
Co-Inoculation of Mesorhizobium ciceri with Either Bacillus sp. or Enterobacter aerogenes on Chickpea Improves Growth and Productivity in Phosphate-Deficient Soils in Dry Areas of a Mediterranean Region



Abstract
:1. Introduction
2. Results
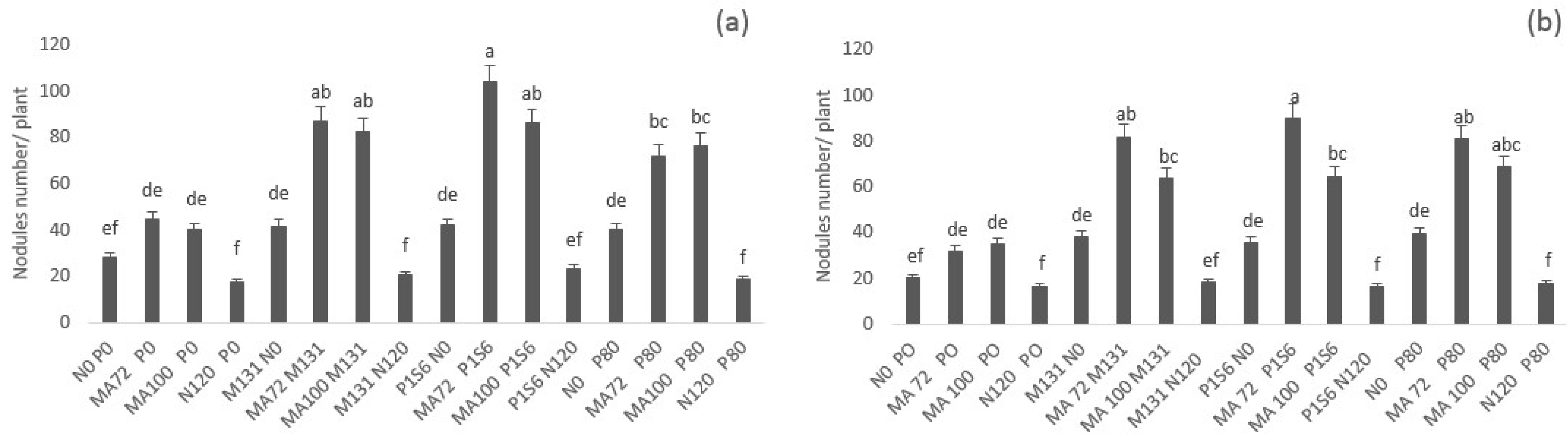
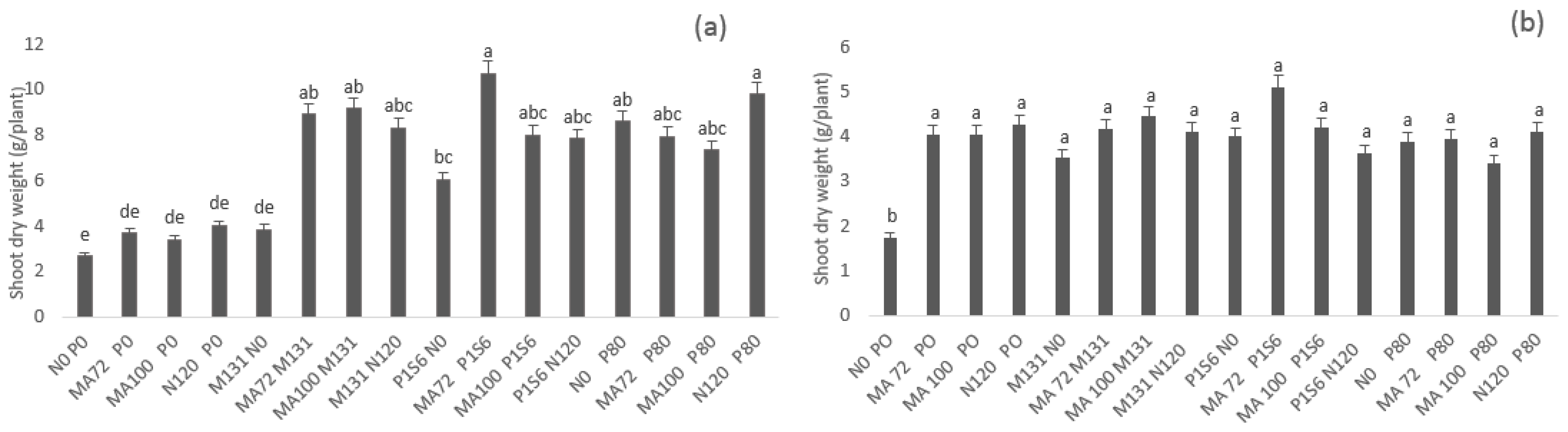
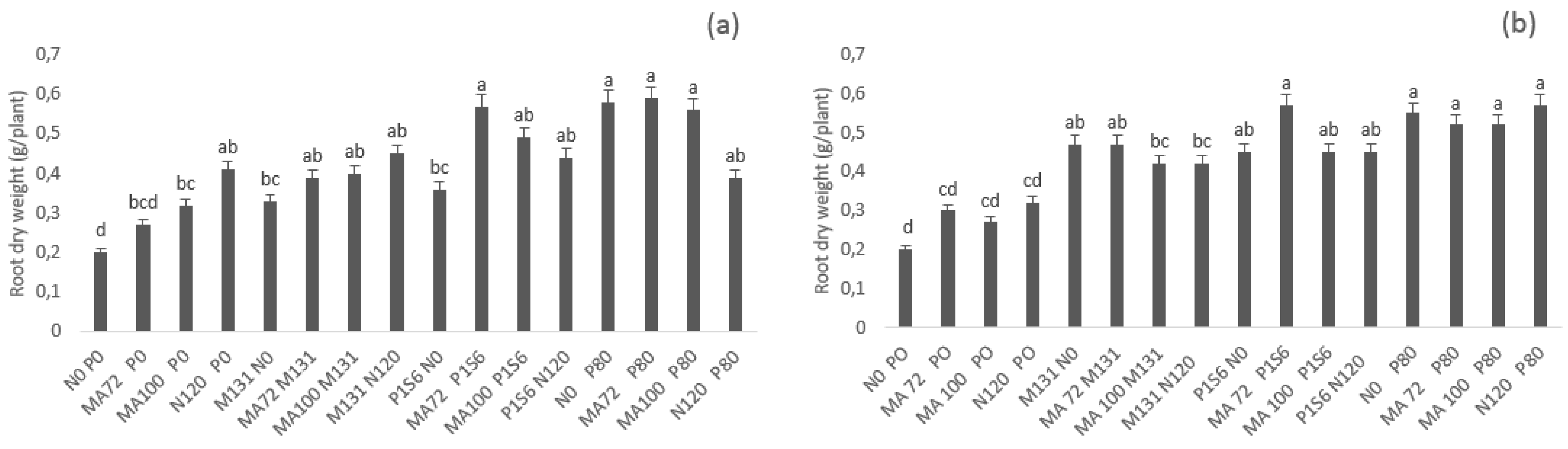
2.1. Effect of Co-Inoculation and Application of N and P Fertilization of Chickpea on Nodulation, Percent of Nitrogen Derived from Air, Shoot, and Root Dry Weight under Low Phosphate Conditions
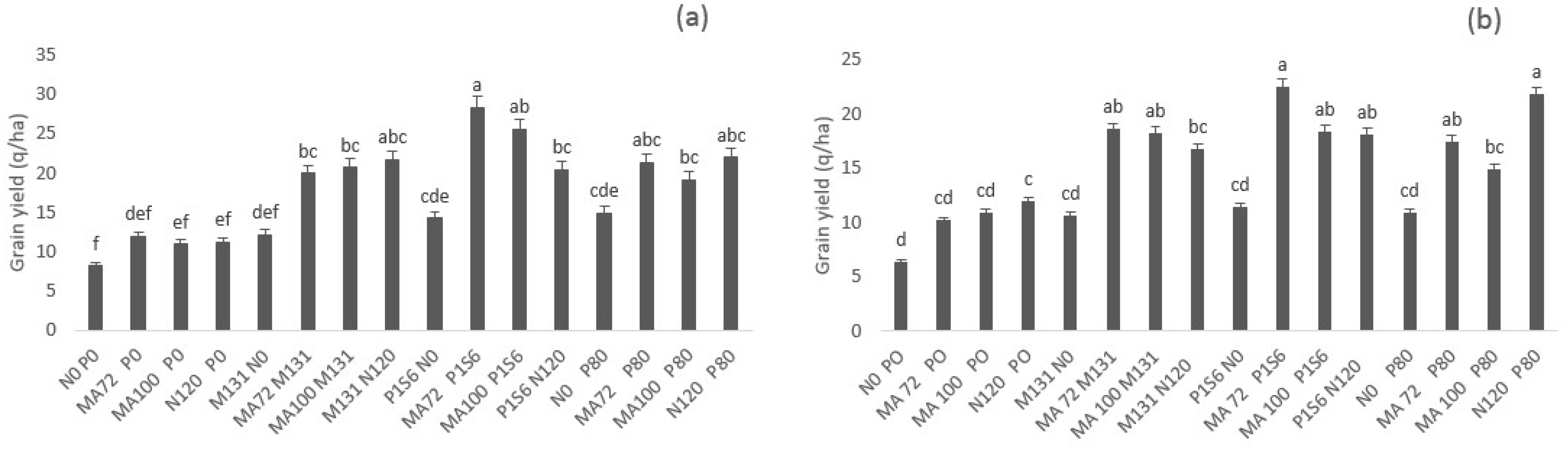
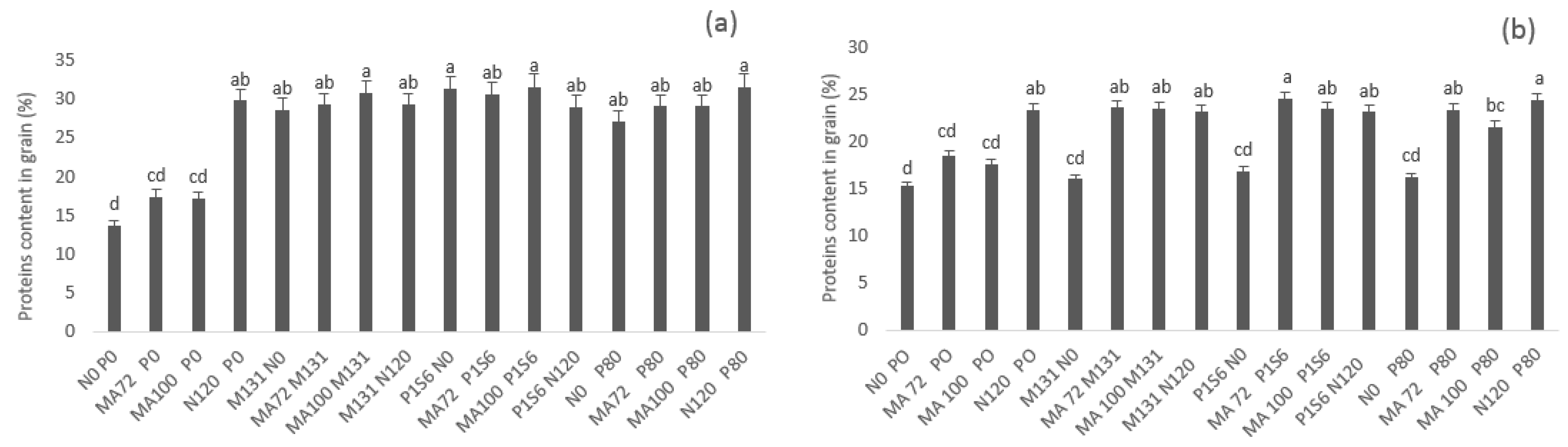
2.2. Effect of Co-Inoculation and Application of N and P Fertilization on Grain Yield, Straw Yield and Grain P, N, and Protein Content under Low Phosphate Conditions
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains Used in This Study
4.2. Assessment of Co-Inoculation of Chickpea with Mesorhizobium spp. and Bacillus sp. or E. aerogenes
4.2.1. Plant Material and Experimental Sites
4.2.2. Inoculum Production
4.2.3. Seeds Inoculation
4.2.4. Field Experiment Design
- N0 P0: Uninoculated and unfertilized treatment (control)
- MA72 P0: Seeds inoculated with MA72
- MA100 P0: Seeds inoculated with MA100
- N120 P0: Application of Nitrogen at a rate of 120 kg N/ ha, no inoculation was applied in this treatment
- M131 N0: Seeds inoculated with M131
- MA72 M131: Seeds inoculated with MA72 + M131
- MA100 M131: Seeds inoculated with MA100 + M131
- M131 N120: Seeds inoculated with M131 + Nitrogen application at a rate of 120 kg N/ ha
- P1S6 N0: Seeds inoculated with P1S6
- MA72 P1S6: Seeds inoculated with MA72 + P1S6
- MA100 P1S6: Seeds inoculated with MA100 + P1S6
- P1S6 N120: Seeds inoculated with P1S6 + Nitrogen application at a rate of 120 kg N/ ha
- N0 P80: Application of phosphate at a rate of 35 kg P/ha, no inoculation was applied in this treatment
- MA72 P80: Seeds inoculated with MA72 + phosphate application at a rate of 35 kg P/ha
- MA100 P80: Seeds inoculated with MA100 + phosphate application at a rate of 35 kg P/ha
- N120 P80: Application of 120 kg N/ ha and of 35 kg P/ha, no inoculation was applied in this treatment
4.3. Data Collection
4.3.1. Nodules Collection and Biomass Measurement
4.3.2. Grain Yield and Total Dry Matter
4.3.3. Nitrogen, Phosphate, and Protein Content in Grain
4.3.4. Nitrogen Derived from the Atmosphere
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. FAO Statistical Databases (Faostat), Food and Agriculture Organization of the United Nations (FAO). 2019. Available online: http://faostat3.fao.org/home/E (accessed on 12 March 2021).
- Khadraji, A.; Mouradi, M.; Ghoulam, C. Growth and mineral nutrition of the chickpea (Cicer arietinum L.)-rhizobia symbiosis under water deficit. Braz. Arch. Biol. Technol. 2017, 60, e17160325. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Al-Whaibi, M.H.; Sakran, A.M.; Basalah, M.O.; Ali, H.M. Effect of calcium and potassium on antioxidant system of Vicia faba L. under cadmium stress. Int. J. Mol. Sci. 2012, 13, 6604–6619. [Google Scholar] [CrossRef] [Green Version]
- Jukanti, A.K.; Gaur, P.M.; Gowda, C.; Chibbar, R.N. Nutritional quality and health benefits of chickpea (Cicer arietinum L.): A review. Brit. J. Nutr. 2012, 108, 11–26. [Google Scholar] [CrossRef] [Green Version]
