
Biostimulant Properties of Seaweed Extracts in Plants: Implications towards Sustainable Crop Production


Abstract
:1. Introduction
2. Seaweed Extracts—Methods of Preparation and Application into Plants
3. Effect of Seaweed Extracts on Plant Growth
4. Effect of Seaweed Extracts on Plants’ Tolerance to Biotic Stresses
5. Effect of Seaweed Extracts on Plants’ Tolerance to Abiotic and Environmental Stresses

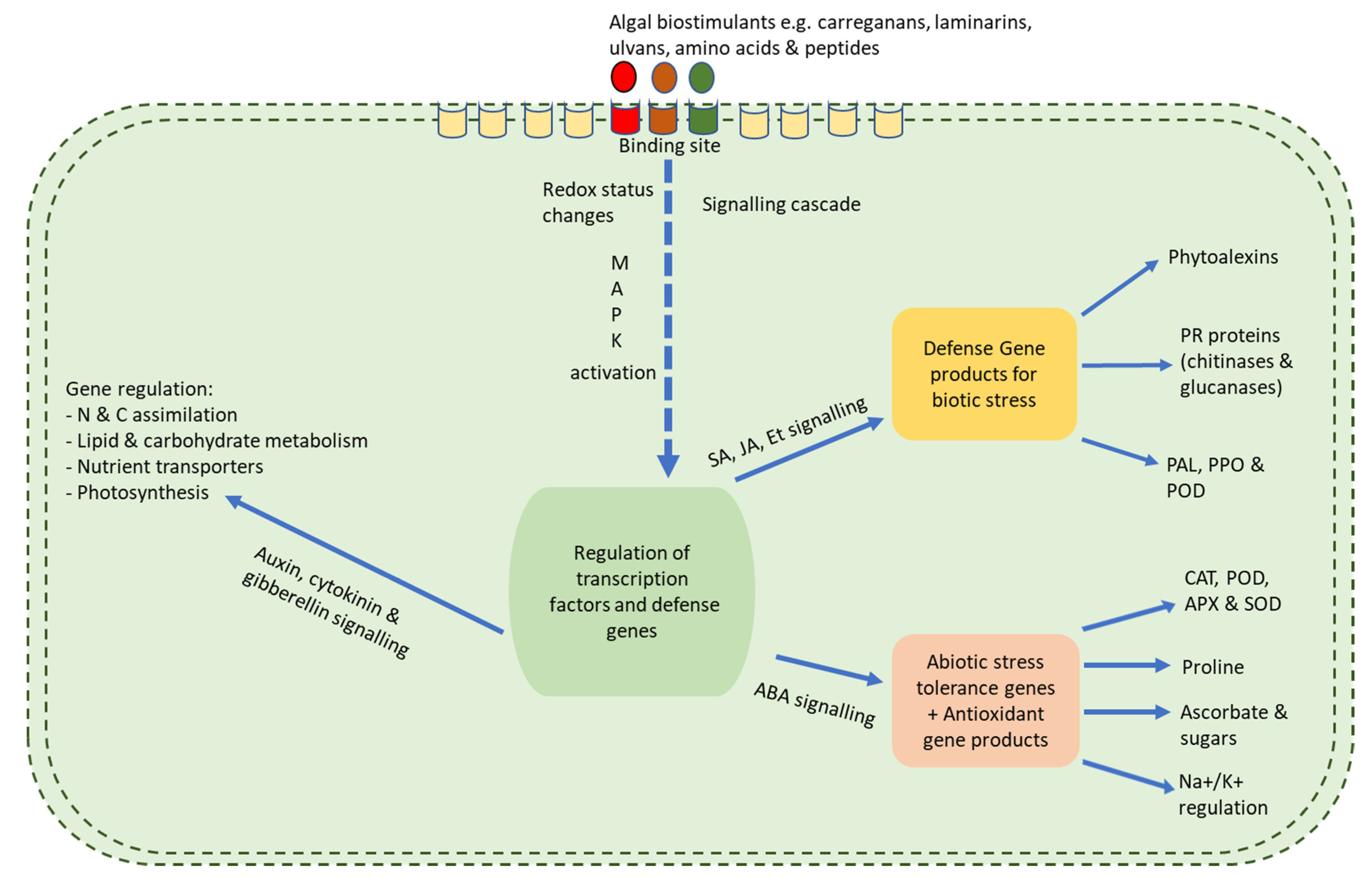{kind=link}
{kind=link}
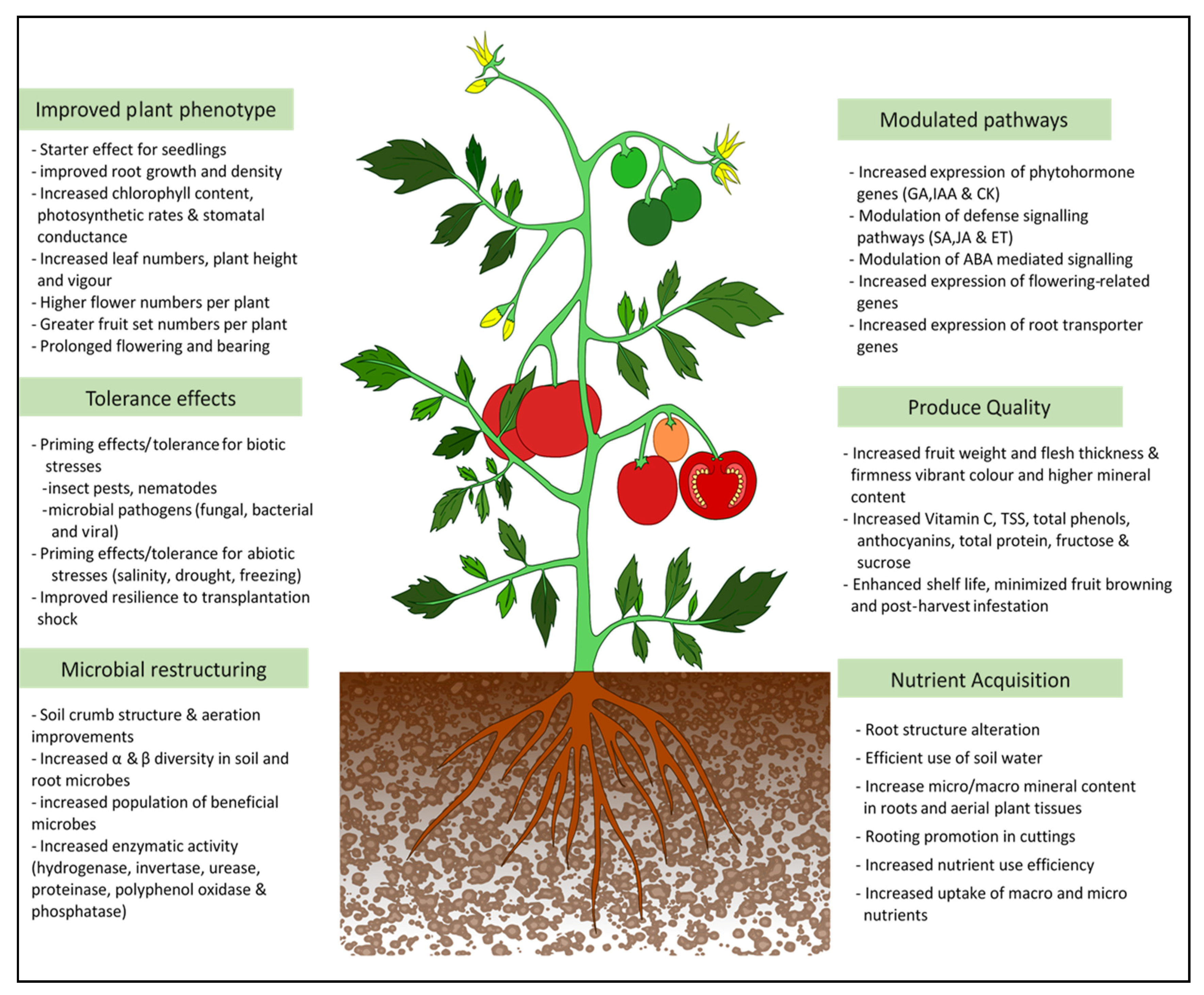{kind=link}
| Crop | Seaweed Extract | Observed Effects | Reference |
|---|---|---|---|
| Tomato (Solanum lycopersicum) |
|
| [3,11,31,32,62,63,64,65,66,67] |
| Sweet pepper (Capsicum annuum) |
|
| [3,11,60,61,63,68] |
| Lettuce (Lactuca sativa) |
|
| [37,68] |
| Cauliflower (Brassica oleracea) |
|
| [69] |
| Soybean (Glycine max) |
|
| [70,71,72] |
| Strawberry (Fragaria x ananassa) |
|
| [6,72,73,74,75] |
| Cucumber (Cucumis sativus) |
|
| [76,77,78] |
| Onion (Allium cepa) |
|
| [79,80,81,82] |
| Potato (Solanum tuberosum) |
|
| [83,84,85,86,87] |
| Broccoli (Brassica oleracea var. italica) |
|
| [87,88] |
| Spinach (S. oleracea) |
|
| [34,89,90,91] |
| Carrot (Daucus carota) |
|
| [92,93,94,95,96] |
| Wheat (Triticum aestivum) |
|
| [43,97,98,99,100,101,102] |
| Rice (Oryza sativa) |
|
| [103,104,105,106] |
| Apple (Malus domestica) |
|
| [39,107,108,109,110] |
| Maize (Zea mays) |
|
| [22,111,112,113,114,115] |
| Orange (Citrus spp.) |
|
| [14,49,116,117,118] |
| Sugarcane (Saccharum officinarum) |
|
| [53,118,119] |
6. Mechanisms and Mode of Biostimulatory Activities

7. Effect of Seaweed Extracts on the Plant and Soil Microbiome Dynamics
8. Bioactive and Elicitor Components of Seaweed Extracts

| Bioactive Compounds | Chlorophyceae (Green) | Rhodophyceae (Red) | Phaeophyceae (Brown) | Reference |
|---|---|---|---|---|
| Polysaccharides |
|
|
| [55,137] |
| Plant Growth Regulators |
|
|
| [16,146,147] |
| Betaines |
|
|
| [148,149] |
| Sterols |
|
|
| [150,151,152,153] |
| Carotenoid |
|
|
| [154,155] |
| Minerals |
|
|
| [156,157,158] |
| Polyphenolics and Phlorotannins |
|
|
| [159,160] |
| Lipids |
|
|
| [153,161,162] |
| Oxylipins |
|
|
| [151,160,161,162,163] |
| Protein, peptides, and amino acids |
|
|
| [164,165,166] |
9. Seaweed Extract as an Input for Integrated Crop Management Program: A Paradigm Shift towards Sustainable Agriculture
10. Challenges and Opportunities for Seaweed Biomass Valorization and Development of Novel Agricultural Inputs
11. Conclusions

Funding
Conflicts of Interest
References
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in Plant Science: A Global Perspective. Front. Plant Sci. 2017, 54, 23–27. [Google Scholar] [CrossRef] [Green Version]
- Du Jardin, P. The Science of Plant Biostimulants–A Bibliographic Analysis, Ad Hoc Study Report; European Commission: Brussels, Belgium, 2012. [Google Scholar]
- Khan, W.; Rayirath, U.P.; Subramanian, S.; Jithesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craigie, J.S.; Norrie, J.; Prithiviraj, B. Seaweed Extracts as Biostimulants of Plant Growth and Development. J. Plant Growth Regul. 2009, 45, 112–134. [Google Scholar] [CrossRef]
- Du Jardin, P. Plant Biostimulants: Definition, Concept, Main Categories and Regulation. Sci. Hortic. (Amst. ) 2015, 53, 3555–3654. [Google Scholar] [CrossRef] [Green Version]
- Ali, O.; Ramsubhag, A.; Jayaraman, J. Biostimulatory Activities of Ascophyllum nodosum Extract in Tomato and Sweet Pepper Crops in a Tropical Environment. PLoS ONE 2019, 14, e0216710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapur, B.; Sarıdaş, M.A.; Çeliktopuz, E.; Kafkas, E.; Paydaş Kargı, S. Health and Taste Related Compounds in Strawberries under Various Irrigation Regimes and Bio-Stimulant Application. Food Chem. 2018. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Mattson, N.S. Effects of Seaweed Extract Application Rate and Method on Post-Production Life of Petunia and Tomato Transplants. Horttechnology 2015. [Google Scholar] [CrossRef] [Green Version]
- Parađiković, N.; Teklić, T.; Zeljković, S.; Lisjak, M.; Špoljarević, M. Biostimulants Research in Some Horticultural Plant Species—A Review. Food Energy Secur. 2019. [Google Scholar] [CrossRef]
- Shukla, P.S.; Mantin, E.G.; Adil, M.; Bajpai, S.; Critchley, A.T.; Prithiviraj, B. Ascophyllum nodosum -Based Biostimulants: Sustainable Applications in Agriculture for the Stimulation of Plant Growth, Stress Tolerance, and Disease Management. Front. Plant Sci. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayaraj, J. Induced Resistance-a Sustainable Biotechnological Approach of Plant Disease Management. In Microbial Antagonists: Their Role in Biological Control of Plant Diseases; Today & Tomorrow’s Printers and Publishers: New Delhi, India, 2018; pp. 770–777. [Google Scholar]
- Jayaraman, J.; Ali, N. Use of Seaweed Extracts for Disease Management of Vegetable Crops. In Sustainable Crop Disease Management Using Natural Products; CABI: Wallingford, UK, 2015; pp. 160–173. [Google Scholar]
- Ali, N.; Farrell, A.; Ramsubhag, A.; Jayaraman, J. The Effect of Ascophyllum nodosum Extract on the Growth, Yield and Fruit Quality of Tomato Grown under Tropical Conditions. J. Appl. Phycol. 2016, 28, 1353–1362. [Google Scholar] [CrossRef]
- Ali, O.; Ramsubhag, A.; Jayaraman, J. Phytoelicitor Activity of Sargassum vulgare and Acanthophora spicifera Extracts and Their Prospects for Use in Vegetable Crops for Sustainable Crop Production. J. Appl. Phycol. 2020. [Google Scholar] [CrossRef]
