.K.; Kim, K.D. Biocontrol activity and root colonization by Pseudomonas corrugata strains CCR04 and CCR80 against Phytophthora blight of pepper. BioControl 2014, 59, 437–448. [Google Scholar] [CrossRef]
- Fan, H.; Zhang, Z.; Li, Y.; Zhang, X.; Duan, Y.; Wang, Q. Biocontrol of Bacterial Fruit Blotch by Bacillus subtilis 9407 via Surfactin-Mediated Antibacterial Activity and Colonization. Front. Microbiol. 2017, 8, 1973. [Google Scholar] [CrossRef]
- Triveni, S.; Prasanna, R.; Kumar, A.; Bidyarani, N.; Singh, R.; Saxena, A.K. Evaluating the promise of Trichoderma and Anabaena based biofilms as multifunctional agents in Macrophomina phaseolina- infected cotton crop. Biocontrol Sci. Technol. 2015, 25, 656–670. [Google Scholar] [CrossRef]
- Prieto, P.; Navarro-Raya, C.; Valverde-Corredor, A.; Amyotte, S.G.; Dobinson, K.F.; Mercado-Blanco, J. Colonization process of olive tissues by Verticillium dahliae and its in planta interaction with the biocontrol root endophyte Pseudomonas fluorescens PICF7. Microb. Biotechnol. 2009, 2, 499–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colvin, K.M.; Irie, Y.; Tart, C.S.; Urbano, R.; Whitney, J.C.; Ryder, C.; Howell, P.L.; Wozniak, D.J.; Parsek, M.R. The Pel and Psl polysaccharides provide Pseudomonas aeruginosa structural redundancy within the biofilm matrix. Environ. Microbiol. 2012, 14, 1913–1928. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, L.; Li, X.; Halverson, L.J. Cell-cell and cell-surface interactions mediated by cellulose and a novel exopolysaccharide contribute to Pseudomonas putida biofilm formation and fitness under water-limiting conditions. Environ. Microbiol. 2011, 13, 1342–1356. [Google Scholar] [CrossRef]
- Porbaran, M.; Habibipour, R. Relationship between Biofilm Regulating Operons and Various Β-Lactamase Enzymes: Analysis of the Clinical Features of Infections caused by Non-Fermentative Gram-Negative Bacilli (Nfgnb) from Iran. J. Pure Appl. Microbiol. 2020, 14, 1723–1736. [Google Scholar] [CrossRef]
- Rybtke, M.; Berthelsen, J.; Yang, L.; Høiby, N.; Givskov, M.; Tolker-Nielsen, T. The LapG protein plays a role in Pseudomonas aeruginosa biofilm formation by controlling the presence of the CdrA adhesin on the cell surface. Microbiology 2015, 4, 917–930. [Google Scholar] [CrossRef] [Green Version]
- Blanco-Romero, E.; Redondo-Nieto, M.; Martínez-Granero, F.; Garrido-Sanz, D.; Ramos-González, M.I.; Martín, M.; Rivilla, R. Genome-wide analysis of the FleQ direct regulon in Pseudomonas fluorescens F113 and Pseudomonas putida KT2440. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Nogales, J.; Vargas, P.; Farias, G.A.; Olmedilla, A.; Sanjuán, J.; Gallegos, M.-T. FleQ Coordinates Flagellum-Dependent and -Independent Motilities in Pseudomonas syringae pv. tomato DC3000. Appl. Environ. Microbiol. 2015, 81, 7533–7545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiménez-Fernández, A.; López-Sánchez, A.; Jiménez-Díaz, L.; Navarrete, B.; Calero, P.; Platero, A.I.; Govantes, F. Complex Interplay between FleQ, Cyclic Diguanylate and Multiple σ Factors Coordinately Regulates Flagellar Motility and Biofilm Development in Pseudomonas putida. PLoS ONE 2016, 11, e0163142. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Nie, H.; Liu, H.; Luo, X.; Chen, W.; Huang, Q. C-di-GMP regulates the expression of lapA and bcs operons via FleQ in Pseudomonas putida KT2440. Environ. Microbiol. Rep. 2016, 8, 659–666. [Google Scholar] [CrossRef]