- Wolde-meskel, E.; van Heerwaarden, J.; Abdulkadir, B.; Kassa, S.; Aliyi, I.; Degefu, T.; Wakweya, K.; Kanampiu, F.; Giller, K.E. Additive yield response of chickpea (Cicer arietinum L.) to rhizobium inoculation and phosphorus fertilizer across smallholder farms in Ethiopia. Agric. Ecosyst. Environ. 2018, 261, 144–152. [Google Scholar] [CrossRef]
- Tena, W.; Wolde-Meskel, E.; Degefu, T.; Walley, F. Genetic and phenotypic diversity of rhizobia nodulating Chickpea (Cicer arietinum L.) in soils from southern and central Ethiopia. Can. J. Microbiol. 2017, 63, 690–707. [Google Scholar] [CrossRef] [Green Version]
- Funga, A.; Ojiewo, C.O.; Turoop, L.; Mwangi, G.S. Symbiotic effectiveness of elite rhizobia strains nodulating desi type chickpea (Cicer arietinum L.) varieties. J. Plant. Sci. 2016, 4, 88–94. [Google Scholar] [CrossRef]
- Mabrouk, Y.; Hemissi, I.; Salem, I.B.; Mejri, S.; Saidi, M.; Belhadj, O. Potential of rhizobia in improving nitrogen fixation and yields of legumes. In Symbiosis; Rigobelo, E., Ed.; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef] [Green Version]
- Benjelloun, I.; Thami-Alami, I.; Douira, A.; Udupa, S.M. Phenotypic and genotypic diversity among symbiotic and non-symbiotic bacteria present in chickpea nodules in Morocco. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Zahran, H.H. Rhizobium-legume symbiosis and nitrogen fixation under severe conditions and in an arid climate. Microbiol. Mol. Biol. Rev. 1999, 63, 968–989. [Google Scholar] [CrossRef] [Green Version]
- Khan, N.; Bano, A.; Zandi, P. Effects of exogenously applied plant growth regulators in combination with PGPR on the physiology and root growth of chickpea (Cicer arietinum) and their role in drought tolerance. J. Plant Interact. 2018, 13, 239–247. [Google Scholar] [CrossRef] [Green Version]
- Getahun, A.; Muleta, D.; Assefa, F.; Kiros, S. Field application of rhizobial inoculants in enhancing faba bean production in acidic soils: An innovative strategy to improve crop productivity. In Salt Stress, Microbes, and Plant Interactions: Causes and Solution; Akhtar, M.S., Ed.; Springer: Singapore, 2019; Volume 1, pp. 147–180. [Google Scholar] [CrossRef]
- Yadav, S.K.; Singh, S.; Singh, H.B.; Sarma, B.K. Compatible rhizosphere-competent microbial consortium adds value to the nutritional quality in edible parts of chickpea. J. Agric. Food Chem. 2017, 65, 6122–6130. [Google Scholar] [CrossRef] [PubMed]
- Bargaz, A.; Lyamlouli, K.; Chtouki, M.; Zeroual, Y.; Dhiba, D. Soil microbial resources for improving fertilizers efficiency in an integrated plant nutrient management system. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, F.; Brígido, C.; Glick, B.; Oliveira, S.; Alho, L. Mesorhizobium ciceri LMS-1 expressing an exogenous 1-aminocyclopropane-1-carboxylate (ACC) deaminase increases its nodulation abilities and chickpea plant resistance to soil constraints. Lett. Appl. Microbiol. 2012, 55, 15–21. [Google Scholar] [CrossRef]
- Howieson, J.; Ballard, R. Optimising the legume symbiosis in stressful and competitive environments within southern Australia—some contemporary thoughts. Soil Biol. Biochem. 2004, 36, 1261–1273. [Google Scholar] [CrossRef]
- Bidyarani, N.; Prasanna, R.; Babu, S.; Hossain, F.; Saxena, A.K. Enhancement of plant growth and yields in Chickpea (Cicer arietinum L.) through novel cyanobacterial and biofilmed inoculants. Microbiol. Res. 2016, 188–189, 97–105. [Google Scholar] [CrossRef]
- Sarma, R.K.; Saikia, R. Alleviation of drought stress in mung bean by strain Pseudomonas aeruginosa GGRJ21. Plant Soil 2014, 377, 111–126. [Google Scholar] [CrossRef]