- Arioli, T.; Mattner, S.W.; Winberg, P.C. Applications of Seaweed Extracts in Australian Agriculture: Past, Present and Future. J. Appl. Phycol. 2015, 27, 2007–2015. [Google Scholar] [CrossRef] [Green Version]
- Rayorath, P.; Khan, W.; Palanisamy, R.; Mackinnon, S.L.; Stefanova, R.; Hankins, S.D.; Critchley, A.T.; Prithiviraj, B. Extracts of the Brown Seaweed Ascophyllum nodosum Induce Gibberellic Acid (GA3)-Independent Amylase Activity in Barley. J. Plant Growth Regul. 2008, 32, 123–128. [Google Scholar] [CrossRef]
- Crouch, I.J.; van Staden, J. Evidence for the Presence of Plant Growth Regulators in Commercial Seaweed Products. Plant Growth Regul. 1993, 6, 345–388. [Google Scholar] [CrossRef]
- Aldworth, S.J.; van Staden, J. The Effect of Seaweed Concentrate on Seedling Transplants. S. Afr. J. Bot. 1987. [Google Scholar] [CrossRef] [Green Version]
- Crouch, I.J.; Smith, M.T.; van Staden, J.; Lewis, M.J.; Hoad, G.V. Identification of Auxins in a Commercial Seaweed Concentrate. J. Plant Physiol. 1992. [Google Scholar] [CrossRef]
- Crouch, I.J.; Van Staden, J. Evidence for Rooting Factors in a Seaweed Concentrate Prepared from Ecklonia maxima. J. Plant Physiol. 1991. [Google Scholar] [CrossRef]
- Atzmon, N.; Van Staden, J. The Effect of Seaweed Concentrate on the Growth of Pinus pinea Seedlings. New For. 1994. [Google Scholar] [CrossRef]
- Leclerc, M.; Caldwell, C.D.; Lada, R.R.; Norrie, J. Effect of Plant Growth Regulators on Propagule Formation in Hemerocallis spp. and Hosta spp. HortScience 2006. [Google Scholar] [CrossRef] [Green Version]
- Crouch, I.J.; Beckett, R.P.; van Staden, J. Effect of Seaweed Concentrate on the Growth and Mineral Nutrition of Nutrient-Stressed Lettuce. J. Appl. Phycol. 1990. [Google Scholar] [CrossRef]
- Senthuran, S.; Balasooriya, B.L.W.K.; Arasakesary, S.J.; Gnanavelrajah, N. Effect of Seaweed Extract Kappaphycus alvarezii on the Growth, Yield and Nutrient Uptake of Leafy Vegetable Amaranthus Polygamous. Trop. Agric. Res. 2019. [Google Scholar] [CrossRef] [Green Version]
- Ertani, A.; Francioso, O.; Tinti, A.; Schiavon, M.; Pizzeghello, D.; Nardi, S. Evaluation of Seaweed Extracts from Laminaria and Ascophyllum nodosum Spp. As Biostimulants in Zea mays L. Using a Combination of Chemical, Biochemical and Morphological Approaches. Front. Plant Sci. 2018. [Google Scholar] [CrossRef] [PubMed]
- Fei, H.; Crouse, M.; Papadopoulos, Y.; Vessey, J.K. Enhancing the Productivity of Hybrid Poplar (Populus × Hybrid) and Switchgrass (Panicum virgatum L.) by the Application of Beneficial Soil Microbes and a Seaweed Extract. Biomass Bioenergy 2017. [Google Scholar] [CrossRef]
- Di Stasio, E.; Rouphael, Y.; Colla, G.; Raimondi, G.; Giordano, M.; Pannico, A.; El-Nakhel, C.; De Pascale, S. The Influence of Ecklonia maxima Seaweed Extract on Growth, Photosynthetic Activity and Mineral Composition of Brassica rapa L. ssp. sylvestris under Nutrient Stress Conditions. Eur. J. Hortic. Sci. 2017. [Google Scholar] [CrossRef]
- Werner, T.; Motyka, V.; Strnad, M.; Schmülling, T. Regulation of Plant Growth by Cytokinin. Proc. Natl. Acad. Sci. USA 2001. [Google Scholar] [CrossRef] [Green Version]
- Blunden, G.; Jenkins, T.; Liu, Y.W. Enhanced Leaf Chlorophyll Levels in Plants Treated with Seaweed Extract. J. Appl. Phycol. 1996, 3, 13–19. [Google Scholar] [CrossRef]
- Whapham, C.A.; Blunden, G.; Jenkins, T.; Hankins, S.D. Significance of Betaines in the Increased Chlorophyll Content of Plants Treated with Seaweed Extract. J. Appl. Phycol. 1993. [Google Scholar] [CrossRef]
- Genard, H.; Le Saos, J.; Billard, J.; Tremolieres, A.; Boucaud, J. Effect of Salinity on Lipid Composition, Glycine Betaine Content and Photosynthetic Activity in Chloroplasts of Suaeda maritima. Plant Physiol. Biochem. 1991, 29, 421–427. [Google Scholar]
- Al-Ghamdi, A.A.; Elansary, H.O. Synergetic Effects of 5-Aminolevulinic Acid and Ascophyllum nodosum Seaweed Extracts on Asparagus Phenolics and Stress Related Genes under Saline Irrigation. Plant Physiol. Biochem. 2018. [Google Scholar] [CrossRef] [PubMed]
- Digruber, T.; Sass, L.; Cseri, A.; Paul, K.; Nagy, A.V.; Remenyik, J.; Molnár, I.; Vass, I.; Toldi, O.; Gyuricza, C.; et al. Stimulation of Energy Willow Biomass with Triacontanol and Seaweed Extract. Ind. Crop. Prod. 2018. [Google Scholar] [CrossRef]
- Ramkissoon, A.; Ramsubhag, A.; Jayaraman, J. Phytoelicitor Activity of Three Caribbean Seaweed Species on Suppression of Pathogenic Infections in Tomato Plants. J. Appl. Phycol. 2017, 34, 123–129. [Google Scholar] [CrossRef]
- Dookie, M.; Ali, O.; Ramsubhag, A.; Jayaraman, J. Flowering Gene Regulation in Tomato Plants Treated with Brown Seaweed Extracts. Sci. Hortic. (Amst. ) 2020. [Google Scholar] [CrossRef]
- El-Yazied, A.; El-Gizawy, A.M.; Ragab, M.I.; Hamed, E.S. Effect of Seaweed Extract and Compost Treatments on Growth, Yield and Quality of Snap Bean. J. Am. Sci. 2012, 8, 1–20. [Google Scholar]
- Kulkarni, M.G.; Rengasamy, K.R.R.; Pendota, S.C.; Gruz, J.; Plačková, L.; Novák, O.; Doležal, K.; Van Staden, J. Bioactive Molecules Derived from Smoke and Seaweed Ecklonia maxima Showing Phytohormone-like Activity in Spinacia oleracea L. N. Biotechnol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Yusuf, R.; Kristianse, P.; Warwick, N. Effect of Two Seaweed Products and Equivalent Mineral Treatments on Lettuce (Lactuca sativa L.) Growth. J. Agron. 2019. [Google Scholar] [CrossRef]
