
(-)-Leucophyllone, a Tirucallane Triterpenoid from Cornus walteri, Enhances Insulin Secretion in INS-1 Cells





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
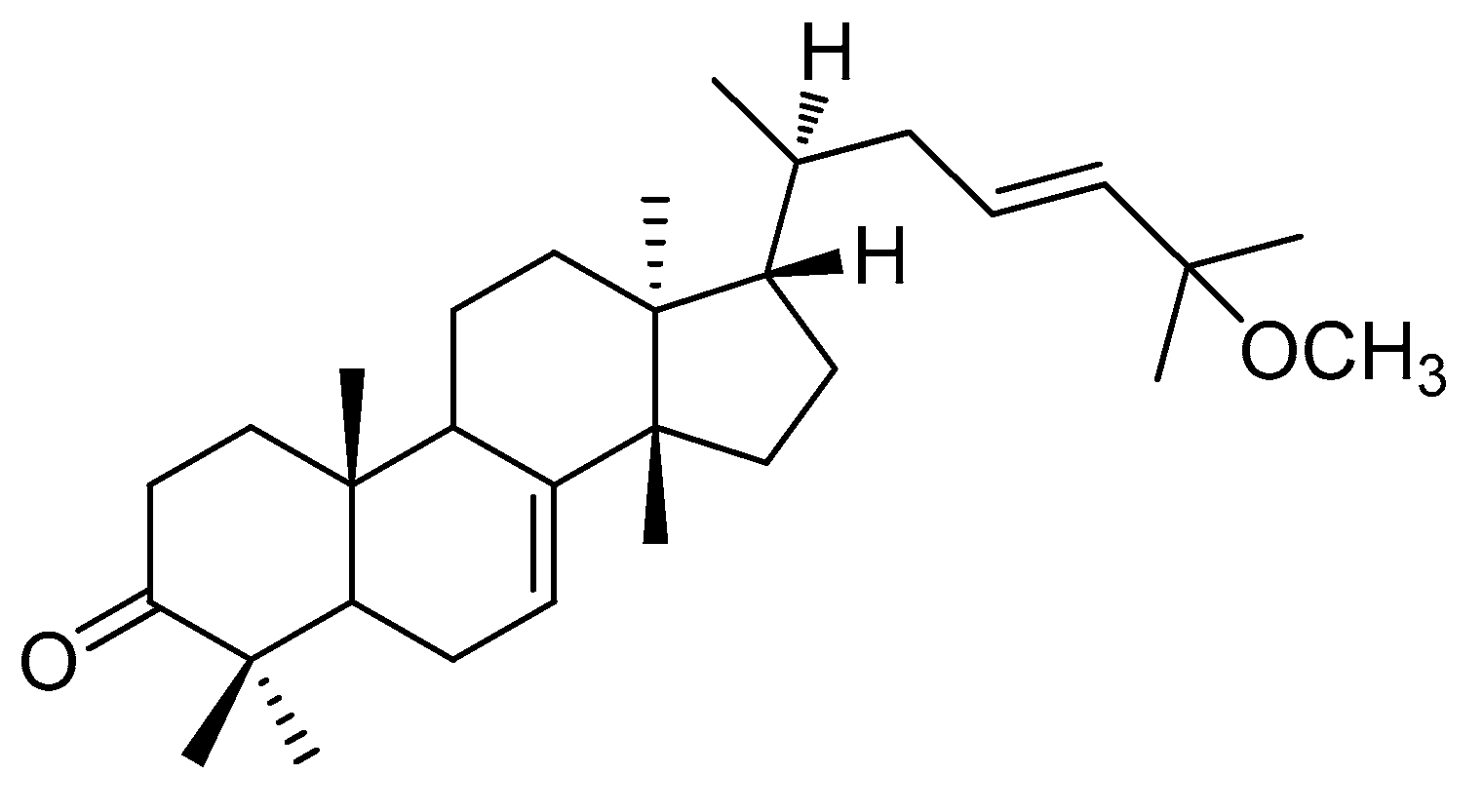
2.1. Isolation and Identification of (-)-Leucophyllone from C. walteri
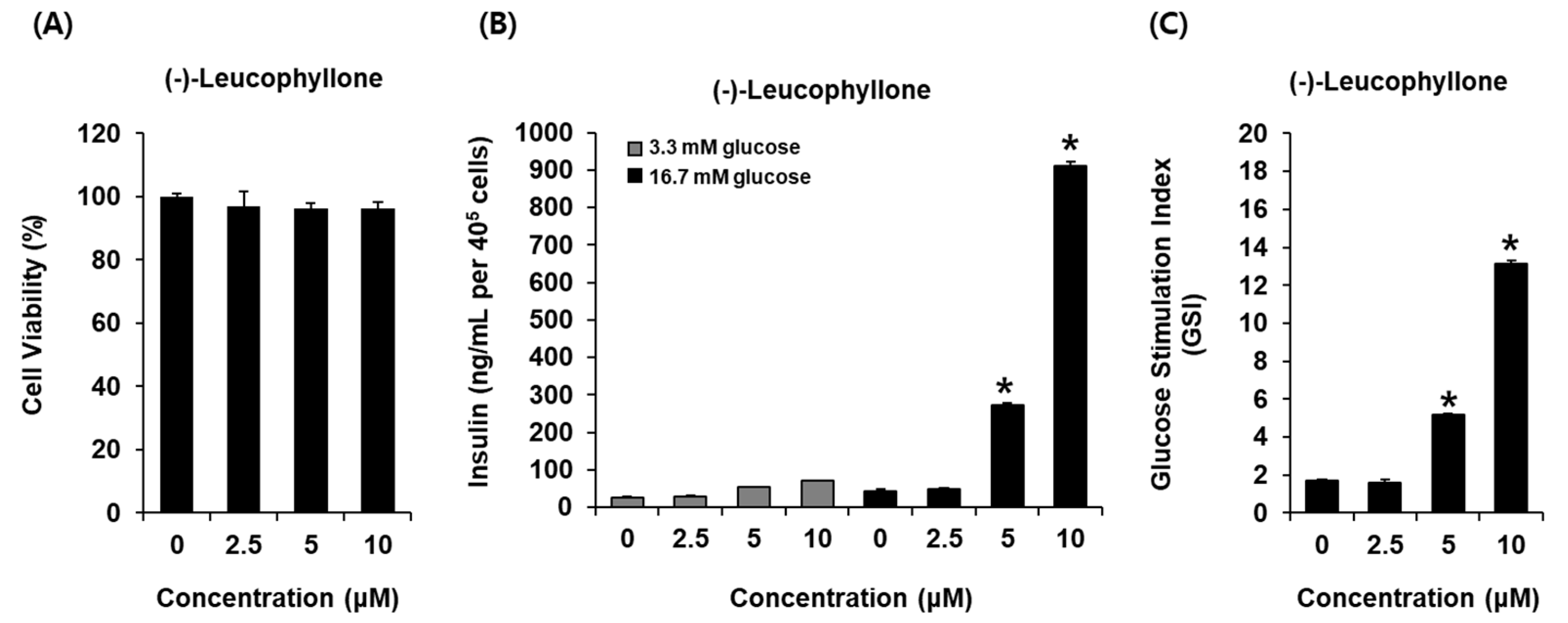
2.2. Glucose-Stimulated Insulin Secretion (GSIS) Effect
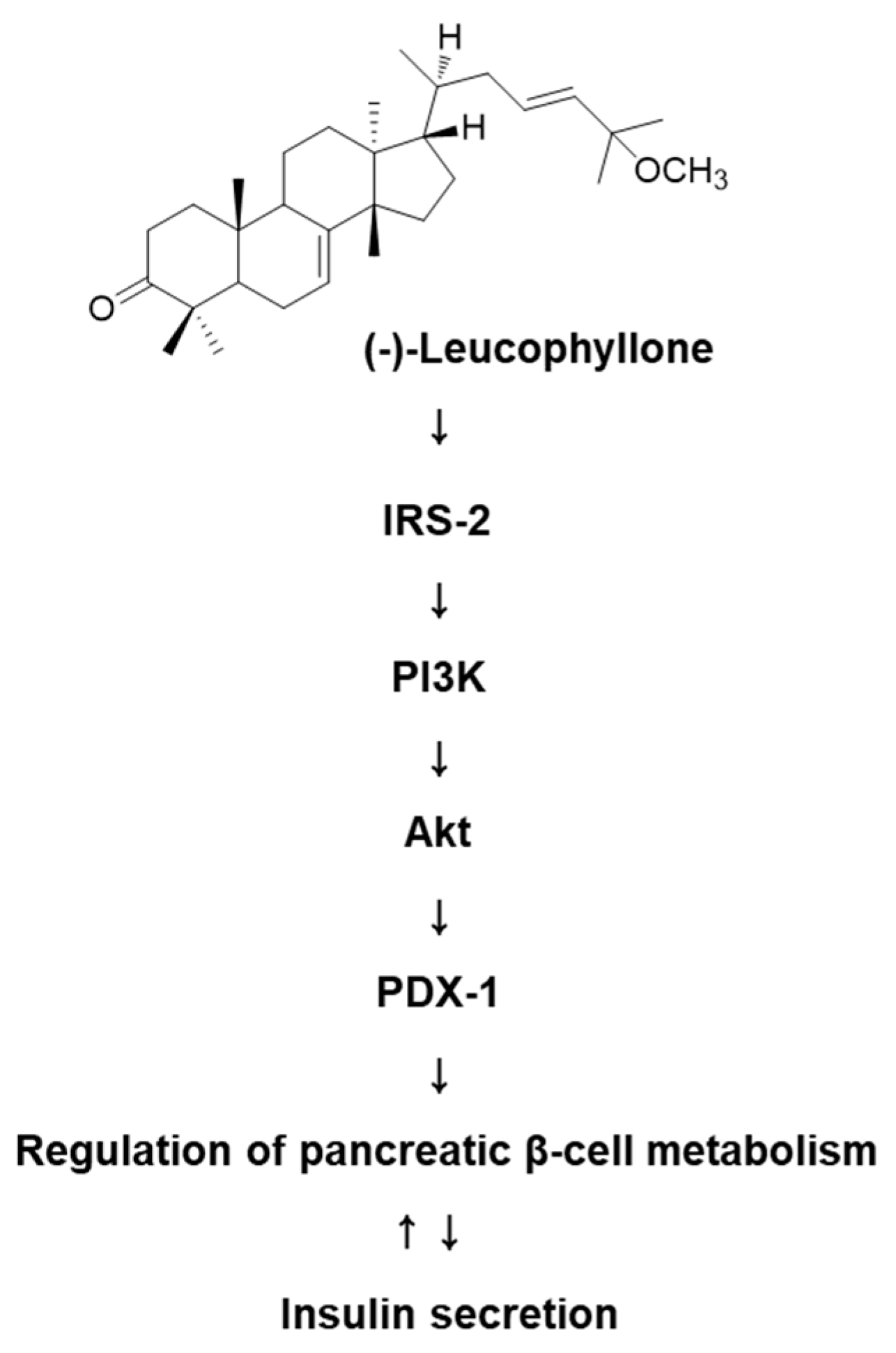
2.3. Protein Expression of IRS-2 (Ser731), P-IRS-2, PI3K, P-PI3K, Akt, P-Akt (Ser473), and PDX-1
3. Discussion
4. Materials and Methods
4.1. Extraction, Fractionation, and Purification Methods
4.2. Cell Culture
4.3. Measurement of Cell Viability
4.4. GSIS Assay
4.5. Western Blotting
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hameed, I.; Masoodi, S.R.; Mir, S.A.; Nabi, M.; Ghazanfar, K.; Ganai, B.A. Type 2 diabetes mellitus: From a metabolic disorder to an inflammatory condition. World J. Diabetes 2015, 6, 598. [Google Scholar] [CrossRef]
- Seino, S.; Shibasaki, T.; Minami, K. Dynamics of insulin secretion and the clinical implications for obesity and diabetes. J. Clin. Investig. 2011, 121, 2118–2125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, A.; Jones, L.C.; de Koning, E.; Hansen, B.C.; Matthews, D.R. Decreased insulin secretion in type 2 diabetes: A problem of cellular mass or function? Diabetes 2001, 50, S169. [Google Scholar] [CrossRef] [Green Version]
- Katiyar, C.; Gupta, A.; Kanjilal, S.; Katiyar, S. Drug discovery from plant sources: An integrated approach. Ayu 2012, 33, 10. [Google Scholar] [CrossRef]