- Rashid, S.; Charles, T.C.; Glick, B.R. Isolation and characterization of new plant growth-promoting bacterial endophytes. Appl. Soil Ecol. 2012, 61, 217–224. [Google Scholar] [CrossRef]
- Sepúlveda-Caamaño, M.; Gerding, M.; Vargas, M.; Moya-Elizondo, E.; Oyarzúa, P.; Campos, J. Lentil (Lens culinaris L.) growth promoting rhizobacteria and their effect on nodulation in coinoculation with rhizobia. Arch. Agron. Soil Sci. 2018, 64, 244–256. [Google Scholar] [CrossRef]
- Benhizia, Y.; Benhizia, H.; Benguedouar, A.; Muresu, R.; Giacomini, A.; Squartini, A. Gamma proteobacteria can nodulate legumes of the genus Hedysarum. Syst. Appl. Microbiol. 2004, 27, 462–468. [Google Scholar] [CrossRef]
- Zaheer, A.; Mirza, B.S.; McLean, J.E.; Yasmin, S.; Shah, T.M.; Malik, K.A.; Mirza, M.S. Association of plant growth-promoting Serratia spp. with the root nodules of chickpea. Res. Microbiol. 2016, 167, 510–520. [Google Scholar] [CrossRef]
- Laranjo, M.; Alexandre, A.; Oliveira, S. Legume growth-promoting rhizobia: An overview on the Mesorhizobium genus. Microbiol. Res. 2014, 169, 2–17. [Google Scholar] [CrossRef]
- Vorholt, J.A. Microbial life in the phyllosphere. Nat. Rev. Microbiol. 2012, 10, 828–840. [Google Scholar] [CrossRef]
- Duca, D.; Lorv, J.; Patten, C.L.; Rose, D.; Glick, B.R. Indole-3-acetic acid in plant–microbe interactions. Antonie Van Leeuwenhoek 2014, 106, 85–125. [Google Scholar] [CrossRef]
- De Vasconcelos Martins Ferreira, L.; De Carvalho, F.; Fonseca Colombo Andrade, J.; Padua Oliveira, D.; Vasconcelos De Medeiros, F.H.; De Souza Moreira, F.M. Co-inoculation of selected nodule endophytic rhizobacterial strains with Rhizobium tropici promotes plant growth and controls damping off in common bean. Pedosphere 2020, 30, 98–108. [Google Scholar] [CrossRef]
- Ali, S.; Charles, T.; Glick, B. Delay of flower senescence by bacterial endophytes expressing 1-aminocyclopropane-1-carboxylate deaminase. J. Appl. Microbiol. 2012, 113, 1139–1144. [Google Scholar] [CrossRef] [PubMed]
- Ahemad, M.; Kibret, M. Mechanisms and applications of plant growth promoting rhizobacteria: Current perspective. J. King Saud Univ. Sci. 2013, 26, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Ju, W.; Liu, L.; Jin, X.; Duan, C.; Cui, Y.; Wang, J.; Ma, D.; Zhao, W.; Wang, Y.; Fang, L. Co-inoculation effect of plant-growth-promoting rhizobacteria and rhizobium on EDDS assisted phytoremediation of Cu contaminated soils. Chemosphere 2020, 254, 126724. [Google Scholar] [CrossRef]
- Suzaki, T.; Yoro, E.; Kawaguchi, M. Leguminous plants: Inventors of root nodules to accommodate symbiotic bacteria. In International Review of Cell and Molecular Biology; Elsevier: Amsterdam, The Netherlands, 2015; Volume 316, pp. 111–158. [Google Scholar] [CrossRef]
- Mitran, T.; Meena, R.S.; Lal, R.; Layek, J.; Kumar, S.; Datta, R. Role of soil phosphorus on legume production. In Legumes for Soil Health and Sustainable Management; Meena, R., Das, A., Yadav, G., Lal, R., Eds.; Springer: Singapure, 2018; pp. 487–510. [Google Scholar] [CrossRef]
- Akram, M.; Hussain, S.; Hamid, A.; Majeed, S.; Chaudary, S.; Shah, Z.; Yaqoob, A.; Kayani, F.; Arif, U.; Fareed, K. Interactive effect of phosphorus and potassium on growth, yield, quality and seed production of chili (Capsicum annuum L.). J. Hortic. 2017, 4. [Google Scholar] [CrossRef] [Green Version]
- Niu, Y.F.; Chai, R.S.; Jin, G.L.; Wang, H.; Tang, C.X.; Zhang, Y.S. Responses of root architecture development to low phosphorus availability: A review. Ann. Bot. 2012, 112, 391–408. [Google Scholar] [CrossRef]