- Tomich, M.; Planet, P.J.; Figurski, D.H. The tad locus: Postcards from the widespread colonization island. Nat. Rev. Genet. 2007, 5, 363–375. [Google Scholar] [CrossRef] [PubMed]
- Eangelov, A.; Ebergen, P.; Enadler, F.; Ehornburg, P.; Elichev, A.; Ãœbelacker, M.; Epachl, F.; Ekuster, B.; Eliebl, W. Novel Flp pilus biogenesis-dependent natural transformation. Front. Microbiol. 2015, 6, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanco-Romero, E.; Garrido-Sanz, D.; Rivilla, R.; Redondo-Nieto, M.; Martín, M. In Silico Characterization and Phylogenetic Distribution of Extracellular Matrix Components in the Model Rhizobacteria Pseudomonas fluorescens F113 and Other Pseudomonads. Microorganisms 2020, 8, 1740. [Google Scholar] [CrossRef] [PubMed]
- Bernard, C.S.; Bordi, C.; Termine, E.; Filloux, A.; De Bentzmann, S. Organization and PprB-Dependent Control of the Pseudomonas aeruginosa tad Locus, Involved in Flp Pilus Biology. J. Bacteriol. 2009, 191, 1961–1973. [Google Scholar] [CrossRef] [Green Version]
- Burrows, L.L. Pseudomonas aeruginosa Twitching Motility: Type IV Pili in Action. Annu. Rev. Microbiol. 2012, 66, 493–520. [Google Scholar] [CrossRef] [Green Version]
- Schilling, J.; Wagner, K.; Seekircher, S.; Greune, L.; Humberg, V.; Schmidt, M.A.; Heusipp, G. Transcriptional Activation of the tad Type IVb Pilus Operon by PypB in Yersinia enterocolitica. J. Bacteriol. 2010, 192, 3809–3821. [Google Scholar] [CrossRef] [Green Version]
- Walukiewicz, H.E.; Tohidifar, P.; Ordal, G.W.; Rao, C.V. Interactions among the three adaptation systems of Bacillus subtilis chemotaxis as revealed by an in vitro receptor-kinase assay. Mol. Microbiol. 2014, 93, 1104–1118. [Google Scholar] [CrossRef]
- Huang, Z.; Pan, X.; Xu, N.; Guo, M. Bacterial chemotaxis coupling protein: Structure, function and diversity. Microbiol. Res. 2019, 219, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Ishihama, A. Transcriptional response of Escherichia coli to external copper. Mol. Microbiol. 2005, 56, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Gudipaty, S.A.; Larsen, A.S.; Rensing, C.; McEvoy, M.M. Regulation of Cu(I)/Ag(I) efflux genes in Escherichia coli by the sensor kinase CusS. FEMS Microbiol. Lett. 2012, 330, 30–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, M.; Zhu, X.; Fan, F.; Xu, H.; Tang, J.; Qin, Y.; Ma, Y.; Zhang, X. Osmotolerance in Escherichia coli Is Improved by Activation of Copper Efflux Genes or Supplementation with Sulfur-Containing Amino Acids. Appl. Environ. Microbiol. 2017, 83, e03050-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nunes, I.; Jacquiod, S.; Brejnrod, A.; Holm, P.E.; Johansen, A.; Brandt, K.K.; Priemé, A.; Sørensen, S.J. Coping with copper: Legacy effect of copper on potential activity of soil bacteria following a century of exposure. FEMS Microbiol. Ecol. 2016, 92, fiw175. [Google Scholar] [CrossRef] [Green Version]