- Chang, C.L.-T.; Liu, H.-Y.; Kuo, T.-F.; Hsu, Y.-J.; Shen, M.-Y.; Pan, C.-Y.; Yang, W.-C. Antidiabetic effect and mode of action of cytopiloyne. Evid. Based Complement. Altern. Med. 2013, 2013, 685642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akiba, Y.; Kato, S.; Katsube, K.-I.; Nakamura, M.; Takeuchi, K.; Ishii, H.; Hibi, T. Transient receptor potential vanilloid subfamily 1 expressed in pancreatic islet β cells modulates insulin secretion in rats. Biochem. Biophys. Res. Commun. 2004, 321, 219–225. [Google Scholar] [CrossRef]
- Leng, S.-H.; Lu, F.-E.; Xu, L.-J. Therapeutic effects of berberine in impaired glucose tolerance rats and its influence on insulin secretion. Acta Pharmacol. Sin. 2004, 25, 496–502. [Google Scholar]
- Best, L.; Elliott, A.C.; Brown, P.D. Curcumin induces electrical activity in rat pancreatic β-cells by activating the volume-regulated anion channel. Biochem. Pharmacol. 2007, 73, 1768–1775. [Google Scholar] [CrossRef]
- Cai, E.P.; Lin, J.-K. Epigallocatechin gallate (EGCG) and rutin suppress the glucotoxicity through activating IRS2 and AMPK signaling in rat pancreatic β cells. J. Agric. Food Chem. 2009, 57, 9817–9827. [Google Scholar] [CrossRef] [PubMed]
- Ohno, T.; Kato, N.; Ishii, C.; Shimizu, M.; Ito, Y.; Tomono, S.; Kawazu, S. Genistein augments cyclic adenosine 3’5’-monophosphate(cAMP) accumulation and insulin release in MIN6 cells. Endocr. Res. 1993, 19, 273–285. [Google Scholar] [CrossRef]
- Choi, W.-H.; Park, W.-Y.; Hwang, B.-Y.; Oh, G.-J.; Kang, S.-J.; Lee, K.-S.; Ro, J.-S. Phenolic compounds from the stem bark of Cornus walteri Wanger. Korean J. Pharmacogn. 1998, 29, 217–224. [Google Scholar]
- Park, H.-C.; Jung, T.K.; Kim, M.J.; Yoon, K.-S. Protective effect of Cornus walteri Wangerin leaf against UVB irradiation induced photoaging in human reconstituted skin. J. Ethnopharmacol. 2016, 193, 445–449. [Google Scholar] [CrossRef]