- Xu, C.; Leskovar, D.I. Effects of A. nodosum Seaweed Extracts on Spinach Growth, Physiology and Nutrition Value under Drought Stress. Sci. Hortic. (Amst.) 2015, 183, 39–47. [Google Scholar] [CrossRef]
- Valencia, R.T.; Acosta, L.S.; Hernández, M.F.; Rangel, P.P.; Gallegos Robles, M.Á.; del Carmen Antonio Cruz, R.; Vázquez, C.V. Effect of Seaweed Aqueous Extracts and Compost on Vegetative Growth, Yield, and Nutraceutical Quality of Cucumber (Cucumis sativus L.) Fruit. Agronomy 2018, 8, 264. [Google Scholar] [CrossRef] [Green Version]
- Frioni, T.; Sabbatini, P.; Tombesi, S.; Norrie, J.; Poni, S.; Gatti, M.; Palliotti, A. Effects of a Biostimulant Derived from the Brown Seaweed Ascophyllum nodosum on Ripening Dynamics and Fruit Quality of Grapevines. Sci. Hortic. (Amst.) 2018. [Google Scholar] [CrossRef]
- Augusto, A.; Simões, T.; Pedrosa, R.; Silva, S.F.J. Evaluation of Seaweed Extracts Functionality as Post-Harvest Treatment for Minimally Processed Fuji Apples. Innov. Food Sci. Emerg. Technol. 2016. [Google Scholar] [CrossRef]
- Finnie, J.F.; van Staden, J. Effect of Seaweed Concentrate and Applied Hormones on In Vitro Cultured Tomato Roots. J. Plant Physiol. 1985. [Google Scholar] [CrossRef]
- Beckett, R.P.; van Staden, J. The Effect of Seaweed Concentrate on the Growth and Yield of Potassium Stressed Wheat. Plant Soil 1989, 53, 187–189. [Google Scholar] [CrossRef]
- Vernieri, P.; Borghesi, E.; Tognoni, F.; Serra, G.; Ferrante, A.; Piaggesi, A. Use of Biostimulants for Reducing Nutrient Solution Concentration in Floating System. Acta Hortic. 2006, 21, 123–134. [Google Scholar] [CrossRef]
- Mancuso, S.; Azzarello, E.; Mugnai, S.; Briand, X. Marine Bioactive Substances (IPA Extract) Improve Foliar Ion Uptake and Water Stress Tolerance in Potted Vitis vinifera Plants. Adv. Hortic. Sci. 2006. [Google Scholar] [CrossRef]
- Ayliffe, M.A.; Lagudah, E.S. Molecular Genetics of Disease Resistance in Cereals. Ann. Bot. 2004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, P.K.; Cunningham, A.A.; Patel, N.G.; Morales, F.J.; Epstein, P.R.; Daszak, P. Emerging Infectious Diseases of Plants: Pathogen Pollution, Climate Change and Agrotechnology Drivers. Trends Ecol. Evol. 2004. [Google Scholar] [CrossRef]
- De Waele, D.; Mcdonald, H.; De Waele, E. Influence of Seaweed Concentrate on the Reproduction of Pratylenchuszeae (Nematoda) on Maize. Nematologica 1988. [Google Scholar] [CrossRef]
- Asaraja, A.; Sahayaraj, K. Screening of Insecticidal Activity of Brown Macroalgal Extracts against Dysdercus cingulatus (Fab.) (Hemiptera: Pyrrhocoridae). J. Biopestic. 2013, 6, 193–203. [Google Scholar]
- Gençsoylu, İ. Effect Of Seaweeds And Organic Foliar Fertilizers On The Cotton Pests, Predators, Yield And Fiber Quality In Cotton. Adnan Menderes Üniv. Ziraat Fakültesi Derg. 2016. [Google Scholar] [CrossRef] [Green Version]
- González-Castro, A.L.; Muñoz-Ochoa, M.; Hernández-Carmona, G.; López-Vivas, J.M. Evaluation of Seaweed Extracts for the Control of the Asian Citrus Psyllid Diaphorina citri. J. Appl. Phycol. 2019. [Google Scholar] [CrossRef]
- Carolina Feitosa de Vasconcelos, A.; Helena Garófalo Chaves, L. Biostimulants and Their Role in Improving Plant Growth under Abiotic Stresses. In Biostimulants in Plant Science; IntechOpen: London, UK, 2019. [Google Scholar]
- Chen, D.; Huang, Y.; Shen, D.; Zhou, W.; Ao, J.; Jiang, Y.; Huang, Z.; Hassan, M.; Fahd, R. Effects of Seaweed Extracts on Promoting Growth and Improving Stress Resistance in Sugarcane. Asian Agric. Res. 2019, 69–76. [Google Scholar] [CrossRef]
- Kubo, I.; Matsumoto, T.; Ichikawa, N. Absolute Configuration Of Crinitol. An Acyclic Diterpene Insect Growth Inhibitor From The Brown Algae Sargassum tortile. Chem. Lett. 1985. [Google Scholar] [CrossRef]
- Vera, J.; Castro, J.; Gonzalez, A.; Moenne, A. Seaweed Polysaccharides and Derived Oligosaccharides Stimulate Defense Responses and Protection against Pathogens in Plants. Mar. Drugs 2011, 44, 2514–2525. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, E.; Dorta, F.; Medina, C.; Ramírez, A.; Ramírez, I.; Peña-Cortés, H. Anti-Phytopathogenic Activities of Macro-Algae Extracts. Mar. Drugs 2011, 9, 739–756. [Google Scholar] [CrossRef] [Green Version]
- Klarzynski, O.; Descamps, V.; Plesse, B.; Yvin, J.C.; Kloareg, B.; Fritig, B. Sulfated Fucan Oligosaccharides Elicit Defense Responses in Tobacco and Local and Systemic Resistance against Tobacco Mosaic Virus. Mol. Plant -Microbe Interact. 2003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, S. Frost Management in Cool Climate Vineyards; Final report to Wine Australia project number: UT 99/1; University of Tasmania: Hobart, Australia, 2001. [Google Scholar]
- Patel, K.; Agarwal, P.; Agarwal, P.K. Kappaphycus alvarezii Sap Mitigates Abiotic-Induced Stress in Triticum durum by Modulating Metabolic Coordination and Improves Growth and Yield. J. Appl. Phycol. 2018. [Google Scholar] [CrossRef]
- Dalal, A.; Bourstein, R.; Haish, N.; Shenhar, I.; Wallach, R.; Moshelion, M. Dynamic Physiological Phenotyping of Drought-Stressed Pepper Plants Treated with “Productivity-Enhancing” and “Survivability-Enhancing” Biostimulants. Front. Plant Sci. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganesan, S.; Vadivel, K.; Jayaraman, J. Sustainable Crop Disease Management Using Natural Products; CABI: Wallingford, UK, 2015. [Google Scholar] [CrossRef]
- Ali, O.; Ramsubhag, A.; Jayaraman, J. Ascophyllum nodosum ( Linnaeus ) Le Jolis Seaweed Extract Improves Seed Germination in Tomato and Sweet Pepper under NaCl-Induced Salt Stress. Trop. Agric. 2018, 95, 141–148. [Google Scholar]
