
Genomic Characterization and Expression Analysis of the SnRK Family Genes in Dendrobium officinale Kimura et Migo (Orchidaceae)



Abstract
:1. Introduction
2. Results
2.1. Genome-Wide Identification of DoSnRK Members in Dendrobium officinale
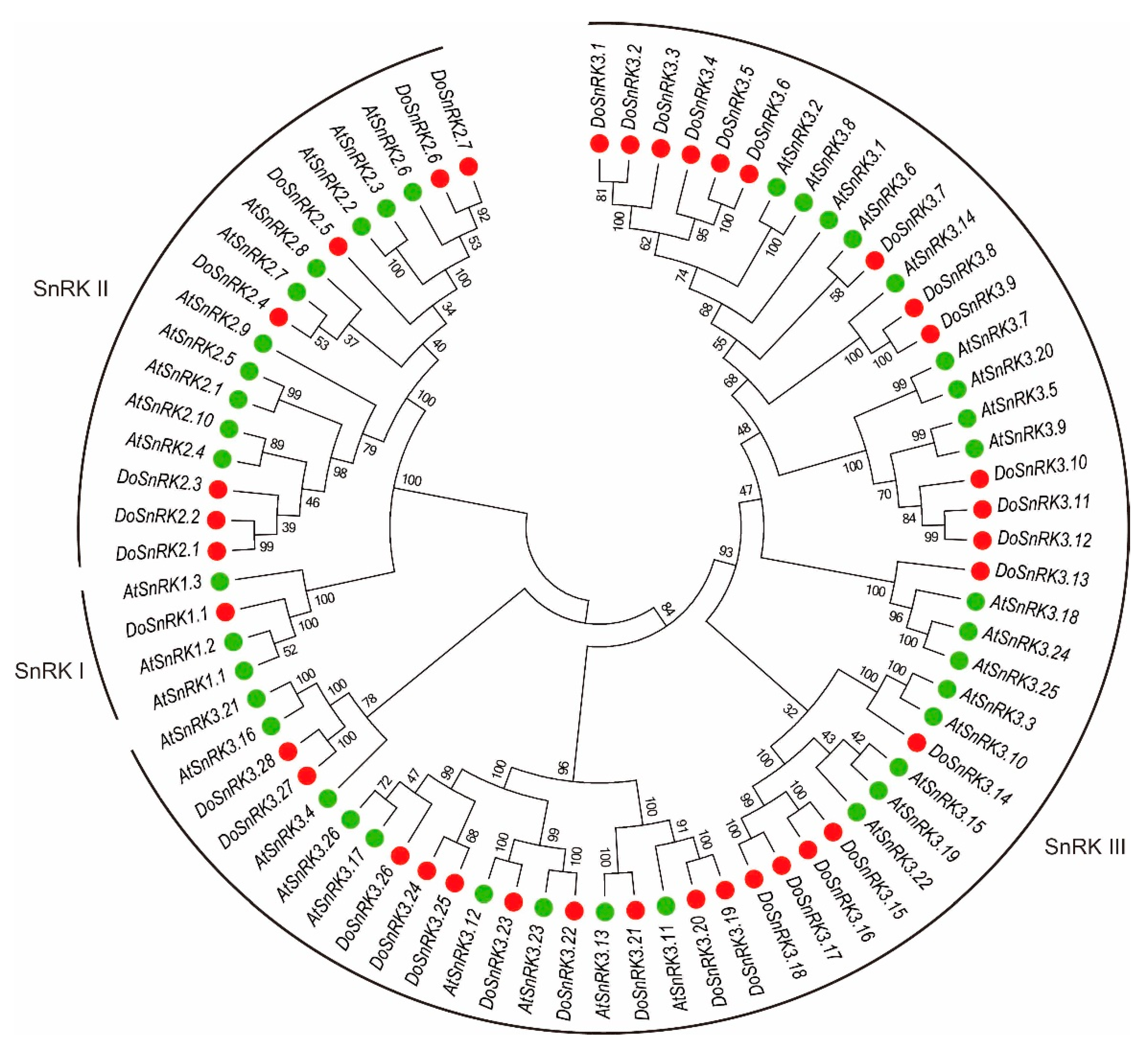
2.2. Phylogenetic Relationship of DoSnRK Proteins
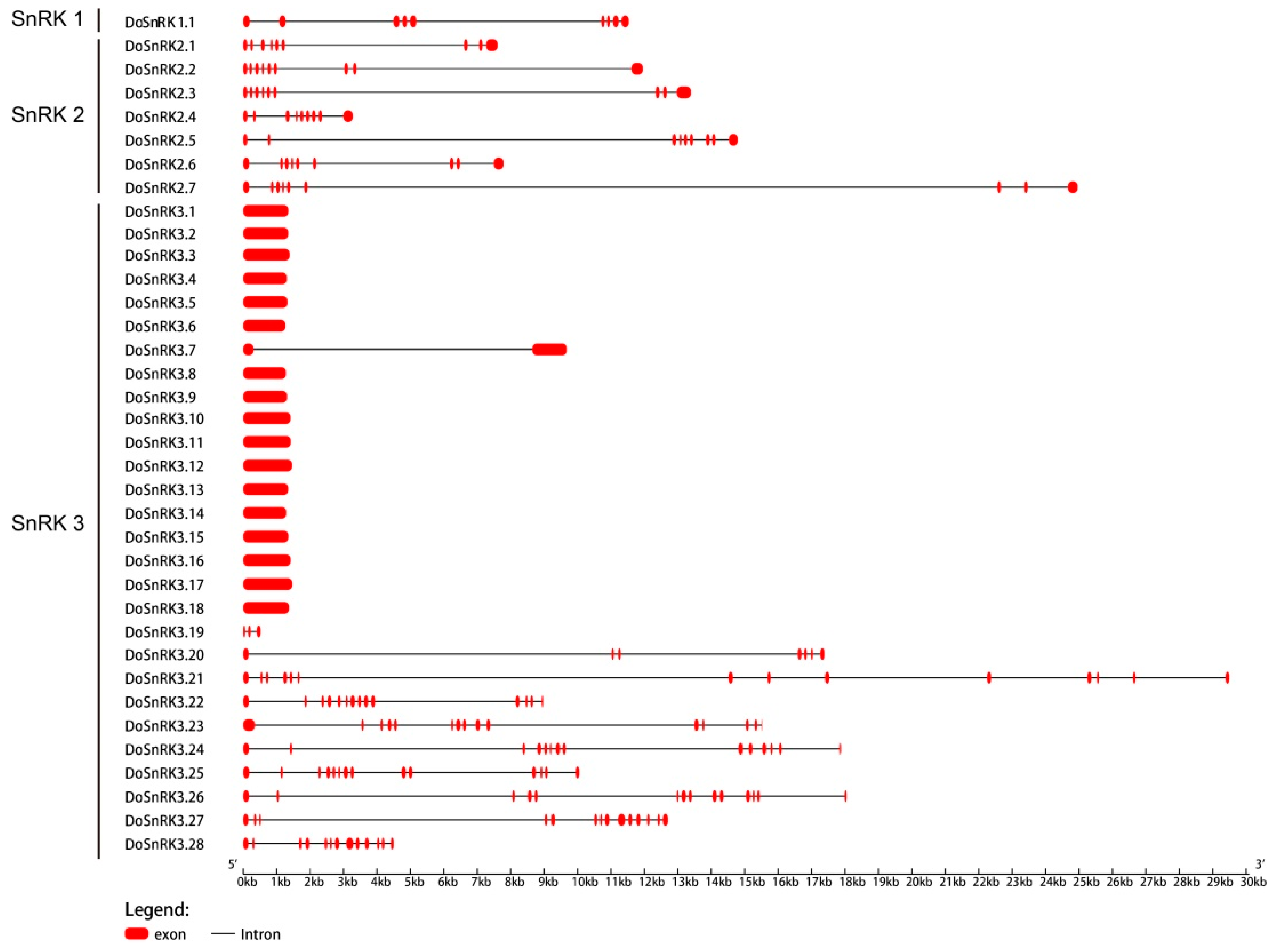
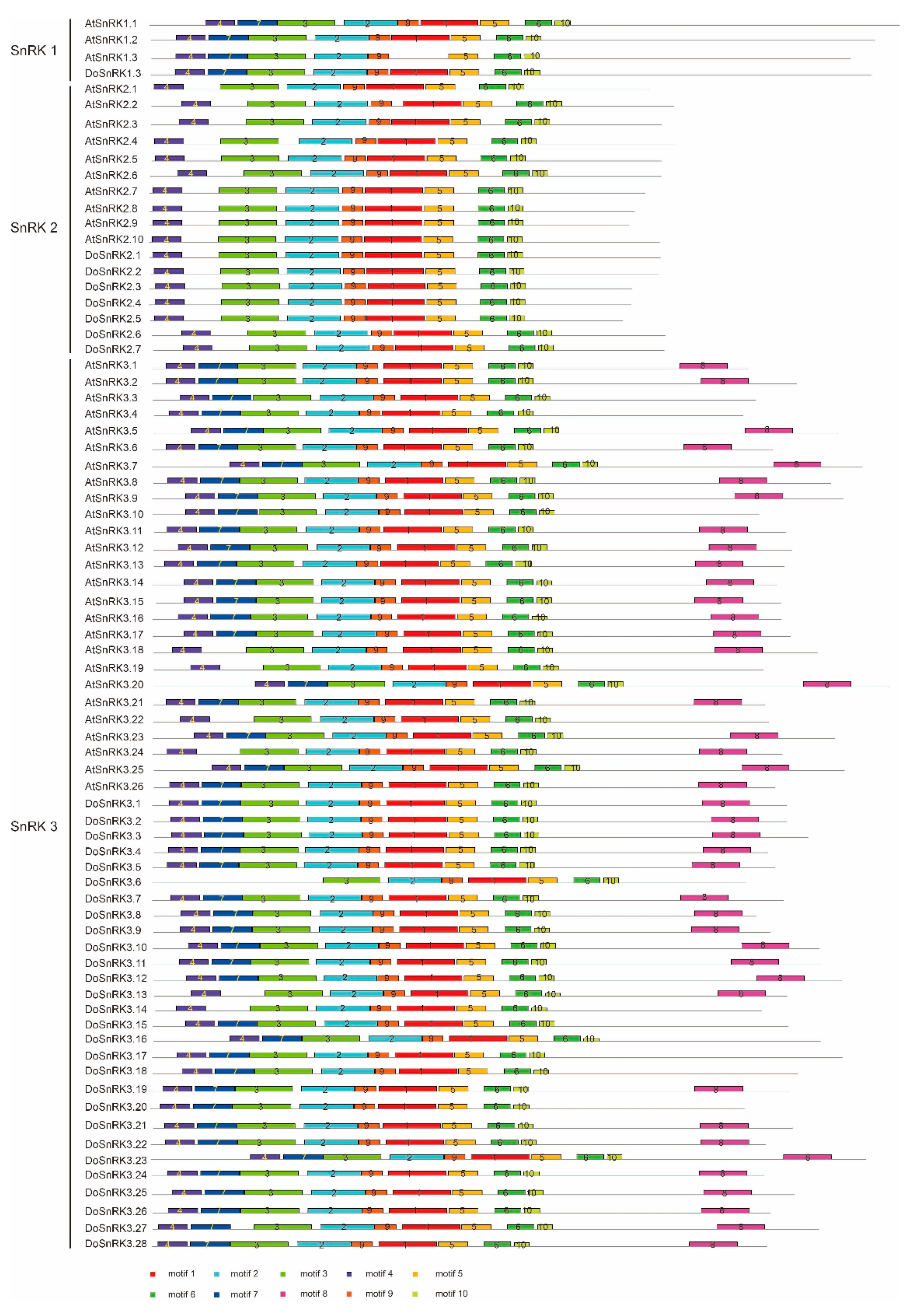
2.3. Gene Structures and Conserved Motifs of DoSnRK Genes
2.4. Cis-Elements in the Promoter Regions of DoSnRK Genes
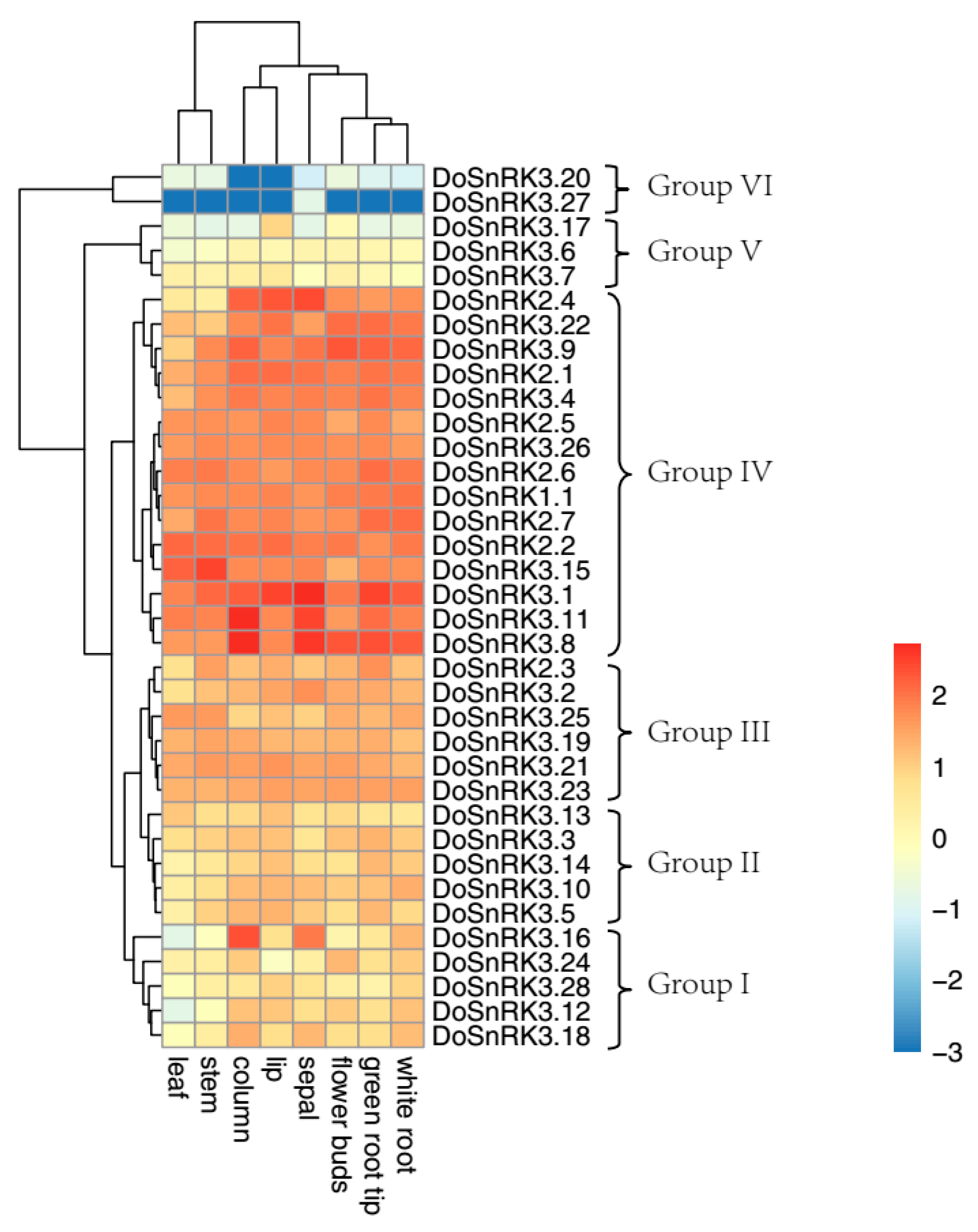
2.5. Expression Analysis of DoSnRK Genes in Eight Tissues
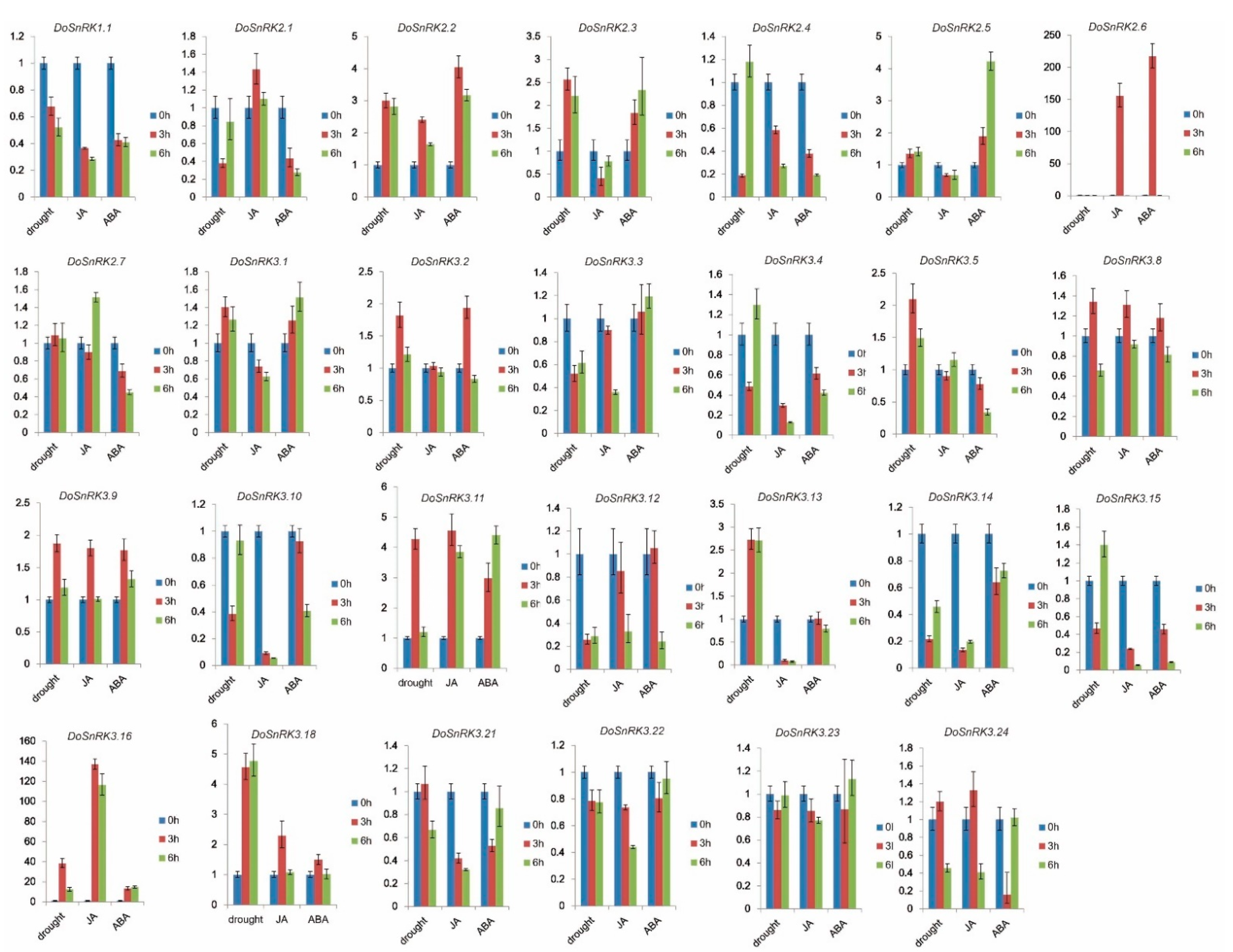
2.6. DoSnRK Gene Expression after Drought, MeJA, and ABA Treatments
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Plant Treatments
4.3. Identification of the SnRK Family in D. officinale
4.4. Phylogenetic Relationship of DoSnRK Proteins
4.5. Exon–Intron Structures and Conserved Motifs in DoSnRK Genes
4.6. Expression Profiles of DoSnRK Genes in Eight Tissues
4.7. qRT-PCR Assays
4.8. Cis-Element Analysis in Promoter Regions of DoSnRK Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Halford, N.G.; Hey, S.J. Snf1-related protein kinases (SnRKs) act within an intricate network that links metabolic and stress signalling in plants. Biochem. J. 2009, 419, 247–259. [Google Scholar] [CrossRef] [Green Version]
- Kulik, A.; Wawer, I.; Krzywińska, E.; Bucholc, M.; Dobrowolska, G. SnRK2 protein kinases--key regulators of plant response to abiotic stresses. OMICS 2011, 15, 859–872. [Google Scholar] [CrossRef]
- Crozet, P.; Margalha, L.; Confraria, A.; Rodrigues, A.; Martinho, C.; Adamo, M.; Elias, C.A.; Baena-González, E. Mechanisms of regulation of SNF1/AMPK/SnRK1 protein kinases. Front. Plant Sci. 2014, 5, 190. [Google Scholar] [CrossRef]
- Zheng, Z.; Xu, X.; Crosley, R.A.; Greenwalt, S.A.; Sun, Y.; Blakeslee, B.; Wang, L.; Ni, W.; Sopko, M.S.; Yao, C.; et al. The protein kinase SnRK2.6 mediates the regulation of sucrose metabolism and plant growth in Arabidopsis. Plant Physiol. 2010, 153, 99–113. [Google Scholar] [CrossRef] [Green Version]
- Coello, P.; Hey, S.J.; Halford, N.G. The sucrose non-fermenting-1-related (SnRK) family of protein kinases: Potential for ma-nipulation to improve stress tolerance and increase yield. J. Exp. Bot. 2011, 62, 883–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujii, H.; Verslues, P.E.; Zhu, J.K. Arabidopsis decuple mutant reveals the importance of SnRK2 kinases in osmotic stress re-sponses in vivo. Proc. Natl. Acad. Sci. USA 2011, 108, 1717–1722. [Google Scholar] [CrossRef] [Green Version]