- Jain, D.; Kour, R.; Bhojiya, A.A.; Meena, R.H.; Singh, A.; Mohanty, S.R.; Rajpurohit, D.; Ameta, K.D. Zinc tolerant plant growth promoting bacteria alleviates phytotoxic effects of zinc on maize through zinc immobilization. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, M.; Goto, M.; Hibi, T. Similarity between Copper Resistance Genes from Pseudomonas syringae pv. actinidiae and P. syringae pv. tomato. J. Gen. Plant Pathol. 2002, 68, 68–74. [Google Scholar] [CrossRef]
- Colonnella, M.A.; Lizarraga, L.; Rossi, L.; Peña, R.D.; Egoburo, D.; López, N.I.; Iustman, L.J.R. Effect of copper on diesel degradation in Pseudomonas extremaustralis. Extremophiles 2019, 23, 91–99. [Google Scholar] [CrossRef]
- Kang, S.-M.; Asaf, S.; Khan, A.L.; Lubna; Khan, A.; Mun, B.-G.; Khan, M.A.; Gul, H.; Lee, I.-J. Complete Genome Sequence of Pseudomonas psychrotolerans CS51, a Plant Growth-Promoting Bacterium, Under Heavy Metal Stress Conditions. Microorganisms 2020, 8, 382. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.-M.; Shahzad, R.; Bilal, S.; Khan, A.L.; You, Y.-H.; Lee, W.-H.; Ryu, H.-L.; Lee, K.-E.; Lee, I.-J. Metabolism-mediated induction of zinc tolerance in Brassica rapa by Burkholderia cepacia CS2-1. J. Microbiol. 2017, 55, 955–965. [Google Scholar] [CrossRef] [PubMed]
- Bagdasarian, M.; Lurz, R.; Ruckert, B.; Franklin, F.; E Frey, J.; Timmis, K.N. Specific-purpose plasmid cloning vectors II. Broad host range, high copy number, RSF 1010-derived vectors, and a host-vector system for gene cloning in Pseudomonas. Gene 1981, 16, 237–247. [Google Scholar] [CrossRef]
- Rodríguez-Moreno, L.; Jiménez, A.J.; Ramos, C. Endopathogenic lifestyle of Pseudomonas savastanoi pv. savastanoi in olive knots. Microb. Biotechnol. 2009, 2, 476–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collado-Romero, M.; Mercado-Blanco, J.; Olivares-García, C.; Valverde-Corredor, A.; Jiménez-Díaz, R.M. Molecular Variability Within and Among Verticillium dahliae Vegetative Compatibility Groups Determined by Fluorescent Amplified Fragment Length Polymorphism and Polymerase Chain Reaction Markers. Phytopathology 2006, 96, 485–495. [Google Scholar] [CrossRef] [Green Version]
- Darling, A.E.; Mau, B.; Perna, N.T. progressiveMauve: Multiple Genome Alignment with Gene Gain, Loss and Rearrangement. PLoS ONE 2010, 5, e11147. [Google Scholar] [CrossRef] [Green Version]
- Huertas-Rosales, Ó.; Ramos-González, M.I.; Espinosa-Urgel, M. Self-Regulation and Interplay of Rsm Family Proteins Modulate the Lifestyle of Pseudomonas putida. Appl. Environ. Microbiol. 2016, 82, 5673–5686. [Google Scholar] [CrossRef] [Green Version]
- Hosseinkhani, B.; Emtiazi, G.; Nahvi, I. Analysis of phytase producing bacteria (Pseudomonas sp.) from poultry faeces and optimization of this enzyme production. Afr. J. Biotechnol. 2009, 8. [Google Scholar] [CrossRef]
- Rademaker, J.L.W.; de Bruijn, F.J. Characterization and classification of microbes by rep-PCR genomic fingerprinting and computer assisted pattern analysis. In DNA Markers: Protocols, Applications and Overviews; Caetano-Anollés, G., Gresshoff, P.M., Eds.; J. Wiley & Sons: New York, NY, USA, 1997; Volume 10, pp. 151–171. [Google Scholar]
- Versalovic, J.; Schneider, M.; Bruijn, F.J.D.; Lupski, J.R. Genomic fingerprinting of bacteria using repetitive sequence-based polymerase chain reaction. Methods Mol. Biol. 1994, 5, 25–40. [Google Scholar]