- Schulze, J.; Temple, G.; Temple, S.J.; Beschow, H.; Vance, C.P. Nitrogen fixation by white lupin under phosphorus deficiency. Ann. Bot. 2006, 98, 731–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Liao, H.; Lucas, W.J. Molecular mechanisms underlying phosphate sensing, signaling, and adaptation in plants. J. Integr. Plant Biol. 2014, 56, 192–220. [Google Scholar] [CrossRef]
- Vance, C.P.; Uhde-Stone, C.; Allan, D.L. Phosphorus acquisition and use: Critical adaptations by plants for securing a nonrenewable resource. New Phytol. 2003, 157, 423–447. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Arredondo, D.L.; Leyva-González, M.A.; González-Morales, S.I.; López-Bucio, J.; Herrera-Estrella, L. Phosphate nutrition: Improving low-phosphate tolerance in crops. Annu. Rev. Plant Biol. 2014, 65, 95–123. [Google Scholar] [CrossRef]
- Sulieman, S.; Tran, L.-S.P. Phosphorus homeostasis in legume nodules as an adaptive strategy to phosphorus deficiency. Plant Sci. 2015, 239, 36–43. [Google Scholar] [CrossRef]
- Rotaru, V.; Sinclair, T.R. Interactive influence of phosphorus and iron on nitrogen fixation by soybean. Environ. Exp. Bot. 2009, 66, 94–99. [Google Scholar] [CrossRef]
- Cabeza, R.A.; Liese, R.; Lingner, A.; von Stieglitz, I.; Neumann, J.; Salinas-Riester, G.; Pommerenke, C.; Dittert, K.; Schulze, J. RNA-seq transcriptome profiling reveals that Medicago truncatula nodules acclimate N2 fixation before emerging P deficiency reaches the nodules. J. Exp. Bot. 2014, 65, 6035–6048. [Google Scholar] [CrossRef] [Green Version]
- Meena, R.S.; Meena, P.D.; Yadav, G.S.; Yadav, S.S. Phosphate solubilizing microorganisms, Principles and application of microphos technology. J. Clean. Prod. 2017, 145. [Google Scholar] [CrossRef]
- Lazali, M.; Blavet, D.; Pernot, C.; Desclaux, D.; Drevon, J.J. Efficiency of phosphorus use for dinitrogen fixation varies between common bean genotypes under phosphorus limitation. Agron. J. 2017, 109, 283–290. [Google Scholar] [CrossRef]
- Tilman, D.; Fargione, J.; Wolff, B.; D’Antonio, C.; Dobson, A.; Howarth, R.; Schindler, D.; Schlesinger, W.H.; Simberloff, D.; Swackhamer, D. Forecasting agriculturally driven global environmental change. Science 2001, 292, 281–284. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Hu, B.; Qi, K.; Chen, W.; Pang, X.; Bao, W.; Tian, G. Effects of phosphorus addition on soil microbial biomass and community composition in a subalpine spruce plantation. Eur. J. Soil Biol. 2016, 72, 35–41. [Google Scholar] [CrossRef]
- Khan, M.S.; Zaidi, A.; Wani, P.A. Role of phosphate solubilizing microorganisms in sustainable agriculture—A review. In Sustainable Agriculture; Lichtfouse, E., Navarrete, M., Debaeke, P., Véronique, S., Alberola, C., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 551–570. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharyya, P.N.; Jha, D.K. Plant growth-promoting rhizobacteria (PGPR): Emergence in agriculture. World J. Microbiol. Biotechnol. 2012, 28, 1327–1350. [Google Scholar] [CrossRef]
- Khan, M.S.; Zaidi, A.; Ahmad, E. Mechanism of phosphate solubilization and physiological functions of phosphate-solubilizing microorganisms. In Phosphate Solubilizing Microorganisms; Khan, M.S., Zaidi, A., Musarrat, J., Eds.; Springer: Cham, Switzerland, 2014; pp. 31–62. [Google Scholar] [CrossRef]