- Emanuelle, S.; Doblin, M.S.; Stapleton, D.I.; Bacic, A.; Gooley, P.R. Molecular Insights into the Enigmatic Metabolic Regulator, SnRK1. Trends Plant Sci. 2016, 21, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Lou, D.; Wang, H.; Liang, G.; Yu, D. OsSAPK2 Confers Abscisic Acid Sensitivity and Tolerance to Drought Stress in Rice. Front. Plant Sci. 2017, 8, 993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, R.; Wang, Y.; Gai, R.; Xi, D.; Mao, C.; Ming, F. Rice SnRK protein kinase OsSAPK8 acts as a positive regulator in abiotic stress responses. Plant Sci. 2020, 292, 110373. [Google Scholar] [CrossRef] [PubMed]
- Hrabak, E.M.; Chan, C.W.; Gribskov, M.; Harper, J.F.; Choi, J.H.; Halford, N.; Kudla, J.; Luan, S.; Nimmo, H.G.; Sussman, M.R.; et al. The Arabidopsis CDPK-SnRK superfamily of protein kinases. Plant Physiol. 2003, 132, 666–680. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, Y.; Yamamoto, S.; Minami, H.; Kagaya, Y.; Hattori, T. Differential activation of the rice sucrose nonferment-ing1-related protein kinase2 family by hyperosmotic stress and abscisic acid. Plant Cell. 2004, 16, 1163–1177. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.T.; Ye, Y.Y.; Jiang, L.Y.; Lin, Y.X.; Gu, X.J.; Chen, Q.; Sun, B.; Zhang, Y.; Luo, Y.; Wang, Y.; et al. Genome-Wide Characterization of Snf1-Related Protein Kinases (SnRKs) and Expression Analysis of SnRK1.1 in Strawberry. Gene 2020, 11, 427. [Google Scholar] [CrossRef]
- Wu, G.Q.; Liu, Z.X.; Xie, L.L.; Wang, J.L. Genome-Wide Identification and Expression Analysis of the BvSnRK2 Genes Family in Sugar Beet (Beta vulgaris L.) Under Salt Conditions. J. Plant Growth Regul. 2020, 1–14. [Google Scholar] [CrossRef]
- Wurzinger, B.; Nukarinen, E.; Nägele, T.; Weckwerth, W.; Teige, M. The SnRK1 Kinase as Central Mediator of Energy Signaling between Different Organelles. Plant Physiol. 2018, 176, 1085–1094. [Google Scholar] [CrossRef] [Green Version]
- Purcell, P.C.; Smith, A.M.; Halford, N.G. Antisense expression of a sucrose nonfermenting-1-related protein kinase sequence in potato results in decreased expression of sucrose synthase in tubers and loss of sucrose-inducibility of sucrose synthase tran-scripts in leaves. Plant J. 1998, 14, 195–202. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Yan, H.; Qiu, Z.; Hu, B.; Zeng, B.; Zhong, C.; Fan, C. Comprehensive Analysis of SnRK Gene Family and their Re-sponses to Salt Stress in Eucalyptus grandis. Int. J. Mol. Sci. 2019, 20, 2786. [Google Scholar] [CrossRef] [Green Version]