- Vinoth, S.; Sundari; Gurusaravanan, P.; Sivakumar, S.; Siva, G.; Kumar, G.P.; Manju; Velmurugan, K.; Lakshminarayana, V.; Jayabalan, N. Evaluation of Seagrass Liquid Extract on Salt Stress Alleviation in Tomato Plants. Asian J. Plant Sci. 2017. [Google Scholar] [CrossRef] [Green Version]
- Murtic, S.; Oljaca, R.; Smajic Murtic, M.; Vranac, A.; Akagic, A.; Civic, H. Cherry Tomato Productivity as Influenced by Liquid Organic Fertilizer under Different Growth Conditions. J. Cent. Eur. Agric. 2018. [Google Scholar] [CrossRef]
- Esserti, S.; Smaili, A.; Rifai, L.A.; Koussa, T.; Makroum, K.; Belfaiza, M.; Kabil, E.M.; Faize, L.; Burgos, L.; Alburquerque, N.; et al. Protective Effect of Three Brown Seaweed Extracts against Fungal and Bacterial Diseases of Tomato. J. Appl. Phycol. 2017. [Google Scholar] [CrossRef]
- Demir, N.; Dural, B.; Yildirim, K. Effect of Seaweed Suspensions on Seed Germination of Tomato, Pepper and Aubergine. J. Biol. Sci. 2006, 1130–1133. [Google Scholar] [CrossRef] [Green Version]
- Paradiković, N.; Vinković, T.; Vinković Vrček, I.; Žuntar, I.; Bojić, M.; Medić-Šarić, M. Effect of Natural Biostimulants on Yield and Nutritional Quality: An Example of Sweet Yellow Pepper (Capsicum annuum L.) Plants. J. Sci. Food Agric. 2011. [Google Scholar] [CrossRef]
- Di Mola, I.; Cozzolino, E.; Ottaiano, L.; Giordano, M.; Rouphael, Y.; Colla, G.; Mori, M. Effect of Vegetal- And Seaweed Extract-Based Biostimulants on Agronomical and Leaf Quality Traits of Plastic Tunnel-Grown Baby Lettuce under Four Regimes of Nitrogen Fertilization. Agronomy 2019, 9, 571. [Google Scholar] [CrossRef] [Green Version]
- Abetz, P.; Young, C.L. The Effect of Seaweed Extract Sprays Derived from Ascophyllum nodosum on Lettuce and Cauliflower Crops. Bot. Mar. 1983. [Google Scholar] [CrossRef]
- Shukla, P.S.; Shotton, K.; Norman, E.; Neily, W.; Critchley, A.T.; Prithiviraj, B. Seaweed Extract Improve Drought Tolerance of Soybean by Regulating Stress-Response Genes. AoB Plants 2018. [Google Scholar] [CrossRef] [PubMed]
- Martynenko, A.; Shotton, K.; Astatkie, T.; Petrash, G.; Fowler, C.; Neily, W.; Critchley, A.T. Thermal Imaging of Soybean Response to Drought Stress: The Effect of Ascophyllum nodosum Seaweed Extract. Springerplus 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meszka, B.; Bielenin, A. Bioproducts in Control of Strawberry verticillium wilt. Phytopathologia 2008, 52, 21–27. [Google Scholar]
- Washington, W.S.; Engleitner, S.; Boontjes, G.; Shanmuganathan, N. Effect of Fungicides, Seaweed Extracts, Tea Tree Oil, and Fungal Agents on Fruit Rot and Yield in Strawberry. Aust. J. Exp. Agric. 1999. [Google Scholar] [CrossRef]
- El-Miniawy, S.M.; Ragab, M.E.; Youssef, S.M.; Metwally, A.A. Influence of Foliar Spraying of Seaweed Extract on Growth, Yield and Quality of Strawberry. J. App. Sci. Res. 2014, 10, 88–94. [Google Scholar]
- Mattner, S.W.; Milinkovic, M.; Arioli, T. Increased Growth Response of Strawberry Roots to a Commercial Extract from Durvillaea potatorum and Ascophyllum nodosum. J. Appl. Phycol. 2018. [Google Scholar] [CrossRef] [Green Version]
- Jayaraman, J.; Norrie, J.; Punja, Z.K. Commercial Extract from the Brown Seaweed Ascophyllum nodosum Reduces Fungal Diseases in Greenhouse Cucumber. J. Appl. Phycol. 2011, 15, 145–155. [Google Scholar] [CrossRef]
- Jaulneau, V.; Lafitte, C.; Corio-Costet, M.F.; Stadnik, M.J.; Salamagne, S.; Briand, X.; Esquerré-Tugayé, M.T.; Dumas, B. An Ulva Armoricana Extract Protects Plants against Three Powdery Mildew Pathogens. Eur. J. Plant Pathol. 2011, 131, 393–401. [Google Scholar] [CrossRef]
- Hidangmayum, A.; Sharma, R. Effect of Different Concentration of Commercial Seaweed Liquid Extract of Ascophylum nodosum on Germination of Onion (Allium cepa L.). Int. J. Sci. Res. 2017. [Google Scholar] [CrossRef]
- Hidangmayum, A.; Sharma, R. Effect of Different Concentrations of Commercial Seaweed Liquid Extract of Ascophyllum nodosum as a Plant Bio Stimulant on Growth, Yield and Biochemical Constituents of Onion (Allium cepa L.). J. Pharmacogn. Phytochem. 2017, 6, 658–663. [Google Scholar]
- Abbas, M.; Anwar, J.; Zafar-Ul-Hye, M.; Khan, R.I.; Saleem, M.; Rahi, A.A.; Danish, S.; Datta, R. Effect of Seaweed Extract on Productivity and Quality Attributes of Four Onion Cultivars. Horticulturae 2020, 6, 28. [Google Scholar] [CrossRef]
- Almaroai, Y.A.; Eissa, M.A. Role of Marine Algae Extracts in Water Stress Resistance of Onion Under Semiarid Conditions. J. Soil Sci. Plant Nutr. 2020. [Google Scholar] [CrossRef]
- Haider, M.W.; Ayyub, C.M.; Pervez, M.A.; Asad, H.U.; Manan, A.; Raza, S.A.; Ashraf, I. Impact of Foliar Application of Seaweed Extract on Growth, Yield and Quality of Potato (Solanum tuberosum L.). Soil Environ. 2012, 31, 157–162. [Google Scholar]
- Blunden, G.; Wildgoose, P.B. The Effects of Aqueous Seaweed Extract and Kinetin on Potato Yields. J. Sci. Food Agric. 1977. [Google Scholar] [CrossRef]
- Dwelle, R.B.; Hurley, P.J. The Effects of Foliar Application of Cytokinins on Potato Yields in Southeastern Idaho. Am. Potato J. 1984. [Google Scholar] [CrossRef]
- Prajapati, A.; Patel, C.K.; Singh, N.; Jain, S.K.; Chongtham, S.K.; Maheshwari, M.N.; Patel, C.R.; Pate, R.N. Evaluation of Seaweed Extract on Growth and Yield of Potato. Environ. Ecol. 2016, 34, 605–608. [Google Scholar]
- Dziugieł, T.; Wadas, W. Possibility of Increasing Early Crop Potato Yield with Foliar Application of Seaweed Extracts and Humic Acids. J. Cent. Eur. Agric. 2020. [Google Scholar] [CrossRef]
