
Transgenic Expression of dsRNA Targeting the Pentalonia nigronervosa acetylcholinesterase Gene in Banana and Plantain Reduces Aphid Populations

 , , , ,
, , , ,
Abstract
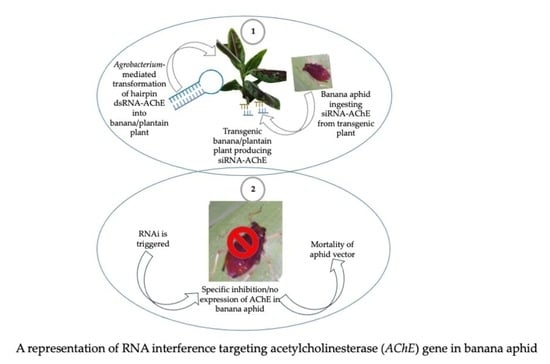
:
1. Introduction
2. Results and Discussion
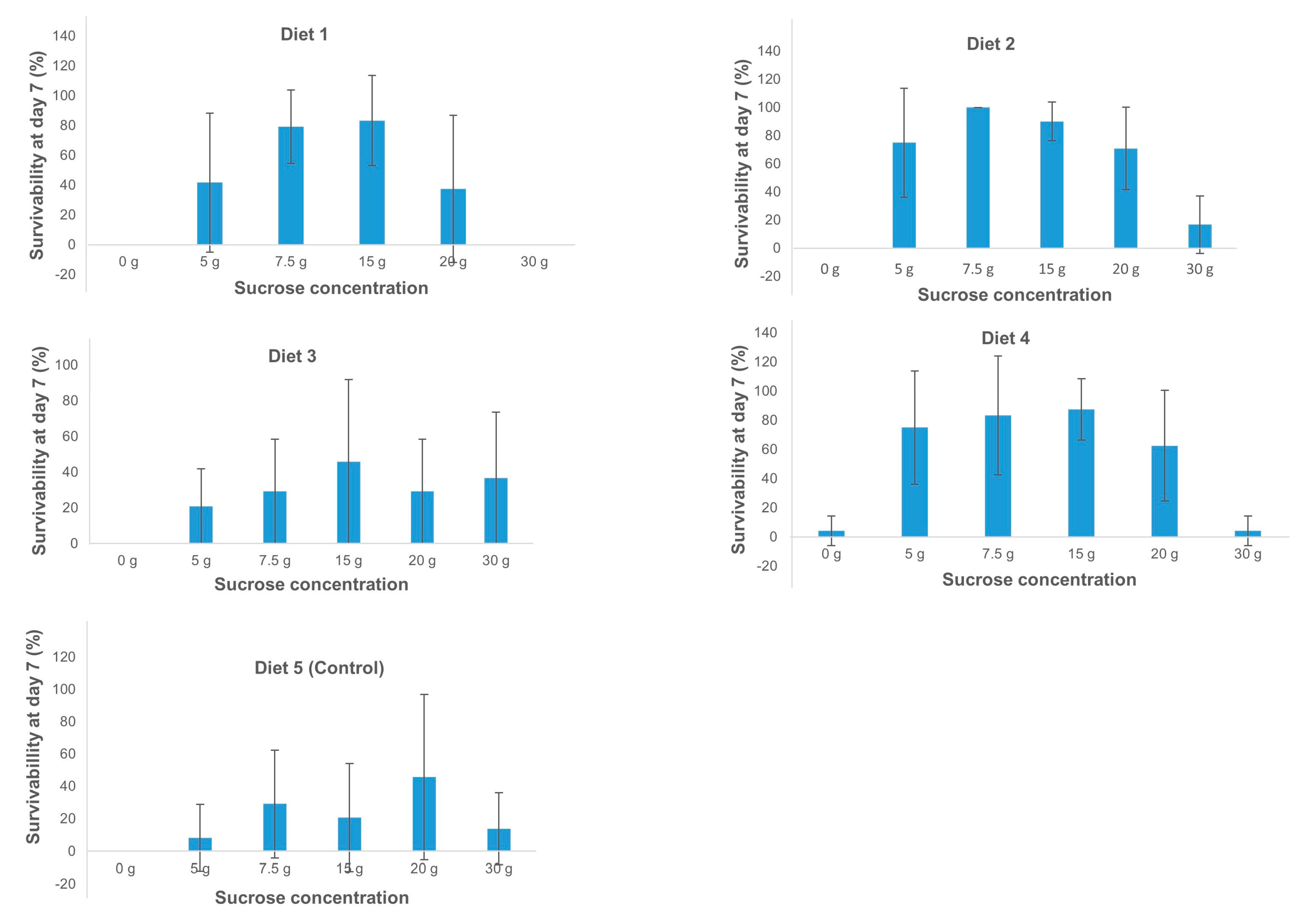