- Filipe, O.; De Vleesschauwer, D.; Haeck, A.; Demeestere, K.; Höfte, M. The energy sensor OsSnRK1a confers broad-spectrum disease resistance in rice. Sci. Rep. 2018, 8, 3864. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wang, H.; He, S.; Meng, F.; Zhang, C.; Fan, S.; Wu, J.; Zhang, S.; Xu, P. GmSnRK1.1, a Sucrose Non-fermenting-1(SNF1)-Related Protein Kinase, Promotes Soybean Resistance to Phytophthora sojae. Front. Plant Sci. 2019, 10, 996. [Google Scholar] [CrossRef] [Green Version]
- Chan, A.; Carianopol, C.; Tsai, A.Y.; Varatharajah, K.; Chiu, R.S.; Gazzarrini, S. SnRK1 phosphorylation of FUSCA3 positively regulates embryogenesis, seed yield, and plant growth at high temperature in Arabidopsis. J. Exp. Bot. 2017, 68, 4219–4231. [Google Scholar] [CrossRef] [Green Version]
- Shin, R.; Alvarez, S.; Burch, A.Y.; Jez, J.M.; Schachtman, D.P. Phosphoproteomic identification of targets of the Arabidopsis sucrose nonfermenting-like kinase SnRK2.8 reveals a connection to metabolic processes. Proc. Natl. Acad. Sci. USA 2007, 104, 6460–6465. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Wang, Y.P.; Chen, P.; Ren, J.; Ji, K.; Li, Q.; Li, P.; Dai, S.J.; Leng, P. Transcriptional regulation of SlPYL, SlPP2C, and SlSnRK2 gene families encoding ABA signal core components during tomato fruit development and drought stress. J. Exp. Bot. 2011, 62, 5659–5669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umezawa, T.; Yoshida, R.; Maruyama, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K. SRK2C, a SNF1-related protein kinase 2, improves drought tolerance by controlling stress-responsive gene expression in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2004, 101, 17306–17311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dey, A.; Samanta, M.K.; Gayen, S.; Maiti, M.K. The sucrose non-fermenting 1-related kinase 2 gene SAPK9 improves drought tolerance and grain yield in rice by modulating cellular osmotic potential, stomatal closure and stress-responsive gene expres-sion. BMC Plant Biol. 2016, 16, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, Y.; Murata, M.; Minami, H.; Yamamoto, S.; Kagaya, Y.; Hobo, T.; Yamamoto, A.; Hattori, T. Abscisic acid-activated SNRK2 protein kinases function in the gene-regulation pathway of ABA signal transduction by phosphorylating ABA response element-binding factors. Plant J. 2005, 44, 939–949. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Fujita, Y.; Kanamori, N.; Katagiri, T.; Umezawa, T.; Kidokoro, S.; Maruyama, K.; Yoshida, T.; Ishiyama, K.; Kobayashi, M.; et al. Three Arabidopsis SnRK2 protein kinases, SRK2D/SnRK2.2, SRK2E/SnRK2.6/OST1 and SRK2I/SnRK2.3, involved in ABA signaling are essential for the control of seed development and dormancy. Plant Cell Physiol. 2009, 50, 1345–1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, X.; Zhang, H.; Tian, S.; Chang, X.; Jing, R. TaSnRK2.4, an SNF1-type serine/threonine protein kinase of wheat (Triticum aestivum L.), confers enhanced multistress tolerance in Arabidopsis. J. Exp. Bot. 2010, 61, 683–696. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Mao, X.; Jing, R.; Chang, X.; Xie, H. Characterization of a common wheat (Triticum aestivum L.) TaSnRK2.7 gene involved in abiotic stress responses. J. Exp. Bot. 2011, 62, 975–988. [Google Scholar] [CrossRef] [Green Version]
- Tian, S.; Mao, X.; Zhang, H.; Chen, S.; Zha, C.; Yang, S.; Jing, R. Cloning and characterization of TaSnRK2.3, a novel SnRK2 gene in common wheat. J. Exp. Bot. 2013, 64, 2063–2080. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Yu, X.; Hori, C.; Demura, T.; Ohtani, M.; Zhuge, Q. Heterologous overexpression of poplar SnRK2 genes enhanced salt stress tolerance in Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 612. [Google Scholar] [CrossRef] [Green Version]
- Phan, T.T.; Sun, B.; Niu, J.Q.; Tan, Q.L.; Li, J.; Yang, L.T.; Li, Y.R. Overexpression of sugarcane gene SoSnRK2.1 confers drought tolerance in transgenic tobacco. Plant Cell Rep. 2016, 35, 1891–1905. [Google Scholar] [CrossRef]
- Lou, D.; Wang, H.; Yu, D. The sucrose non-fermenting-1-related protein kinases SAPK1 and SAPK2 function collaboratively as positive regulators of salt stress tolerance in rice. BMC Plant Biol. 2018, 18, 203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szymanska, K.P.; Polkowska-Kowalczyk, L.; Lichocka, M.; Maszkowska, J.; Dobrowolska, G. SNF1-Related Protein Kinases SnRK2.4 and SnRK2.10 Modulate ROS Homeostasis in Plant Response to Salt Stress. Int. J. Mol. Sci. 2019, 20, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, J.; Wang, L.; Wu, Y.; Luo, Q.; Zhang, Y.; Qiu, D.; Han, J.; Su, P.; Xiong, Z.; Chang, J.; et