- Mattner, S.W.; Wite, D.; Riches, D.A.; Porter, I.J.; Arioli, T. The Effect of Kelp Extract on Seedling Establishment of Broccoli on Contrasting Soil Types in Southern Victoria, Australia. Biol. Agric. Hortic. 2013. [Google Scholar] [CrossRef]
- Gajc-Wolska, J.; Spiewski, T.; Grabowska, A. The Effect of Seaweed Extracts on the Yield and Quality Parameters of Broccoli (Brassica oleracea var. cymosa L.) in Open Field Production. Acta Hortic. 2012. [Google Scholar] [CrossRef]
- Rouphael, Y.; Giordano, M.; Cardarelli, M.; Cozzolino, E.; Mori, M.; Kyriacou, M.C.; Bonini, P.; Colla, G. Plant-and Seaweed-Based Extracts Increase Yield but Differentially Modulate Nutritional Quality of Greenhouse Spinach through Biostimulant Action. Agronomy 2018, 8, 126. [Google Scholar] [CrossRef] [Green Version]
- Zahid, P.B. Preparation of Organic Fertilizer from Seaweed and Its Effect on the Growth of Some Vegetable and Ornamental Plants. Pakistan J. Biol. Sci. 1999. [Google Scholar] [CrossRef]
- Poberezny, J.; Szczepanek, M.; Wszelaczynska, E.; Prus, P. The Quality of Carrot after Field Biostimulant Application and after Storage. Sustainability 2020, 12, 1386. [Google Scholar] [CrossRef] [Green Version]
- Kiraci, S. Effects of Seaweed and Different Farm Manures on Growth and Yield of Organic Carrots. J. Plant Nutr. 2018. [Google Scholar] [CrossRef]
- Alam, M.Z.; Braun, G.; Norrie, J.; Hodges, D.M. Ascophyllum Extract Application Can Promote Plant Growth and Root Yield in Carrot Associated with Increased Root-Zone Soil Microbial Activity. Can. J. Plant Sci. 2014. [Google Scholar] [CrossRef] [Green Version]
- Jayaraj, J.; Wan, A.; Rahman, M.; Punja, Z.K. Seaweed Extract Reduces Foliar Fungal Diseases on Carrot. Crop Prot. 2008, 44, 345–366. [Google Scholar] [CrossRef]
- Eggen, T.; Asp, T.N.; Grave, K.; Hormazabal, V. Uptake and Translocation of Metformin, Ciprofloxacin and Narasin in Forage- and Crop Plants. Chemosphere 2011. [Google Scholar] [CrossRef] [PubMed]
- Kasim, W.A.; Hamada, E.A.M.; Shams El-Din, N.G.; Eskander, S.K. Influence of Seaweed Extracts on the Growth, Some Metabolic Activities and Yield of Wheat Grown under Drought Stress. Int. J. Agron. Agric. Res. 2015, 7, 173–189. [Google Scholar]
- Sharma, S.; Chen, C.; Khatri, K.; Rathore, M.S.; Pandey, S.P. Gracilaria Dura Extract Confers Drought Tolerance in Wheat by Modulating Abscisic Acid Homeostasis. Plant Physiol. Biochem. 2019. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.T.; Zodape, S.T.; Chaudhary, D.R.; Eswaran, K.; Chikara, J. Seaweed Sap As An Alternative Liquid Fertilizer For Yield And Quality Improvement Of Wheat. J. Plant Nutr. 2013. [Google Scholar] [CrossRef]
- Salim, B.B.M. Influence of Biochar and Seaweed Extract Applications on Growth, Yield and Mineral Composition of Wheat (Triticum aestivum L.) under Sandy Soil Conditions. Ann. Agric. Sci. 2016. [Google Scholar] [CrossRef] [Green Version]
- Amaral Carvalho, M.E.; de Camargo e Castro, P.R.; Gallo, L.A.; de Castro Ferraz Junior, M.V. Seaweed Extract Provides Development and Production of Wheat. Rev. Agrar. 2014, 7, 166–170. [Google Scholar]
- Carolina Feitosa de Vasconcelos, A. Amelioration of Drought Stress on Plants under Biostimulant Sources. Plant Stress Physiol. 2020. [Google Scholar] [CrossRef]
- Begum, M.; Bordoloi, B.C.; Singha, D.D.; Ojha, N.J. Role of Seaweed Extract on Growth, Yield and Quality of Some Agricultural Crops: A Review. Agric. Rev. 2018. [Google Scholar] [CrossRef]
- Pramanick, B.; Brahmachari, K.; Ghosh, A.; Zodape, S.T. Effect of Seaweed Saps on Growth and Yield Improvement of Transplanted Rice in Old Alluvial Soil of West Bengal. Bangladesh J. Bot. 2014. [Google Scholar] [CrossRef] [Green Version]
- Effect of Seaweed Extracts on Growth and Yield of Rice Plants. Nusant. Biosci. 2010. [CrossRef]
- Layek, J.; Das, A.; Idapuganti, R.G.; Sarkar, D.; Ghosh, A.; Zodape, S.T.; Lal, R.; Yadav, G.S.; Panwar, A.S.; Ngachan, S.; et al. Seaweed Extract as Organic Bio-Stimulant Improves Productivity and Quality of Rice in Eastern Himalayas. J. Appl. Phycol. 2018. [Google Scholar] [CrossRef]
- Spinelli, F.; Fiori, G.; Noferini, M.; Sprocatti, M.; Costa, G. Perspectives on the Use of a Seaweed Extract to Moderate the Negative Effects of Alternate Bearing in Apple Trees. J. Hortic. Sci. Biotechnol. 2009, 84, 131–137. [Google Scholar] [CrossRef]
- Malaguti, D.; Rombolà, A.D.; Gerin, M.; Simoni, G.; Tagliavini, M.; Marangoni, B. Effect of Seaweed Extracts-Based Leaf Sprays on the Mineral Status, Yield and Fruit Quality of Apple. Acta Hortic. 2002. [Google Scholar] [CrossRef]
- de Sousa, A.M.; Ayub, R.A.; Viencz, T.; Botelho, R.V. Fruit Set and Yield of Apple Trees Cv. Gala Treated with Seaweed Extract of Ascophyllum nodosum and Thidiazuron. Rev. Bras. Frutic. 2019. [Google Scholar] [CrossRef] [Green Version]
- Soppelsa, S.; Kelderer, M.; Casera, C.; Bassi, M.; Robatscher, P.; Andreotti, C. Use of Biostimulants for Organic Apple Production: Effects on Tree Growth, Yield, and Fruit Quality at Harvest and during Storage. Front. Plant Sci. 2018. [Google Scholar] [CrossRef] [PubMed]
- Jeannin, I.; Lescure, J.C.; Morot-Gaudry, J.F. The Effects of Aqueous Seaweed Sprays on the Growth of Maize. Bot. Mar. 1991. [Google Scholar] [CrossRef]
- Basavaraja, P.K.; Yogendra, N.D.; Zodape, S.T.; Prakash, R.; Ghosh, A. Effect of Seaweed Sap as Foliar Spray on Growth and Yield of Hybrid Maize. J. Plant Nutr. 2018. [Google Scholar] [CrossRef]