2.1. Optimization of an In Vitro Feeding Diet for Banana Aphids
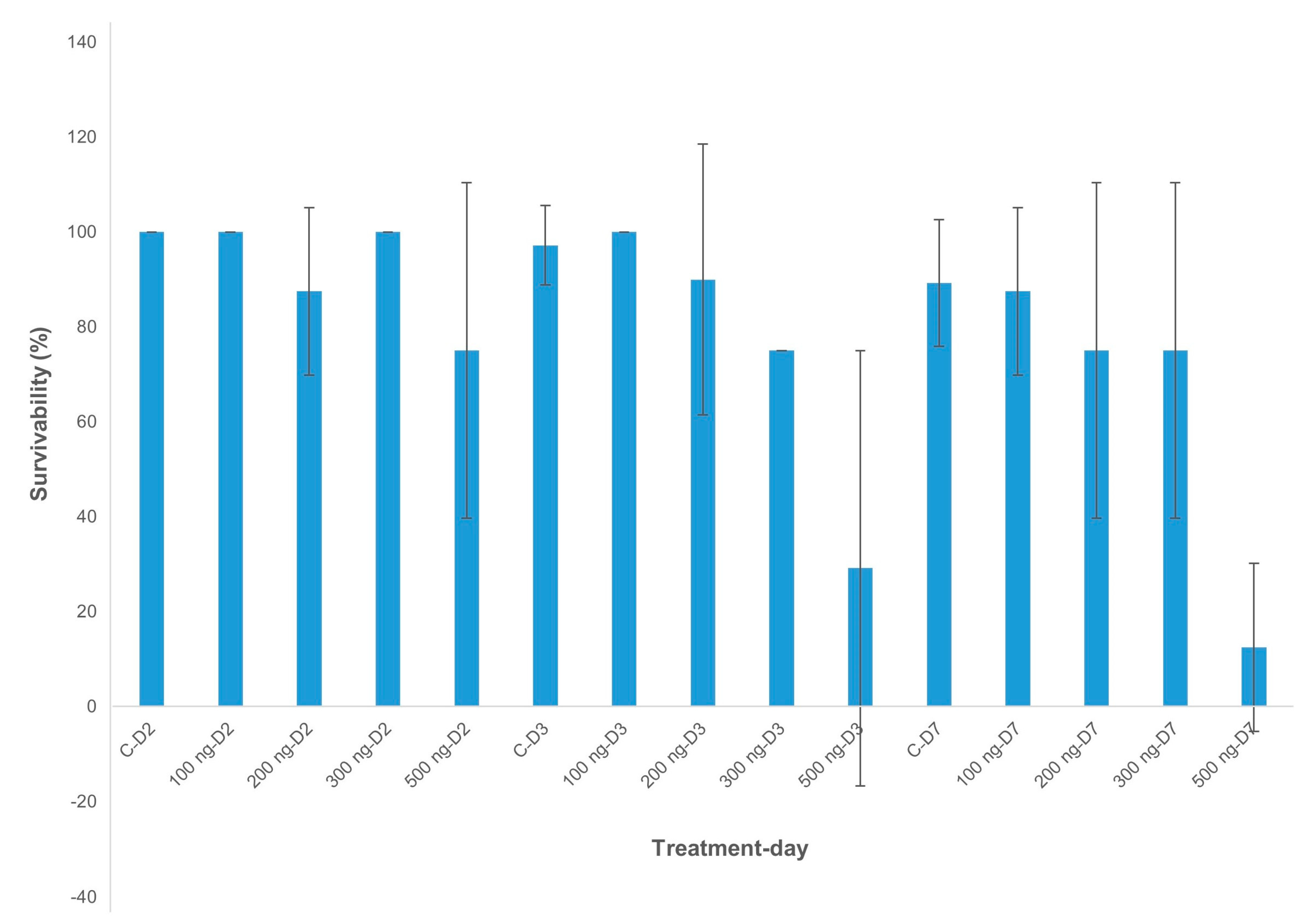
2.2. Dose Effects of dsRNA-AChE on Survival of Banana Aphids
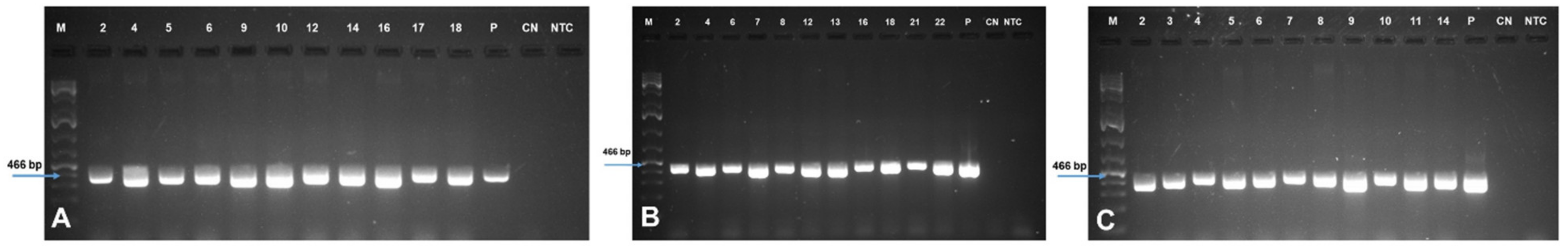
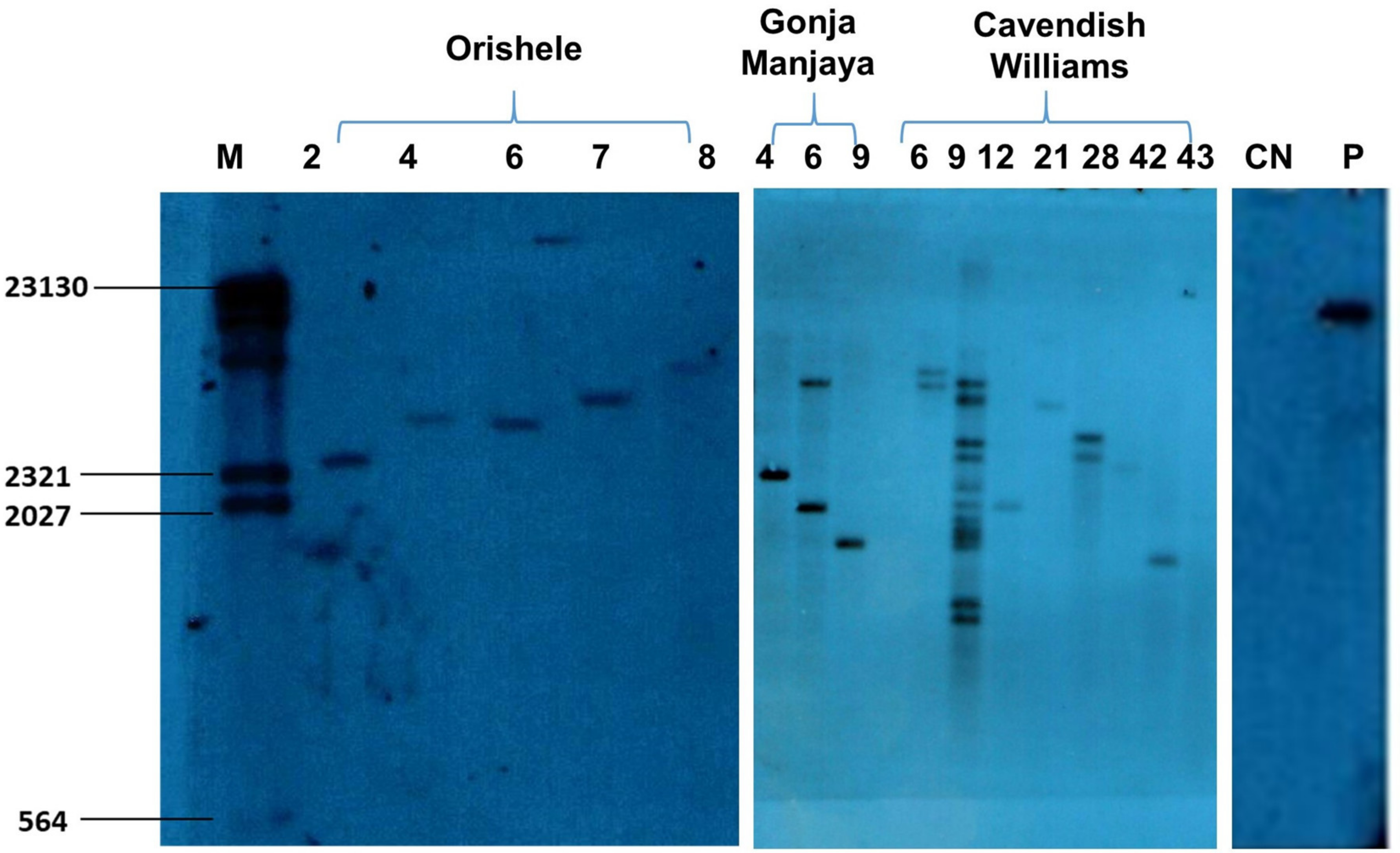