- Korir, H.; Mungai, N.; Thuita, M.; Hamba, Y.; Masso, C. Co-inoculation effect of rhizobia and plant growth promoting rhizobacteria on common bean growth in a low phosphorus soil. Front. Plant Sci. 2017, 8, 141. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, A.C.; Gutiérrez, R.T.; Santana, R.C.; Urrutia, A.R.; Fauvart, M.; Michiels, J.; Vanderleyden, J. Effects of co-inoculation of native Rhizobium and Pseudomonas strains on growth parameters and yield of two contrasting Phaseolus vulgaris L. genotypes under Cuban soil conditions. Eur. J. Soil Biol. 2014, 62, 105–112. [Google Scholar] [CrossRef]
- Raklami, A.; Bechtaoui, N.; Tahiri, A.-i.; Anli, M.; Meddich, A.; Oufdou, K. Use of rhizobacteria and mycorrhizae consortium in the open field as a strategy for improving crop nutrition, productivity and soil fertility. Front. Microbiol. 2019, 10, 1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumsane, T.M.; Cheng-Hua, H.; Yuh-Ming, H.; Ming-Yi, Y. Effects of coinoculation of Rhizobium with plant growth promoting rhizobacteria on the nitrogen fixation and nutrient uptake of Trifolium repens in low phosphorus soil. J. Plant Nutr. 2020, 43, 739–752. [Google Scholar] [CrossRef]
- Morel, M.; Braña, V.; Castro-Sowinski, S. Legume crops, importance and uses of bacterial inoculation to increase production. In Crop Plant; Goyal, A., Ed.; InTech: London, UK, 2012; pp. 217–240. [Google Scholar] [CrossRef] [Green Version]
- Fox, S.; O’Hara, G.; Bräu, L. Enhanced nodulation and symbiotic effectiveness of Medicago truncatula when co-inoculated with Pseudomonas fluorescens WSM3457 and Ensifer (Sinorhizobium) medicae WSM419. Plant Soil 2011, 348, 245–254. [Google Scholar] [CrossRef] [Green Version]
- Fatnassi, I.C.; Chiboub, M.; Saadani, O.; Jebara, M.; Jebara, S.H. Phytostabilization of moderate copper contaminated soils using co-inoculation of Vicia faba with plant growth promoting bacteria. J. Basic Microbiol. 2015, 55, 303–311. [Google Scholar] [CrossRef]
- Kong, Z.; Deng, Z.; Glick, B.R.; Wei, G.; Chou, M. A nodule endophytic plant growth-promoting Pseudomonas and its effects on growth, nodulation and metal uptake in Medicago lupulina under copper stress. Ann. Microbiol. 2017, 67, 49–58. [Google Scholar] [CrossRef]
- Sibponkrung, S.; Kondo, T.; Tanaka, K.; Tittabutr, P.; Boonkerd, N.; Yoshida, K.-i.; Teaumroong, N. Co-inoculation of Bacillus velezensis strain S141 and Bradyrhizobium Strains promotes nodule growth and nitrogen fixation. Microorganisms 2020, 8, 678. [Google Scholar] [CrossRef]
- Singh, O.; Gupta, M.; Mittal, V.; Kiran, S.; Nayyar, H.; Gulati, A.; Tewari, R. Novel phosphate solubilizing bacteria ‘Pantoea cypripedii PS1′ along with Enterobacter aerogenes PS16 and Rhizobium ciceri enhance the growth of chickpea (Cicer arietinum L.). Plant Growth Regul. 2013, 73. [Google Scholar] [CrossRef]
- Van Noorden, G.E.; Verbeek, R.; Dinh, Q.D.; Jin, J.; Green, A.; Ng, J.L.P.; Mathesius, U. Molecular signals controlling the inhibition of nodulation by nitrate in Medicago truncatula. Int. J. Mol. Sci. 2016, 17, 1060. [Google Scholar] [CrossRef] [Green Version]
- Zeijl, A.v.; Guhl, K.; Xiao, T.T.; Shen, D.; Geurts, R.; Kohlen, W. Nitrate inhibition of nodule formation in Medicago truncatula is mediated by ACC Synthase10. bioRxiv 2018, 434829. [Google Scholar] [CrossRef] [Green Version]
- Streeter, J.G. Nitrate inhibition of legume nodule growth and activity: I. Long term studies with a continuous supply of nitrate. Plant Physiol. 1985, 77, 321–324. [Google Scholar] [CrossRef] [Green Version]