- Pal, A.; Dwivedi, S.K.; Maurya, P.K.; Kanwar, P. Effect of Seaweed Saps on Growth, Yield, Nutrient Uptake and Economic Improvement of Maize (Sweet Corn). J. Appl. Nat. Sci. 2015. [Google Scholar] [CrossRef] [Green Version]
- Layek, J.; Das, A.; Ramkrushna, G.I.; Ghosh, A.; Panwar, A.S.; Krishnappa, R.; Ngachan, S.V. Effect of Seaweed Sap on Germination, Growth and Productivity of Maize (Zea mays) in North Eastern Himalayas. Indian J. Agron. 2016, 61, 354–359. [Google Scholar]
- Devi, R.; Singh, S.; Singh, M.K.; Pal, S.K.; Perween, S.; Kumari, J.; Zodape, S.T.; Ghosh, A. Seaweed Sap As Productivity Booster Of Maize. Bioscan 2015, 10, 1303–1305. [Google Scholar]
- Al-Musawi, M.A.H.M. Effect of Foliar Application with Algae Extracts on Fruit Quality of Sour Orange, Citrus aurantium L. J. Environ. Sci. Pollut. Res. 2018. [Google Scholar] [CrossRef]
- Fornes, F.; Sánchez-Perales, M.; Guardiola, J.L. Effect of a Seaweed Extract on the Productivity of “de Nules” Clementine Mandarin and Navelina Orange. Bot. Mar. 2002, 43, 345–354. [Google Scholar] [CrossRef]
- Fornes, F.; Sánchez-Perales, M.; Guardiola, J.L. Effect of A Seaweed Extract on Citrus Fruit Maturation. Acta Hortic. 1995. [Google Scholar] [CrossRef]
- Gomathi, R.; Kohila, S.; Ramachandiran, K. Evaluating the Effect of Seaweed Formulations on the Quality and Yield of Sugarcane. Madras Agric. J. 2017, 104, 1. [Google Scholar]
- da Silveira, L.C.I.; Mattos, P.; Mógor, Á.F.; Daros, E.; de Oliveira Bettini, M.; Norrie, J. Effect of Kelp Extract on Sugarcane Plantlets Biomass Accumulation. Idesia (Arica) 2015. [Google Scholar] [CrossRef] [Green Version]
- Grundy, J.; Stoker, C.; Carré, I.A. Circadian Regulation of Abiotic Stress Tolerance in Plants. Front. Plant Sci. 2015. [Google Scholar] [CrossRef] [PubMed]
- Billard, V.; Etienne, P.; Jannin, L.; Garnica, M.; Cruz, F.; Garcia-Mina, J.M.; Yvin, J.C.; Ourry, A. Two Biostimulants Derived from Algae or Humic Acid Induce Similar Responses in the Mineral Content and Gene Expression of Winter Oilseed Rape (Brassica napus L.). J. Plant Growth Regul. 2014. [Google Scholar] [CrossRef]
- El Boukhari, M.E.M.; Barakate, M.; Bouhia, Y.; Lyamlouli, K. Trends in Seaweed Extract Based Biostimulants: Manufacturing Process and Beneficial Effect on Soil-Plant Systems. Plants 2020, 9, 359. [Google Scholar] [CrossRef] [Green Version]
- Fan, D.; Hodges, D.M.; Critchley, A.T.; Prithiviraj, B. A Commercial Extract of Brown Macroalga (Ascophyllum nodosum ) Affects Yield and the Nutritional Quality of Spinach In Vitro. Commun. Soil Sci. Plant Anal. 2013. [Google Scholar] [CrossRef]
- Jannin, L.; Arkoun, M.; Etienne, P.; Laîné, P.; Goux, D.; Garnica, M.; Fuentes, M.; Francisco, S.S.; Baigorri, R.; Cruz, F.; et al. Brassica Napus Growth Is Promoted by Ascophyllum nodosum (L.) Le Jol. Seaweed Extract: Microarray Analysis and Physiological Characterization of N, C, and S Metabolisms. J. Plant Growth Regul. 2013. [Google Scholar] [CrossRef]
- Nair, P.; Kandasamy, S.; Zhang, J.; Ji, X.; Kirby, C.; Benkel, B.; Hodges, M.D.; Critchley, A.T.; Hiltz, D.; Prithiviraj, B. Transcriptional and Metabolomic Analysis of Ascophyllum nodosum Mediated Freezing Tolerance in Arabidopsis thaliana. BMC Genom. 2012, 13, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamani-Babgohari, M.; Critchley, A.T.; Norrie, J.; Prithiviraj, B. Increased Freezing Stress Tolerance of Nicotiana Tabacum L. Cv. Bright Yellow-2 Cell Cultures with the Medium Addition of Ascophyllum nodosum (L.) Le Jolis Extract. Vitr. Cell. Dev. Biol. Plant 2019. [Google Scholar] [CrossRef]
- Santaniello, A.; Scartazza, A.; Gresta, F.; Loreti, E.; Biasone, A.; Di Tommaso, D.; Piaggesi, A.; Perata, P. Ascophyllum nodosum Seaweed Extract Alleviates Drought Stress in Arabidopsis by Affecting Photosynthetic Performance and Related Gene Expression. Front. Plant Sci. 2017. [Google Scholar] [CrossRef] [Green Version]
- Goñi, O.; Quille, P.; O’Connell, S. Ascophyllum nodosum Extract Biostimulants and Their Role in Enhancing Tolerance to Drought Stress in Tomato Plants. Plant Physiol. Biochem. 2018. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zheng, X.; Song, Y.; Zhu, L.; Yu, Z.; Gan, L.; Zhou, S.; Liu, H.; Wen, F.; Zhu, C. NtLTP4, a Lipid Transfer Protein That Enhances Salt and Drought Stresses Tolerance in Nicotiana tabacum. Sci. Rep. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jithesh, M.N.; Shukla, P.S.; Kant, P.; Joshi, J.; Critchley, A.T.; Prithiviraj, B. Physiological and Transcriptomics Analyses Reveal That Ascophyllum nodosum Extracts Induce Salinity Tolerance in Arabidopsis by Regulating the Expression of Stress Responsive Genes. J. Plant Growth Regul. 2019. [Google Scholar] [CrossRef]
- Elansary, H.O.; Yessoufou, K.; Abdel-Hamid, A.M.E.; El-Esawi, M.A.; Ali, H.; Elshikh, M.S. Seaweed Extracts Enhance Salam Turfgrass Performance during Prolonged Irrigation Intervals and Saline Shock. Front. Plant Sci. 2017. [Google Scholar] [CrossRef] [Green Version]
- Cluzet, S.; Torregrosa, C.; Jacquet, C.; Lafitte, C.; Fournier, J.; Mercier, L.; Salamagne, S.; Briand, X.; Esquerré-Tugayé, M.T.; Dumas, B. Gene Expression Profiling and Protection of Medicago truncatula against a Fungal Infection in Response to an Elicitor from Green Algae Ulva Spp. Plant Cell Environ. 2004. [Google Scholar] [CrossRef]
- Kunkel, B.N.; Brooks, D.M. Cross Talk between Signaling Pathways in Pathogen Defense. Curr. Opin. Plant Biol. 2002, 34, 354–366. [Google Scholar] [CrossRef]