- Lim, C.-S.; Li, C.-Y.; Kim, Y.-M.; Lee, W.-Y.; Rhee, H.-I. The inhibitory effect of Cornus walteri extract against α−amylase. Appl. Biol. Chem. 2005, 48, 103–108. [Google Scholar]
- Lee, S.-H.; Yoon, K.-R.; Lee, E.; Cha, Y.-Y. Anti-inflammatory effect of Cornus walteri. J. Physiol. Pathol. Korean Med. 2011, 25, 982–988. [Google Scholar]
- Park, W.-H.; Cha, Y.-Y. Effects of stem bark extracts of Cornus walteri Wanger on the lipid lowering, anti-oxidative activity and concentration of proinflammatory cytokines in rat fed high fat diet. J. Korean Med. Rehabil. 2009, 19, 59–78. [Google Scholar]
- Lee, D.; Lee, S.R.; Kang, K.S.; Ko, Y.; Pang, C.; Yamabe, N.; Kim, K.H. Betulinic acid suppresses ovarian cancer cell proliferation through induction of apoptosis. Biomolecules 2019, 9, 257. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.; Lee, S.R.; Kang, K.S.; Kim, K.H. Benzyl salicylate from the stems and stem barks of Cornus walteri as a nephroprotective agent against cisplatin-induced apoptotic cell death in LLC-PK1 cells. RSC Adv. 2020, 10, 5777–5784. [Google Scholar] [CrossRef]
- Benosman, A.; Richomme, P.; Sevenet, T.; Perromat, G.; Hadi, A.H.A.; Bruneton, J. Tirucallane triterpenes from the stem bark of Aglaia leucophylla. Phytochemistry 1995, 40, 1485–1487. [Google Scholar] [CrossRef]
- Komatsu, M.; Takei, M.; Ishii, H.; Sato, Y. Glucose-stimulated insulin secretion: A newer perspective. J. Diabetes Investig. 2013, 4, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Kahn, B.B. Type 2 diabetes: When insulin secretion fails to compensate for insulin resistance. Cell 1998, 92, 593–596. [Google Scholar] [CrossRef] [Green Version]
- Soares, J.M.D.; Pereira Leal, A.E.B.; Silva, J.C.; Almeida, J.; de Oliveira, H.P. Influence of flavonoids on mechanism of modulation of insulin secretion. Pharmacogn. Mag. 2017, 13, 639–646. [Google Scholar]