- Shin, C.-S.; Sharif, M.O.; Lee, H.-Y. Evaluating the Effect of Bacterial Inoculation and Fertilization on the Soil Nutrient Status of Coal Mine Soil by Growing Soybean (Glycine max) and Shrub Lespedeza (Lespedeza bicolor). Sustainability 2018, 10, 4793. [Google Scholar] [CrossRef] [Green Version]
- Jacoby, R.; Peukert, M.; Succurro, A.; Koprivova, A.; Kopriva, S. The Role of Soil Microorganisms in Plant Mineral Nutrition-Current Knowledge and Future Directions. Front. Plant Sci. 2017, 8, 1617. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.S.; Zaidi, A.; Wani, P.A. Role of phosphate-solubilizing microorganisms in sustainable agriculture—A review. Agron. Sustain. Dev. 2007, 27, 29–43. [Google Scholar] [CrossRef]
- Ben Romdhane, S.; Aouani, M.E.; Mhamdi, R. Inefficient nodulation of chickpea (Cicer arietinum L.) in the arid and Saharan climates in Tunisia by Sinorhizobium meliloti biovar medicaginis. Ann. Microbiol. 2007, 57, 15–19. [Google Scholar] [CrossRef]
- Remans, R.; Ramaekers, L.; Schelkens, S.; Hernandez, G.; Garcia, A.; Reyes, J.; Mendez, N.; Toscano, V.; Mulling, M.; Galvez, L.; et al. Effect of Rhizobium–Azospirillum coinoculation on nitrogen fixation and yield of two contrasting Phaseolus vulgaris L. genotypes cultivated across different environments in Cuba. Plant Soil 2008, 312, 25–37. [Google Scholar] [CrossRef]
- Berg, G. Plant–microbe interactions promoting plant growth and health: Perspectives for controlled use of microorganisms in agriculture. Appl. Microbiol. Biotechnol. 2009, 84, 11–18. [Google Scholar] [CrossRef]
- Bedoussac, L.; Journet, E.-P.; Hauggaard-Nielsen, H.; Naudin, C.; Corre-Hellou, G.; Jensen, E.S.; Prieur, L.; Justes, E. Ecological principles underlying the increase of productivity achieved by cereal-grain legume intercrops in organic farming: A review. Agron. Sustain. Dev. 2015, 35, 911–935. [Google Scholar] [CrossRef]
- Jebara, M.; Aouani, M.E.; Payre, H.; Drevon, J.-J. Nodule conductance varied among common bean (Phaseolus vulgaris) genotypes under phosphorus deficiency. J. Plant Physiol. 2005, 162, 309–315. [Google Scholar] [CrossRef]
- Sumit, K.; Chakrabarti, B.; Prasanna, R.; Pratap, D.; Singh, S.D.; Purakayastha, T.J.; Pathak, H. Elevated carbon dioxide level along with phosphorus application and cyanobacterial inoculation enhances nitrogen fixation and uptake in cowpea crop. Arch. Agron. Soil Sci. 2017, 63, 1927–1937. [Google Scholar] [CrossRef]
- Kyei-Boahen, S.; Savala, C.E.; Chikoye, D.; Abaidoo, R. Growth and yield responses of cowpea to inoculation and phosphorus fertilization in different environments. Front. Plant Sci. 2017, 8, 646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudresh, D.; Shivaprakash, M.; Prasad, R. Effect of combined application of Rhizobium, phosphate solubilizing bacterium and Trichoderma spp. on growth, nutrient uptake and yield of chickpea (Cicer aritenium L.). Appl. Soil Ecol. 2005, 28, 139–146. [Google Scholar] [CrossRef]
- Elkoca, E.; Kantar, F.; Sahin, F. Influence of nitrogen fixing and phosphorus solubilizing bacteria on the nodulation, plant growth, and yield of chickpea. J. Plant Nutr. 2008, 31, 157–171. [Google Scholar] [CrossRef]
- Zafar, M.; Abbasi, M.; Rahim, N.; Khaliq, A.; Shaheen, A.; Jamil, M.; Shahid, M. Influence of integrated phosphorus supply and plant growth promoting rhizobacteria on growth, nodulation, yield and nutrient uptake in Phaseolus vulgaris. Afr. J. Biotechnol. 2011, 10, 16781–16792. [Google Scholar] [CrossRef]