- Miller, E.M.; Nickoloff, J.A. Escherichia coli Electrotransformation. In Electroporation Protocols for Microorganisms; Humana Press: Totowa, NJ, USA, 2003; Volume 47, pp. 105–114. [Google Scholar]
- Leyva-Pérez, M.d.l.O.; Jiménez-Ruiz, J.; Gómez-Lama Cabanás, C.; Valverde-Corredor, A.; Barroso, J.B.; Luque, F.; Mercado-Blanco, J. Tolerance of olive (Olea europaea) cv Frantoio to Verticillium dahliae relies on both basal and pathogen-induced differential transcriptomic responses. New Phytol. 2017, 217, 671–686. [Google Scholar] [CrossRef] [Green Version]
- Campbell, C.L.; Madden, L.V. Introduction to Plant Disease Epidemiology; John Wiley & Sons: New York, NY, USA, 1990. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mutant Name | Length of the DNA Sequence Analyzed | Phenotype | Accession Number | Closest Strain | Query Cover (%) | E-Value | Identity (%) | Definition (GenBank) | Definition (GenName) |
|---|---|---|---|---|---|---|---|---|---|
| Cop1 | 294 | Modified copper tolerance | WP_069672392.1 | Pseudomonas simiae WCS417 | 61 | 1 × 10−31 | 100 | chemotaxis protein CheV | cheV |
| Cop33 | 549 | WP_176332083.1 | Pseudomonas simiae WCS417 | 41 | 8 × 10−44 | 100 | Heavy metal response regulator transcription factor | cusR | |
| Bfm8 | 468 | Impaired in biofilm formation | PHX42735.1 | Pseudomonas simiae PCL1751 | 100 | 7 × 10−108 | 100 | ATPase AAA | fleQ |
| Bfm9 | 212 | WP_045791339.1 | Pseudomonas simiae PCL1751 | 99 | 4 × 10−21 | 100 | membrane protein | - 1 |
| Experiment I | Experiment II | ||||
|---|---|---|---|---|---|
| Media | Media | ||||
| Strain | PDA | NA | Strain | PDA | NA |
| PICF7 | 41.49 | 21.70 | PICF7 | 47.69 | 21.90 |
| Cop1 | 47.68 | 27.63 | Cop1 | 42.38 | 26.97 |
| Cop33 | 41.72 | 26.97 | Cop33 | 42.38 | 23.68 |
| Bfm8 | 64.36 * | 29.72 * | Bfm8 | 46.15 | 21.90 |
| Bfm9 | 37.77 | 20.75 | Bfm9 | 54.36 * | 25.24 |
| Phy17 | ni | 25.47 | Phy17 | 0.51 * | 30.48 |
| Phy18 | ni | 21.23 | Phy18 | 1.03 * | 18.10 |
| Experiment I | Experiment II | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Disease Parameters | Disease Parameters | |||||||||
| Treatments | FS | Final DI (%) | Final DII | M (%) | AUDPC | FS | Final DI (%) | Final DII | M (%) | AUDPC |
| V. dahliae | 2.13a | 91.67 | 0.53 | 25 | 88.11a | 2.79a | 91.67 | 0.70 | 41.67 | 101.64a |
| V. dahliae/PICF7 | 1.17b | 75 | 0.29 | 16.67 | 31.43b | 2.04ab | 66.67 | 0.51 | 33.33 | 83.75ab |
| V. dahliae/Bfm8 | 0.63b | 91.67 | 0.15 | 0 | 19.71b | 1.58bc | 66.67 | 0.40 | 16.67 | 59.00bc |
| V. dahliae/Bfm9 | 1.00b | 75 | 0.25 | 0 | 29.24b | 1.50bc | 50 | 0.38 | 33.33 | 56.10bc |
| V. dahliae/Phy17 | 0.81b | 83.33 | 0.2 | 0 | 19.02b | 0.96c | 50 | 0.24 | 8.33 | 38.19c |
| V. dahliae/Phy18 | 0.85b | 75 | 0.21 | 8.33 | 24.35b | 1.04c | 33.33 | 0.26 | 16.67 | 30.06c |
| V. dahliae/Cop1 | 1.27b | 83.33 | 0.31 | 8.33 | 40.09b | 1.56bc | 66.67 | 0.39 | 25 | 49.58bc |
| V. dahliae/Cop33 | 0.92b | 91.67 | 0.22 | 0 | 31.51b | 1.31bc | 66.67 | 0.33 | 16.67 | 43.44bc |
| Microorganisms and Plasmid | Characteristics | Reference/Source |
|---|---|---|
| Bacterial Strains and Mutants | ||