al. TaSnRK2.9, a Sucrose Non-fermenting 1-Related Protein Kinase Gene, Positively Regulates Plant Response to Drought and Salt Stress in Transgenic Tobacco. Front. Plant Sci. 2019, 9, 2003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, Y.; Huang, Y.; Xiong, L. Characterization of stress-responsive CIPK genes in rice for stress tolerance improvement. Plant Physiol. 2007, 144, 1416–1428. [Google Scholar] [CrossRef] [Green Version]
- Fujii, H.; Zhu, J.K. An autophosphorylation site of the protein kinase SOS2 is important for salt tolerance in Arabidopsis. Mol. Plant 2009, 2, 183–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, T.B.; Liu, J.; Wong, J.H.; Ye, X.; Sze, S.C.W.; Tong, Y.; Zhang, K.Y. Review of research on Dendrobium, a prized folk medicine. Appl. Microbiol. Biotechnol. 2012, 93, 1795–1803. [Google Scholar] [CrossRef]
- Yan, L.; Wang, X.; Liu, H.; Tian, Y.; Lian, J.; Yang, R.; Hao, S.; Wang, X.; Yang, S.; Li, Q.; et al. The Genome of Dendrobium officinale Illuminates the Biology of the Important Traditional Chinese Orchid Herb. Mol. Plant 2015, 8, 922–934. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.; Zhao, T.; Sheng, Y.; Zheng, T.; Fu, L.; Zhang, Y. Dendrobium officinale Kimura et Migo: A Review on Its Eth-nopharmacology, Phytochemistry, Pharmacology, and Industrialization. Evid. Based Complement. Alternat. Med. 2017, 2017, 7436259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, J.; Chatterjee, C.; Sengupta, A.; Gupta, K.; Gupta, B. Genome-wide analysis and evolutionary study of sucrose non-fermenting 1-related protein kinase 2 (SnRK2) gene family members in Arabidopsis and Oryza. Comput. Biol. Chem. 2014, 49, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Qin, Y.; Zou, Y.; Ma, F. Genome-wide identification and expression profiling of the SnRK2 gene family in Malus prunifolia. Gene 2014, 552, 87–97. [Google Scholar] [CrossRef]
- Wang, L.; Hu, W.; Sun, J.; Liang, X.; Yang, X.; Wei, S.; Wang, X.; Zhou, Y.; Xiao, Q.; Yang, G.; et al. Genome-wide analysis of SnRK gene family in Brachypodium distachyon and functional characterization of BdSnRK2.9. Plant Sci. 2015, 237, 33–45. [Google Scholar] [CrossRef]
- Zhu, W.; Wu, D.; Jiang, L.; Ye, L. Genome-wide identification and characterization of SnRK family genes in Brassica napus. BMC Plant Biol. 2020, 20, 287. [Google Scholar] [CrossRef]