- White, M.F. IRS proteins and the common path to diabetes. Am. J. Physiol. Endocrinol. Metab. 2002, 283, E413–E422. [Google Scholar] [CrossRef] [Green Version]
- Costa, M.M.; Violato, N.M.; Taboga, S.R.; Góes, R.M.; Bosqueiro, J.R. Reduction of insulin signalling pathway IRS-1/IRS-2/AKT/mTOR and decrease of epithelial cell proliferation in the prostate of glucocorticoid-treated rats. Int. J. Exp. Pathol. 2012, 93, 188–195. [Google Scholar] [CrossRef]
- Li, J.-M.; Wang, W.; Fan, C.-Y.; Wang, M.-X.; Zhang, X.; Hu, Q.-H.; Kong, L.-D. Quercetin preserves β-cell mass and function in fructose-induced hyperinsulinemia through modulating pancreatic Akt/FoxO1 activation. Evid. Based Complement. Altern. Med. 2013, 2013, 303902. [Google Scholar]
- Zhang, B.; Sun, P.; Liu, X.; Sun, J.; Li, D.; Liu, Z.; Zhang, W.; Zhang, K.; Niu, Y. Role and mechanism of PI3K/AKT/FoxO1/PDX-1 signaling pathway in functional changes of pancreatic islets in rats after severe burns. Life Sci. 2020, 258, 118145. [Google Scholar] [CrossRef]
- Gauthier, B.R.; Wiederkehr, A.; Baquié, M.; Dai, C.; Powers, A.C.; Kerr-Conte, J.; Pattou, F.; MacDonald, R.J.; Ferrer, J.; Wollheim, C.B. PDX1 deficiency causes mitochondrial dysfunction and defective insulin secretion through TFAM suppression. Cell Metab. 2009, 10, 110–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Liu, Q.; Zhou, Z.; Ikeda, Y. PDX1, Neurogenin-3, and MAFA: Critical transcription regulators for beta cell development and regeneration. Stem Cell Res. Ther. 2017, 8, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brissova, M.; Shiota, M.; Nicholson, W.E.; Gannon, M.; Knobel, S.M.; Piston, D.W.; Wright, C.V.; Powers, A.C. Reduction