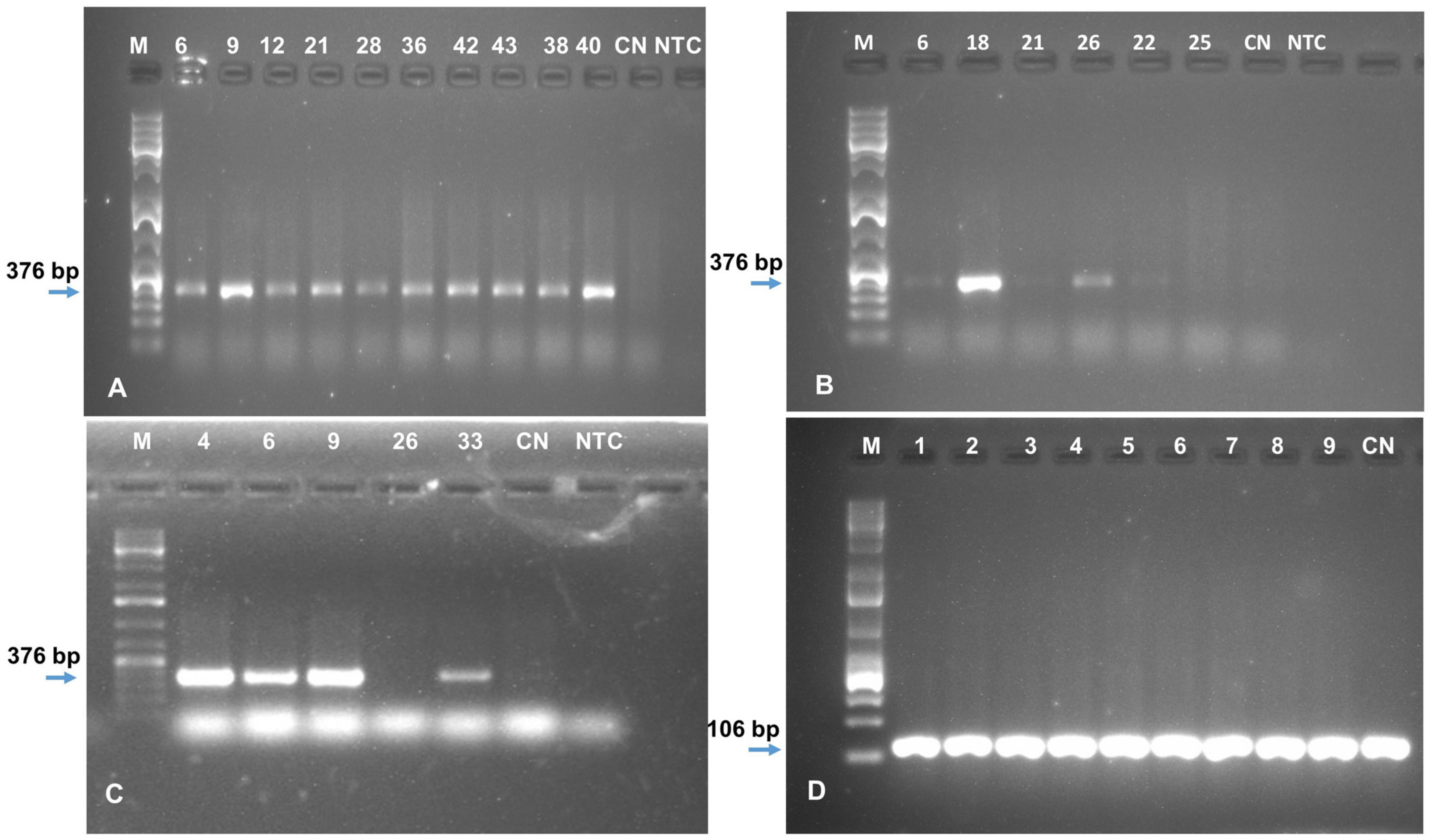
2.3. Generation and Molecular Characterization of Transgenic Events
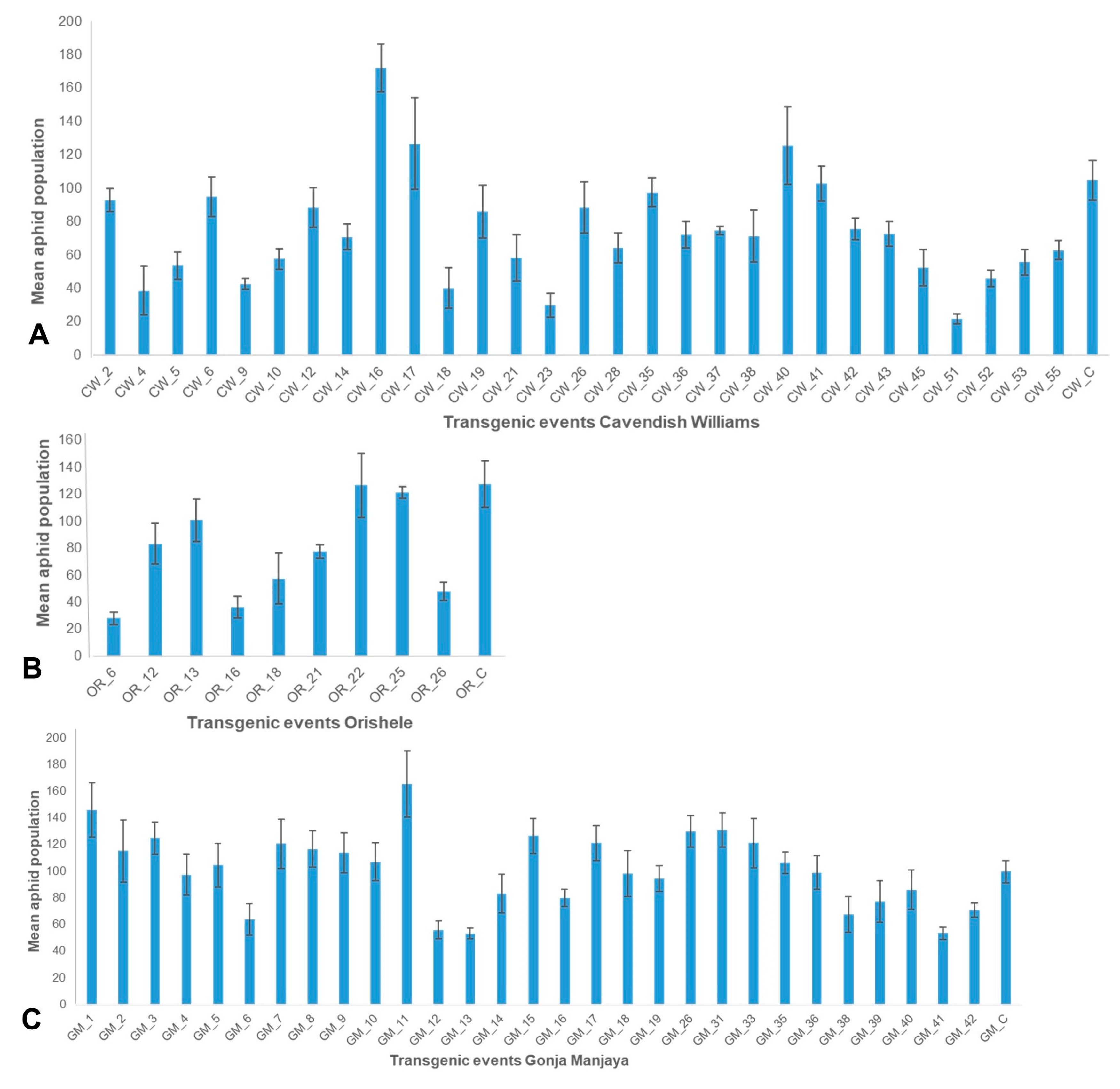
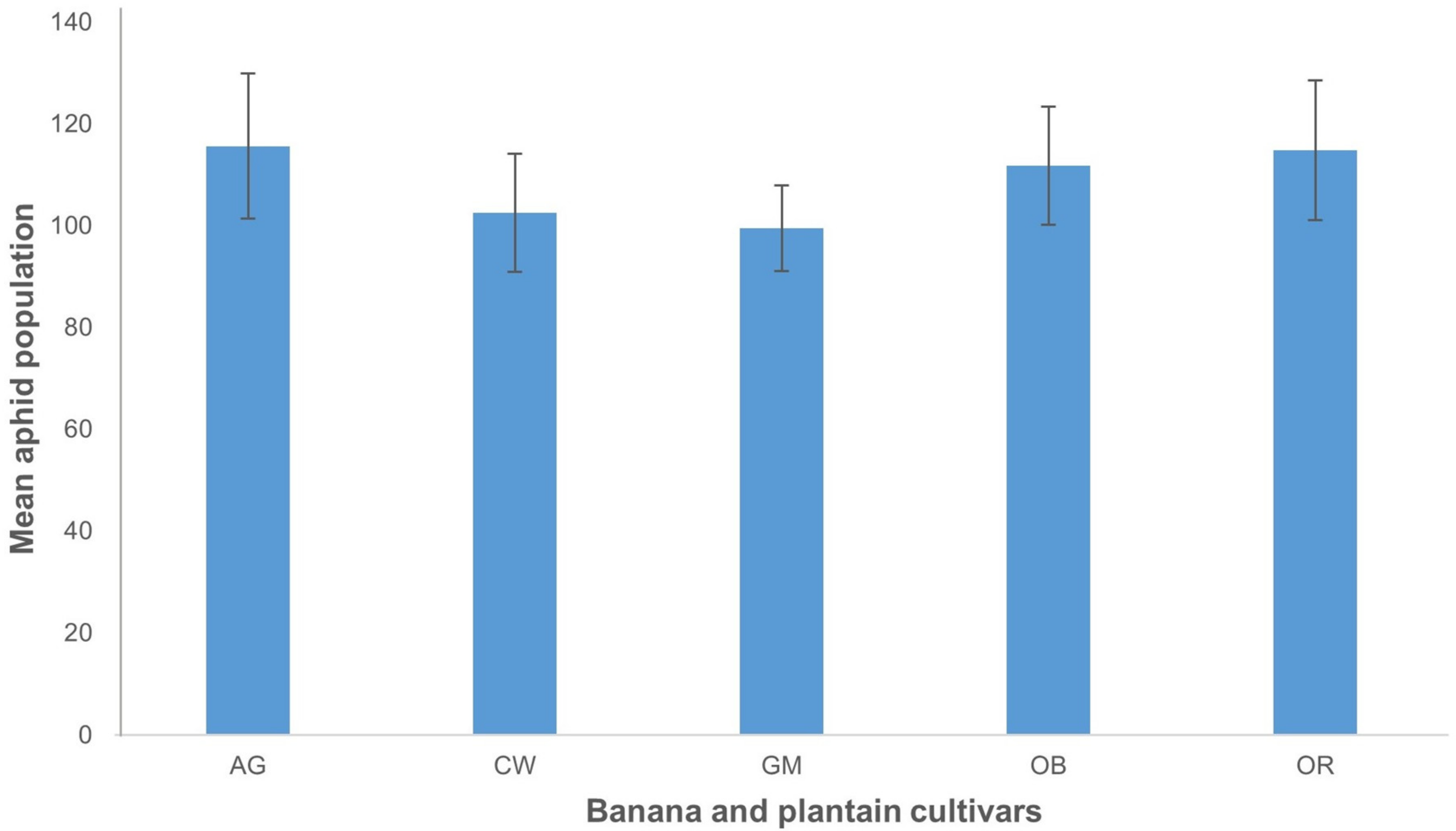
2.4. Evaluation of Transgenic Events for Resistance to Aphids
3. Materials and Methods