- Zhang, G.Q.; Liu, K.W.; Li, Z.; Lohaus, R.; Hsiao, Y.Y.; Niu, S.C.; Wang, J.Y.; Lin, Y.C.; Xu, Q.; Chen, L.J.; et al. The Apostasia genome and the evolution of orchids. Nature 2017, 549, 379–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.H.; Wan, S.Q.; Wang, W.D.; Chen, J.F.; Huang, L.L.; Duan, M.S.; Yu, Y.B. Genome-wide identification and charac-terization of the CsSnRK2 family in Camellia sinensis. Plant Physiol. Biochem. 2018, 132, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Nong, Q.; Xie, J.; Wang, Z.; Liang, Q.; Solanki, M.K.; Malviya, M.K.; Liu, X.; Li, Y.; Htun, R.; et al. Molecular Charac-terization and Co-expression Analysis of the SnRK2 Gene Family in Sugarcane (Saccharum officinarum L.). Sci. Rep. 2017, 7, 17659. [Google Scholar] [CrossRef] [Green Version]
- Huai, J.; Wang, M.; He, J.; Zheng, J.; Dong, Z.; Lv, H.; Zhao, J.; Wang, G. Cloning and characterization of the SnRK2 gene family from Zea mays. Plant Cell Rep. 2008, 27, 1861–1868. [Google Scholar] [CrossRef]
- Yoo, M.J.; Ma, T.; Zhu, N.; Liu, L.; Harmon, A.C.; Wang, Q.; Chen, S. Genome-wide identification and homeolog-specific expression analysis of the SnRK2 genes in Brassica napus guard cells. Plant Mol. Biol. 2016, 91, 211–227. [Google Scholar] [CrossRef]
- Liu, Z.; Ge, X.; Yang, Z.; Zhang, C.; Zhao, G.; Chen, E.; Liu, J.; Zhang, X.; Li, F. Genome-wide identification and characterization of SnRK2 gene family in cotton (Gossypium hirsutum L.). BMC Genet. 2017, 18, 54. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Cheng, Y.H.; Zhang, C.; Shen, X.J.; You, Q.B.; Guo, W.; Li, X.; Song, X.J.; Zhou, X.A.; Jiao, Y.Q. Genome-Wide Identification and Characterization of the GmSnRK2 Family in Soybean. Int. J. Mol. Sci. 2017, 18, 1834. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.; Guo, X.; Zhao, D.; Zhang, Q.; Jiang, Y.; Wang, Y.; Peng, X.; Wei, Y.; Zhai, Z.; Zhao, W.; et al. Cloning and expression profiling of the PacSnRK2 and PacPP2C gene families during fruit development, ABA treatment, and dehydration stress in sweet cherry. Plant Physiol. Biochem. 2017, 119, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol. Plant 1962, 15, 437–497. [Google Scholar]
- Zhang, G.Q.; Xu, Q.; Bian, C.; Tsai, W.C.; Yeh, C.M.; Liu, K.W.; Yoshida, K.; Zhang, L.S.; Chang, S.B.; Chen, F.; et al. The Dendrobium catenatum Lindl. genome sequence provides insights into polysaccharide synthase, floral development and adaptive evolution. Sci. Rep. 2016, 6, 19029. [Google Scholar] [CrossRef] [Green Version]