- Burdman, S.; Kigel, J.; Okon, Y. Effects of Azospirillum brasilense on nodulation and growth of common bean (Phaseolus vulgaris L.). Soil Biol. Biochem. 1997, 29, 923–929. [Google Scholar] [CrossRef]
- Cakmakçi, R.; Dönmez, F.; Aydın, A.; Şahin, F. Growth promotion of plants by plant growth-promoting rhizobacteria under greenhouse and two different field soil conditions. Soil Biol. Biochem. 2006, 38, 1482–1487. [Google Scholar] [CrossRef]
- Israr, D.; Mustafa, G.; Khan, K.S.; Shahzad, M.; Ahmad, N.; Masood, S. Interactive effects of phosphorus and Pseudomonas putida on chickpea (Cicer arietinum L.) growth, nutrient uptake, antioxidant enzymes and organic acids exudation. Plant Physiol. Biochem. 2016, 108, 304–312. [Google Scholar] [CrossRef]
- Valverde, A.; Burgos, A.; Fiscella, T.; Rivas, R.; Velázquez, E.; Rodríguez-Barrueco, C.; Cervantes, E.; Chamber, M.; Igual, J.-M. Differential effects of coinoculations with Pseudomonas jessenii PS06 (a phosphate-solubilizing bacterium) and Mesorhizobium ciceri C-2/2 strains on the growth and seed yield of chickpea under greenhouse and field conditions. Plant Soil 2006, 287, 43–50. [Google Scholar] [CrossRef]
- Wani, P.A.; Khan, M.S. Bacillus species enhance growth parameters of chickpea (Cicer arietinum L.) in chromium stressed soils. Food Chem. Toxicol. 2010, 48, 3262–3267. [Google Scholar] [CrossRef]
- Verma, J.; Yadav, J.; Tiwari, K. Application of Rhizobium sp. BHURC01 and plant growth promoting rhizobactria on nodulation, plant biomass and yields of chickpea (Cicer arietinum L.). Int. J. Agric. Res. 2010, 5, 148–156. [Google Scholar] [CrossRef] [Green Version]
- Vurukonda, S.S.K.P.; Vardharajula, S.; Shrivastava, M.; SkZ, A. Enhancement of drought stress tolerance in crops by plant growth promoting rhizobacteria. Microbiol. Res. 2016, 184, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Tilak, K.; Ranganayaki, N.; Manoharachari, C. Synergistic effects of plant-growth promoting rhizobacteria and Rhizobium on nodulation and nitrogen fixation by pigeonpea (Cajanus cajan). Eur. J. Soil Sci. 2006, 57, 67–71. [Google Scholar] [CrossRef]
- Mishra, P.K.; Mishra, S.; Selvakumar, G.; Bisht, J.; Kundu, S.; Gupta, H.S. Coinoculation of Bacillus thuringeinsis-KR1 with Rhizobium leguminosarum enhances plant growth and nodulation of pea (Pisum sativum L.) and lentil (Lens culinaris L.). World J. Microbiol. Biotechnol. 2009, 25, 753–761. [Google Scholar] [CrossRef]
- Kumar, B.D.; Berggren, I.; Mårtensson, A. Potential for improving pea production by co-inoculation with fluorescent Pseudomonas and Rhizobium. Plant Soil 2001, 229, 25–34. [Google Scholar] [CrossRef]
- Singh, N.; Singh, G.; Aggarwal, N.; Khanna, V. Yield enhancement and phosphorus economy in lentil (Lens culinaris Medikus) with integrated use of phosphorus, Rhizobium and plant growth promoting rhizobacteria. J. Plant Nutr. 2018, 41, 737–748. [Google Scholar] [CrossRef]
- Vincent, J.M. A Manual for the Practical Study of the Root-Nodule Bacteria; Oxford, International Biological Programme; Blackwell Scientific Publications: London, UK, 1970; Volume 15. [Google Scholar]
- Dobbelaere, S.; Croonenborghs, A.; Thys, A.; Ptacek, D.; Vanderleyden, J.; Dutto, P.; Labandera-Gonzalez, C.; Caballero-Mellado, J.; Aguirre Medina, J.; Kapulnik, Y.; et al. Responses of agronomically important crops to inoculation with Azospirillum. Funct. Plant Biol. 2001, 28, 871–879. [Google Scholar] [CrossRef]