| Escherichia coli DH5α | recA1 endA1 Φ80d lacZ dam-15 | Clontech |
| Bacillus sp. PIC28 | PGPR, negative control used in phytase activity assays | [7] |
| Pseudomonas indica PIC128 | PGPR, negative control used in biofilm formation assays, isolated from the rhizosphere of a nursery-produced olive plant | Lab. Plant-Microorganism Interactions/This study |
| Pseudomonas putida KT2440 | PGPR, positive control used in biofilm formation assays | [84] |
| Pseudomonas simiae PICF7 | Wild-type PGPR | [14] |
| Cop1 | PICF7 Tn5-TcR mutant (cheV::Tn5-TcR) affected in copper resistance | This work |
| Cop33 | PICF7 Tn5-TcR mutant (CusR::Tn5-TcR) affected in copper resistance | This work |
| Bfm8 | PICF7 Tn5-TcR mutant (fleQ::Tn5-TcR) impaired in biofilm formation | This work |
| Bfm9 | PICF7 Tn5-TcR mutant impaired in biofilm formation | This work |
| Phy17 | PICF7 Tn5-TcR mutant deficient in phytase activity | This work |
| Phy18 | PICF7 Tn5-TcR mutant deficient in phytase activity | This work |
| P. simiae PICF7 (pLRM1) | PICF7 (GmR) GFP-labeled mutant derivative | [19] |
| Cop1 (pLRM1) | Cop1 (TcR and GmR) GFP-labeled mutant derivative | This work |
| Cop33 (pLRM1) | Cop33 (TcR and GmR) GFP-labeled mutant derivative | This work |
| Bfm8 (pLRM1) | Bfm8 (TcR and GmR) GFP-labeled mutant derivative | This work |
| Bfm9 (pLRM1) | Bfm9 (TcR and GmR) GFP-labeled mutant derivative | This work |
| Phy17 (pLRM1) | Phy17 (TcR and GmR) GFP-labeled mutant derivative | This work |
| Phy18 (pLRM1) | Phy18 (TcR and GmR) GFP-labeled mutant derivative | This work |
| Plasmid | ||
| pLRM1 | pBBR1-MCS5 carrying a fusion of the PA1/04/03 promoter to the gfpmut3* gene, GmR | [85] |
| Fungal pathogen | ||
| Verticillium dahliae V937I | Representative of the defoliating pathotype, originating from a diseased olive tree | [86] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montes-Osuna, N.; Gómez-Lama Cabanás, C.; Valverde-Corredor, A.; Berendsen, R.L.; Prieto, P.; Mercado-Blanco, J. Assessing the Involvement of Selected Phenotypes of Pseudomonas simiae PICF7 in Olive Root Colonization and Biological Control of Verticillium dahliae. Plants 2021, 10, 412. https://doi.org/10.3390/plants10020412
Montes-Osuna N, Gómez-Lama Cabanás C, Valverde-Corredor A, Berendsen RL, Prieto P, Mercado-Blanco J. Assessing the Involvement of Selected Phenotypes of Pseudomonas simiae PICF7 in Olive Root Colonization and Biological Control of Verticillium dahliae. Plants. 2021; 10(2):412. https://doi.org/10.3390/plants10020412
Chicago/Turabian StyleMontes-Osuna, Nuria, Carmen Gómez-Lama Cabanás, Antonio Valverde-Corredor, Roeland L. Berendsen, Pilar Prieto, and Jesús Mercado-Blanco. 2021. "Assessing the Involvement of Selected Phenotypes of Pseudomonas simiae PICF7 in Olive Root Colonization and Biological Control of Verticillium dahliae" Plants 10, no. 2: 412. https://doi.org/10.3390/plants10020412
APA StyleMontes-Osuna, N., Gómez-Lama Cabanás, C., Valverde-Corredor, A., Berendsen, R. L., Prieto, P., & Mercado-Blanco, J. (2021). Assessing the Involvement of Selected Phenotypes of Pseudomonas simiae PICF7 in Olive Root Colonization and Biological Control of Verticillium dahliae. Plants, 10(2), 412. https://doi.org/10.3390/plants10020412