- Mabeau, S.; Kloareg, B. Isolation and Analysis of the Cell Walls of Brown Algae: Fucus Spiralis, F. ceranoides, F. vesiculosus, F. serratus, Bifurcaria bifurcata and Laminaria digitata. J. Exp. Bot. 1987. [Google Scholar] [CrossRef]
- Chevolot, L.; Mulloy, B.; Ratiskol, J.; Foucault, A.; Colliec-Jouault, S. A Disaccharide Repeat Unit Is the Major Structure in Fucoidans from Two Species of Brown Algae. Carbohydr. Res. 2001. [Google Scholar] [CrossRef]
- Klarzynski, O.; Plesse, B.; Joubert, J.M.; Yvin, J.C.; Kopp, M.; Kloareg, B.; Fritig, B. Linear SS-1,3-Glucans Are Elicitors of Defense Responses in Tobacco. Plant Physiol. 2000, 45, 567–588. [Google Scholar] [CrossRef] [Green Version]
- Mercier, L.; Lafitte, C.; Borderies, G.; Briand, X.; Esquerré-Tugayé, M.T.; Fournier, J. The Algal Polysaccharide Carrageenans Can Act as an Elicitor of Plant Defence. New Phytol. 2001, 6, 66–87. [Google Scholar] [CrossRef]
- El Modafar, C.; Elgadda, M.; El Boutachfaiti, R.; Abouraicha, E.; Zehhar, N.; Petit, E.; El Alaoui-Talibi, Z.; Courtois, B.; Courtois, J. Induction of Natural Defence Accompanied by Salicylic Acid-Dependant Systemic Acquired Resistance in Tomato Seedlings in Response to Bioelicitors Isolated from Green Algae. Sci. Hortic. (Amsterdam) 2012, 138, 55–63. [Google Scholar] [CrossRef]
- Sahayaraj, K.; Kalidas, S. Evaluation of Nymphicidal and Ovicidal Effect of a Seaweed, Padina pavonica (Linn.) (Phaeophyceae) on Cotton Pest, Dysdercus cingulatus (Fab.). Indian J. Mar. Sci. 2011, 40, 125–129. [Google Scholar]
- El-Ansary, M.S.M.; Hamouda, R.A. Biocontrol of Root-Knot Nematode Infected Banana Plants by Some Marine Algae. Russ. J. Mar. Biol. 2014. [Google Scholar] [CrossRef]
- Dubiella, U.; Seybold, H.; Durian, G.; Komander, E.; Lassig, R.; Witte, C.-P.; Schulze, W.X.; Romeis, T. Calcium-Dependent Protein Kinase/NADPH Oxidase Activation Circuit Is Required for Rapid Defense Signal Propagation. Proc. Natl. Acad. Sci. USA 2013, 110, 8744–8749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delaunois, B.; Farace, G.; Jeandet, P.; Clément, C.; Baillieul, F.; Dorey, S.; Cordelier, S. Elicitors as Alternative Strategy to Pesticides in Grapevine? Current Knowledge on Their Mode of Action from Controlled Conditions to Vineyard. Environ. Sci. Pollut. Res. 2014, 43, 111–166. [Google Scholar] [CrossRef] [PubMed]
- Nihorimbere, V.; Ongena, M.; Smargiassi, M.; Thonart, P. Beneficial Effect of the Rhizosphere Microbial Community for Plant Growth and Health. Benef. Eff. Rhizosph. Microb. Community Plant Growth Health 2011, 15, 327–337. [Google Scholar]
- Renaut, S.; Masse, J.; Norrie, J.P.; Blal, B.; Hijri, M. A Commercial Seaweed Extract Structured Microbial Communities Associated with Tomato and Pepper Roots and Significantly Increased Crop Yield. Microb. Biotechnol. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Fu, F.; Li, J.; Wang, G.; Wu, M.; Zhan, J.; Chen, X.; Mao, Z. Effects of Seaweed Fertilizer on the Growth of Malus hupehensis Rehd. Seedlings, Soil Enzyme Activities and Fungal Communities under Replant Condition. Eur. J. Soil Biol. 2016. [Google Scholar] [CrossRef]
- Yalçın, S.; Şükran Okudan, E.; Karakaş, Ö.; Önem, A.N.; Sözgen Başkan, K. Identification and Quantification of Some Phytohormones in Seaweeds Using UPLC-MS/MS. J. Liq. Chromatogr. Relat. Technol. 2019. [Google Scholar] [CrossRef]
- Stirk, W.A.; Van Staden, J. Plant Growth Regulators in Seaweeds: Occurrence, Regulation and Functions; Elsevier: Amsterdam, The Netherlands, 2014; Volume 71. [Google Scholar]
- MacKinnon, S.L.; Hiltz, D.; Ugarte, R.; Craft, C.A. Improved Methods of Analysis for Betaines in Ascophyllum nodosum and Its Commercial Seaweed Extracts. J. Appl. Phycol. 2010, 22, 489–494. [Google Scholar] [CrossRef] [Green Version]
- McNeil, S.D.; Nuccio, M.L.; Hanson, A.D. Betaines and Related Osmoprotectants. Targets for Metabolic Engineering of Stress Resistance. Plant Physiol. 1999, 67, 45–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nabil, S.; Cosson, J. Seasonal Variations in Sterol Composition of Delesseria sanguinea (Ceramiales, Rhodophyta). Hydrobiologia 1996. [Google Scholar] [CrossRef]
- Hamdy, A.E.A.; Dawes, C.J. Proximate Constituents and Lipid Chemistry in Two Species of Sargassum from the West Coast of Florida. Bot. Mar. 1988. [Google Scholar] [CrossRef]
- Govindan, M.; Hodge, J.D.; Brown, K.A.; Nuñez-Smith, M. Distribution of Cholesterol in Caribbean Marine Algae. Steroids 1993. [Google Scholar] [CrossRef]
- Kumari, P.; Kumar, M.; Reddy, C.R.K.; Jha, B. Algal Lipids, Fatty Acids and Sterols. In Functional Ingredients from Algae for Foods and Nutraceuticals; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Poojary, M.M.; Barba, F.J.; Aliakbarian, B.; Donsì, F.; Pataro, G.; Dias, D.A.; Juliano, P. Innovative Alternative Technologies to Extract Carotenoids from Microalgae and Seaweeds. Mar. Drugs 2016, 14, 214. [Google Scholar] [CrossRef]
- Othman, R.; Amin, N.A.; Sani, M.S.A.; Fadzillah, N.A.; Jamaludin, M.A. Carotenoid and Chlorophyll Profiles in Five Species of Malaysian Seaweed as Potential Halal Active Pharmaceutical Ingredient (API). Int. J. Adv. Sci. Eng. Inf. Technol. 2018. [Google Scholar] [CrossRef]
- Circuncisão, A.R.; Catarino, M.D.; Cardoso, S.M.; Silva, A.M.S. Minerals from Macroalgae Origin: Health Benefits and Risks for Consumers. Mar. Drugs 2018, 16, 400. [Google Scholar] [CrossRef] [Green Version]
- Seaweeds, O.F. Mineral Nutrients Found in Seaweed and Their Function. Available online: http://www.ocean-fresh-seaweeds.com/seaweed_mineral_nutrients_and_function.html (accessed on 18 September 2020).