- El Oumlouki, K.; Moussadek, R.; Zouahri, A.; Dakak, H.; Chati, M.; El Amrani, M. Study of physic-chemical quality of water and soil in the region Souss Massa (Case perimeter Issen), Morocco. J. Mater. Environ. Sci. 2014, 5, 2365–2374. [Google Scholar]
- Jaetzold, R.; Schmidt, H.; Hornetz, B.; Shisanya, C. Farm Management Handbook of Kenya, 2nd ed.; Ministry of Agriculture of Kenya, in Cooperation with the German Agency for Technical Cooperation (GTZ): Nairobi, Kenya, 2005; Volume 2, p. 317. [Google Scholar]
- Kyei-Boahen, S.; Slinkard, A.E.; Walley, F.L. Evaluation of rhizobial inoculation methods for chickpea. Agron. J. 2002, 94, 851–859. [Google Scholar] [CrossRef]
- Jackson, M.L. Soil Chemical Analysis: Advanced Course; UW-Madison Libraries Parallel Press: Madison, WI, USA, 2005. [Google Scholar]
- Nater, E.A. Soil chemical analysis: Advanced course. Soil Sci. 2006, 171, 432–433. [Google Scholar] [CrossRef]
- Herridge, D.F.; Danso, S.K.A. Enhancing crop legume N2 fixation through selection and breeding. In Management of Biological Nitrogen Fixation for the Development of More Productive and Sustainable Agricultural Systems; Ladha, J.K., Peoples, M.B., Eds.; Springer Science + Business Media: Dordrecht, The Netherlands, 1995; Volume 65, pp. 51–82. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | % Ndfa in Ain Sbit | % Ndfa in Merchouch |
|---|---|---|
| MA72 | 22.26 ± 8.1 bc | 16 ± 4.1 b |
| MA72 M131 | 52.18 ± 3.2 a | 48 ± 7.6 a |
| MA72 P1S6 | 57.6 ± 2.8 a | 50.94 ± 5.3 a |
| MA72 P80 | 51.9 ± 6.3 a | 48.5 ± 3.7 a |
| MA100 | 16.78 ± 5.6 c | 16.12 ± 1.3 b |
| MA100 M131 | 52.21 ± 2.7 a | 51.13 ± 2.1 a |
| MA100 P1S6 | 51.41 ± 5.1 a | 52.38 ± 6.6 a |
| MA100 P80 | 58.39 ± 2.7 a | 52.68 ± 6.4 a |
| Site | Mean Rainfall (mm) a | Mean Temperature (°C) a | Climate | Soil Properties | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| pH (HCl) | pH (H2O) | P (ppm) b | K (ppm) b | Total N (%) | Organic Matter (%) | EC (ds/m) c | ||||
| Merchouch | 449 | 17.1 | Semi-arid | 5.7 | 5.0 | 6.72 | 189.27 | 0.4 | 2.5 | 1.6 |
| Ain Sbit | 464 | 17.3 | Semi-arid | 7.9 | 6.9 | 4.49 | 364 | 0.5 | 2.9 | 2.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benjelloun, I.; Thami Alami, I.; El Khadir, M.; Douira, A.; Udupa, S.M. Co-Inoculation of Mesorhizobium ciceri with Either Bacillus sp. or Enterobacter aerogenes on Chickpea Improves Growth and Productivity in Phosphate-Deficient Soils in Dry Areas of a Mediterranean Region. Plants 2021, 10, 571. https://doi.org/10.3390/plants10030571
Benjelloun I, Thami Alami I, El Khadir M, Douira A, Udupa SM. Co-Inoculation of Mesorhizobium ciceri with Either Bacillus sp. or Enterobacter aerogenes on Chickpea Improves Growth and Productivity in Phosphate-Deficient Soils in Dry Areas of a Mediterranean Region. Plants. 2021; 10(3):571. https://doi.org/10.3390/plants10030571
Chicago/Turabian StyleBenjelloun, Imane, Imane Thami Alami, Mohamed El Khadir, Allal Douira, and Sripada M. Udupa. 2021. "Co-Inoculation of Mesorhizobium ciceri with Either Bacillus sp. or Enterobacter aerogenes on Chickpea Improves Growth and Productivity in Phosphate-Deficient Soils in Dry Areas of a Mediterranean Region" Plants 10, no. 3: 571. https://doi.org/10.3390/plants10030571
APA StyleBenjelloun, I., Thami Alami, I., El Khadir, M., Douira, A., & Udupa, S. M. (2021). Co-Inoculation of Mesorhizobium ciceri with Either Bacillus sp. or Enterobacter aerogenes on Chickpea Improves Growth and Productivity in Phosphate-Deficient Soils in Dry Areas of a Mediterranean Region. Plants, 10(3), 571. https://doi.org/10.3390/plants10030571