in pancreatic transcription factor PDX-1 impairs glucose-stimulated insulin secretion. J. Biol. Chem. 2002, 277, 11225–11232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Sterr, M.; Burtscher, I.; Böttcher, A.; Beckenbauer, J.; Siehler, J.; Meitinger, T.; Häring, H.-U.; Staiger, H.; Cernilogar, F.M. Point mutations in the PDX1 transactivation domain impair human β-cell development and function. Mol. Metab. 2019, 24, 80–97. [Google Scholar] [CrossRef]
- Iftikhar, A.; Aslam, B.; Iftikhar, M.; Majeed, W.; Batool, M.; Zahoor, B.; Amna, N.; Gohar, H.; Latif, I. Effect of Caesalpinia bonduc polyphenol extract on alloxan-induced diabetic rats in attenuating hyperglycemia by upregulating insulin secretion and inhibiting JNK signaling pathway. Oxid. Med. Cell. Longev. 2020, 2020, 9020219. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-W.; Yang, S.-H.; Kim, J.-B. Protein fractions from Korean mistletoe (Viscum album coloratum) extract induce insulin secretion from pancreatic beta cells. Evid. Based Complement. Altern. Med. 2014, 2014, 703624. [Google Scholar] [CrossRef]
- Zhang, Y.; Ren, C.; Lu, G.; Mu, Z.; Cui, W.; Gao, H.; Wang, Y. Anti-diabetic effect of mulberry leaf polysaccharide by inhibiting pancreatic islet cell apoptosis and ameliorating insulin secretory capacity in diabetic rats. Int. Immunopharmacol. 2014, 22, 248–257. [Google Scholar] [CrossRef]
- Kim, H.; Choi, P.; Kim, T.; Kim, Y.; Song, B.G.; Park, Y.-T.; Choi, S.-J.; Yoon, C.H.; Lim, W.-C.; Ko, H. Ginsenosides Rk1 and Rg5 inhibit transforming growth factor-β1-induced epithelial-mesenchymal transition and suppress migration, invasion, anoikis resistance, and development of stem-like features in lung cancer. J. Ginseng Res. 2021, 45, 134–148. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.-H.; Hwang, H.-J.; Kim, J.W.; Kim, J.A.; Lee, Y.B.; Roh, E.; Choi, K.M.; Baik, S.H.; Yoo, H.J. Ginsenoside compound-Mc1 attenuates oxidative stress and apoptosis in cardiomyocytes through an AMP-activated protein kinase–dependent mechanism. J. Ginseng Res. 2020, 44, 664–671. [Google Scholar] [CrossRef] [PubMed]
- Hoa, H.T.; Thu, N.T.; Dong, N.T.; Oanh, T.T.; Hien, T.T.; Ha, D.T. Effects of compounds from Physalis angulata on fatty acid synthesis and glucose metabolism in HepG2 cells via the AMP-activated protein kinase pathway. Nat. Prod. Sci. 2020, 26, 200–206. [Google Scholar]
- Ryu, Y.-S.; Hyun, J.-W.; Chung, H.-S. Fucoidan induces apoptosis in A2058 cells through ROS-exposed activation of MAPKs signaling pathway. Nat. Prod. Sci. 2020, 26, 191–199. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, D.; Kim, K.H.; Jang, T.; Kang, K.S. (-)-Leucophyllone, a Tirucallane Triterpenoid from Cornus walteri, Enhances Insulin Secretion in INS-1 Cells. Plants 2021, 10, 431. https://doi.org/10.3390/plants10030431
Lee D, Kim KH, Jang T, Kang KS. (-)-Leucophyllone, a Tirucallane Triterpenoid from Cornus walteri, Enhances Insulin Secretion in INS-1 Cells. Plants. 2021; 10(3):431. https://doi.org/10.3390/plants10030431
Chicago/Turabian StyleLee, Dahae, Ki Hyun Kim, Taesu Jang, and Ki Sung Kang. 2021. "(-)-Leucophyllone, a Tirucallane Triterpenoid from Cornus walteri, Enhances Insulin Secretion in INS-1 Cells" Plants 10, no. 3: 431. https://doi.org/10.3390/plants10030431
APA StyleLee, D., Kim, K. H., Jang, T., & Kang, K. S. (2021). (-)-Leucophyllone, a Tirucallane Triterpenoid from Cornus walteri, Enhances Insulin Secretion in INS-1 Cells. Plants, 10(3), 431. https://doi.org/10.3390/plants10030431