- Finn, R.D.; Clements, J.; Arndt, W.; Miller, B.L.; Wheeler, T.J.; Schreiber, F.; Bateman, A.; Eddy, S.R. HMMER web server: 2015 update. Nucleic Acids Res. 2015, 43, W30–W38. [Google Scholar] [CrossRef]
- Wilkins, M.R.; Gasteiger, E.; Bairoch, A.; Sanchez, J.C.; Williams, K.L.; Appel, R.D.; Hochstrasser, D.F. Protein identification and analysis tools in the ExPASy server. Methods Mol. Biol. 1999, 112, 531–552. [Google Scholar] [PubMed]
- McWilliam, H.; Li, W.; Uludag, M.; Squizzato, S.; Park, Y.M.; Buso, N.; Cowley, A.P.; Lopez, R. Analysis Tool Web Services from the EMBL-EBI. Nucleic Acids Res. 2013, 41, W597–W600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Apweiler, R.; Attwood, T.K.; Bairoch, A.; Bateman, A.; Birney, E.; Biswas, M.; Bucher, P.; Cerutti, L.; Corpet, F.; Croning, M.D.; et al. The InterPro database, an integrated documentation resource for protein families, domains and functional sites. Nucleic Acids Res. 2001, 29, 37–40. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, A. Genomic Characterization and Expression Analysis of Basic Helix-Loop-Helix (bHLH) Family Genes in Traditional Chinese Herb Dendrobium officinale. Plants 2020, 8, 1044. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Scaffold | Length of Coding Sequences (CDS) (bp) | Protein Length (aa) | Mol. Wt. (kDa) | pI | Location | |
|---|---|---|---|---|---|---|---|---|
| DoSnRK1.1 | LOC110093905 | 737 | 1530 | 509 | 58.01 | 8.77 | a,b | |
| DoSnRK2.1 | LOC110104807 | 231 | 1086 | 361 | 41.6 | 5.81 | a,b | |
| DoSnRK2.2 | LOC110107470 | 1105 | 1080 | 359 | 41.39 | 5.58 | a,b | |
| DoSnRK2.3 | LOC110094403 | 209636 | 1164 | 387 | 44.66 | 5.17 | b,a | |
| DoSnRK2.4 | LOC110102148 | 211 | 1026 | 341 | 39.04 | 5.62 | a,b | |
| DoSnRK2.5 | LOC110092671 | 526 | 1005 | 334 | 37.87 | 5.69 | a,b | |
| DoSnRK2.6 | LOC110112577 | 459 | 1092 | 363 | 41.17 | 4.90 | a,c | |
| DoSnRK2.7 | LOC110116324 | 601 | 1086 | 361 | 40.92 | 4.77 | c,a | |
| DoSnRK3.1 | LOC110116107 | 456 | 1350 | 449 | 50.97 | 9.25 | b,c,a | |
| DoSnRK3.2 | LOC110099074 | 313 | 1347 | 448 | 50.97 | 9.07 | b,c,a | |
| DoSnRK3.3 | LOC110112518 | 1217 | 1389 | 462 | 52.43 | 9.08 | c,a | |
| DoSnRK3.4 | LOC110112938 | 61322 | 1305 | 434 | 49.76 | 9.18 | a,c,b | |
| DoSnRK3.5 | LOC110100634 | 1056 | 1323 | 440 | 50.23 | 9.16 | b,a,c | |
| DoSnRK3.6 | LOC110104808 | 231 | 1263 | 420 | 46.87 | 9.37 | c,a | |
| DoSnRK3.7 | LOC110113686 | 624 | 1341 | 446 | 49.92 | 9.37 | b,c,a | |