- Parthiban, C.; Saranya, C.; Girija, K.; Hemalatha, A.; Suresh, M.; Anantharaman, P. Biochemical Composition of Some Selected Seaweeds from Tuticorin Coast. Pelagia Res. Libr. 2013, 4, 362–366. [Google Scholar]
- Cotas, J.; Leandro, A.; Monteiro, P.; Pacheco, D.; Figueirinha, A.; Goncąlves, A.M.M.; Da Silva, G.J.; Pereira, L. Seaweed Phenolics: From Extraction to Applications. Mar. Drugs 2020, 18, 384. [Google Scholar] [CrossRef]
- Chojnacka, K.; Kim, S.K. Introduction of Marine Algae Extracts. Mar. Algae Extr. Process. Prod. Appl. 2015. [Google Scholar] [CrossRef]
- El-Baroty, G.S.; El-Baz, F.K.; Abd-Elmoein, A.; Abd El Baky, H.H.; Ali, M.M.; Ibrahim, A.E. Evaluation of Glycolipids of Some Egyptian Marine Algae as a Source of Bioactive Substances. Electron. J. Environ. Agric. Food Chem. 2011, 10, 2114–2128. [Google Scholar]
- Guschina, I.A.; Harwood, J.L. Lipids and Lipid Metabolism in Eukaryotic Algae. Prog. Lipid Res. 2006. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, M.; Valentão, P.; Andrade, P.B. Biologically Active Oxylipins from Enzymatic and Nonenzymatic Routes in Macroalgae. Mar. Drugs 2016, 14, 23. [Google Scholar] [CrossRef] [Green Version]
- Pangestuti, R.; Kim, S.K. Seaweed Proteins, Peptides, and Amino Acids. Seaweed Sustain. Food Non-Food Appl. 2015. [Google Scholar] [CrossRef]
- Fleurence, J.; Morançais, M.; Dumay, J. Seaweed Proteins. In Proteins in Food Processing, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Yu, K.X.; Jantan, I.; Ahmad, R.; Wong, C.L. The Major Bioactive Components of Seaweeds and Their Mosquitocidal Potential. Parasitol. Res. 2014. [Google Scholar] [CrossRef] [PubMed]
- Dubey, N.K. Natural Products in Plant Pest Management; CABI: Wallingford, UK, 2010. [Google Scholar]
- Ratnadass, A.; Fernandes, P.; Avelino, J.; Habib, R. Plant Species Diversity for Sustainable Management of Crop Pests and Diseases in Agroecosystems: A Review. Agron. Sustain. Dev. 2012. [Google Scholar] [CrossRef] [Green Version]
- Thundiyil, J.G.; Stober, J.; Besbelli, N.; Pronczuk, J. Acute Pesticide Poisoning: A Proposed Classification Tool. Bull. World Health Organ. 2008. [Google Scholar] [CrossRef]
- Omura, M.; Hirata, M.; Zhao, M.; Tanaka, A.; Inoue, N. Comparative Testicular Toxicities of Two Isomers of Dichloropropanol, 2,3-Dichloro-1-Propanol, and 1,3-Dichloro-2-Propanol, and Their Metabolites Alpha-Chlorohydrin and Epichlorohydrin, and the Potent Testicular Toxicant 1,2-Dibromo-3-Chloropropane. Bull. Environ. Contam. Toxicol. 1995. [Google Scholar] [CrossRef]
- Feng, W.; Zheng, X. Essential Oils to Control Alternaria Alternata in Vitro and in Vivo. Food Control 2007. [Google Scholar] [CrossRef]
- Caradonia, F.; Battaglia, V.; Righi, L.; Pascali, G.; La Torre, A. Plant Biostimulant Regulatory Framework: Prospects in Europe and Current Situation at International Level. J. Plant Growth Regul. 2019. [Google Scholar] [CrossRef]
- Singh, A.K. Integrated Crop, Nutrient and Pest Management for Improving Tomato, Brinjal and Chilli Productivity in Acid Soils. Int. J. Plant Prot. 2017. [Google Scholar] [CrossRef]
- Chinnadurai, C.; Ramkissoon, A.; Rajendran, R.; deAspa, S.; Ramsubhag, A.; Jayaraj, J. Integrated Disease Management in Pumpkin in the Southern Caribbean. Trop. Agric. Univ. West Indies 2018, 95, 132–140. [Google Scholar]
- Zodape, S.T. Seaweeds As a Biofertilizer. J. Sci. Ind. Res. (India) 2001, 60, 378–382. [Google Scholar]
- Tejada-Tejada, P.; Rodriguez-Rodriguez, Y.; Rodriguez, L.E.; Perdomo, O.P.; Boluda, C.J. Quantification of Hevy Metals in Sargassum natans and Sargassum fluitans Arrived to the Coasts of Dominican Republic during 2019. In Proceedings of the Caribbean Science and Innovation Meeting, Pointe-à-Pitre, France, 19–22 October 2019. [Google Scholar]
- Godlewska, K.; Michalak, I.; Tuhy, L.; Chojnacka, K. Plant Growth Biostimulants Based on Different Methods of Seaweed Extraction with Water. Biomed Res. Int. 2016. [Google Scholar] [CrossRef] [Green Version]
- El-Naggar, N.E.A.; Rabei, N.H. Bioprocessing Optimization for Efficient Simultaneous Removal of Methylene Blue and Nickel by Gracilaria Seaweed Biomass. Sci. Rep. 2020. [Google Scholar] [CrossRef] [PubMed]
- Susilorini, R.M.I.R.; Hardjasaputra, H.; Sri, T.; Galih, H.; RPeksa, W.S.; Ginanjar, H.; Joko, S. The Advantage of Natural Polymer Modified Mortar with Seaweed: Green Construction Material Innovation for Sustainable Concrete. Procedia Eng. 2014. [Google Scholar] [CrossRef] [Green Version]
- van Tussenbroek, B.I.; Hernández Arana, H.A.; Rodríguez-Martínez, R.E.; Espinoza-Avalos, J.; Canizales-Flores, H.M.; González-Godoy, C.E.; Barba-Santos, M.G.; Vega-Zepeda, A.; Collado-Vides, L. Severe Impacts of Brown Tides Caused by Sargassum spp. on near-Shore Caribbean Seagrass Communities. Mar. Pollut. Bull. 2017. [Google Scholar] [CrossRef]
| Phaeophyceae | Rhodophyta | Chlorophyta |
|---|---|---|
| Ascophyllum nodosum | Macrocycstis pyrifera | Ulva lactuca |
| Ecklonia maxima | Porphyra perforate | Enteromorpha prolifera |
| Durvillea antarctica | Nereocystis spp. | Caulerpa paspaloides |
| Durvillea protatorum | Cyanidium caldarium | Ulva armoricana |
| Fucus vesiculosus | Gelidium serrulatum | Codium Liyengarii |
| Sargassum spp. | Acanthophora spicifera | Codium tomentosum |
| Hydroclathrus spp. | Kappaphycus alvarezii | Caulerpa sertularioides |
| Ralfsia spp. | Gracilaria edulis | |
| Laminaria digitata | Gracilaria dura | |
| Cystoseira myriophylloides | Laurencia johnstonii | |
| Fucus spiralis | ||
| Padina pavonica | ||
| Fucus gardneri | ||
| Durvillaea antarctica |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, O.; Ramsubhag, A.; Jayaraman, J. Biostimulant Properties of Seaweed Extracts in Plants: Implications towards Sustainable Crop Production. Plants 2021, 10, 531. https://doi.org/10.3390/plants10030531
Ali O, Ramsubhag A, Jayaraman J. Biostimulant Properties of Seaweed Extracts in Plants: Implications towards Sustainable Crop Production. Plants. 2021; 10(3):531. https://doi.org/10.3390/plants10030531
Chicago/Turabian StyleAli, Omar, Adesh Ramsubhag, and Jayaraj Jayaraman. 2021. "Biostimulant Properties of Seaweed Extracts in Plants: Implications towards Sustainable Crop Production" Plants 10, no. 3: 531. https://doi.org/10.3390/plants10030531
APA StyleAli, O., Ramsubhag, A., & Jayaraman, J. (2021). Biostimulant Properties of Seaweed Extracts in Plants: Implications towards Sustainable Crop Production. Plants, 10(3), 531. https://doi.org/10.3390/plants10030531