| DoSnRK3.8 | LOC110094439 | 629 | 1281 | 426 | 47.15 | 9.04 | a,b | |
| DoSnRK3.9 | LOC110093407 | 1038 | 1311 | 436 | 48.19 | 9.10 | d,c,b | |
| DoSnRK3.10 | LOC110095265 | 768 | 1416 | 471 | 53.46 | 8.79 | d | |
| DoSnRK3.11 | LOC110113865 | 370 | 1422 | 473 | 53.5 | 7.99 | d | |
| DoSnRK3.12 | LOC110109095 | 117410 | 1461 | 486 | 54.74 | 8.01 | b,a | |
| DoSnRK3.13 | LOC110093408 | 1038 | 1344 | 447 | 49.78 | 8.59 | d,c,b | |
| DoSnRK3.14 | LOC110095499 | 827 | 1290 | 429 | 48.2 | 9.32 | b,a | |
| DoSnRK3.15 | LOC110102687 | 123 | 1350 | 449 | 50.5 | 8.28 | b,a,c | |
| DoSnRK3.16 | LOC110098084 | 313 | 1419 | 472 | 52.82 | 9.30 | a,b,c | |
| DoSnRK3.17 | LOC110100633 | 1056 | 1467 | 488 | 54.69 | 9.10 | a,b,c | |
| DoSnRK3.18 | LOC110112951 | 61322 | 1371 | 456 | 50.94 | 8.80 | a,b,c | |
| DoSnRK3.19 | LOC110105714 | 260 | 1353 | 450 | 51.06 | 8.37 | a,b | |
| DoSnRK3.20 | LOC110098208 | 231 | 1254 | 417 | 47.59 | 5.91 | a,c | |
| DoSnRK3.21 | LOC110113559 | 1557 | 1350 | 449 | 51.09 | 8.01 | a | |
| DoSnRK3.22 | LOC110110059 | 1220 | 1299 | 432 | 48.82 | 8.94 | b,c,a | |
| DoSnRK3.23 | LOC110101878 | 974 | 1509 | 502 | 57.14 | 9.24 | b,c,a | |
| DoSnRK3.24 | LOC110103756 | 716 | 1293 | 430 | 48.82 | 8.87 | a,b,c | |
| DoSnRK3.25 | LOC110113526 | 1721 | 1356 | 451 | 51.47 | 8.70 | b,a,c | |
| DoSnRK3.26 | LOC110114482 | 1608 | 1305 | 434 | 49.36 | 8.32 | b,c,a | |
| DoSnRK3.27 | LOC110112680 | 627 | 1407 | 468 | 52.97 | 7.51 | b,c,a | |
| DoSnRK3.28 | LOC110107680 | 142 | 1299 | 432 | 48.78 | 8.01 | b,a,c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Liu, A. Genomic Characterization and Expression Analysis of the SnRK Family Genes in Dendrobium officinale Kimura et Migo (Orchidaceae). Plants 2021, 10, 479. https://doi.org/10.3390/plants10030479
Wang Y, Liu A. Genomic Characterization and Expression Analysis of the SnRK Family Genes in Dendrobium officinale Kimura et Migo (Orchidaceae). Plants. 2021; 10(3):479. https://doi.org/10.3390/plants10030479
Chicago/Turabian StyleWang, Yue, and Aizhong Liu. 2021. "Genomic Characterization and Expression Analysis of the SnRK Family Genes in Dendrobium officinale Kimura et Migo (Orchidaceae)" Plants 10, no. 3: 479. https://doi.org/10.3390/plants10030479
APA StyleWang, Y., & Liu, A. (2021). Genomic Characterization and Expression Analysis of the SnRK Family Genes in Dendrobium officinale Kimura et Migo (Orchidaceae). Plants, 10(3), 479. https://doi.org/10.3390/plants10030479