3.1. Source of Plant Material
3.2. Rearing and Maintenance of Aphids in the Laboratory
3.3. Optimization of an Artificial Aphid Feeding Medium
3.4. Construction of Plasmids for In Vitro dsRNA Synthesis and Plant Transformation
3.4.1. Amplification and Sequencing of a P. nigronervosa Partial AChE Gene Fragment
3.4.2. Cloning of P. nigronervosa Ache Gene Target Sequence
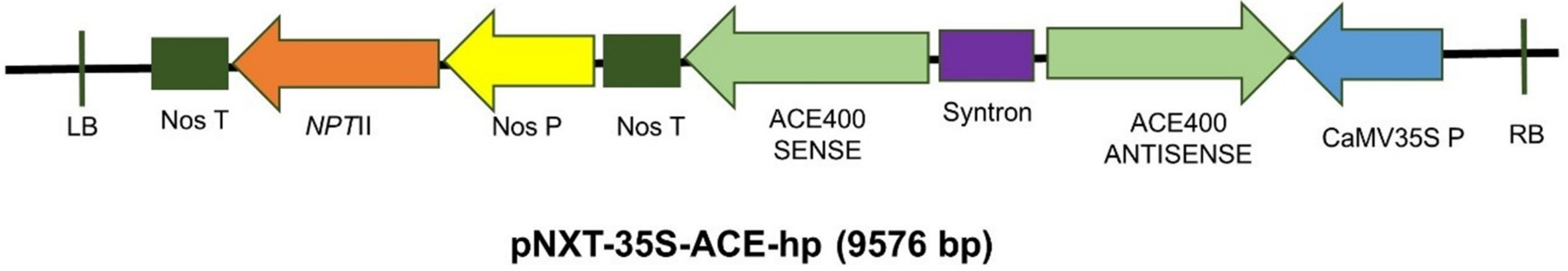
3.4.3. Assembly of a Binary Vector Capable of Expressing P. nigronervosa AChE dsRNA
3.5. In Vitro Synthesis of dsRNA-AChE
3.6. Development of an Artificial Aphid Feeding Assay and dsRNA-AChE Dose Response
3.7. Agrobacterium-Mediated Transformation of Plantain and Banana and Generation of Transgenic Events
3.8. Molecular Characterization of Transgenic Plants
3.8.1. Genomic DNA Extraction and Validation of Transgenic Events by PCR Analysis
3.8.2. Southern Blot Analysis to Confirm the Integration of Transgene
3.8.3. RT-PCR Analysis to Confirm Expression of AChE-dsRNA in Transgenic Plants
3.8.4. Evaluation of Transgenic Events for Resistance to Aphids
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’ s Worst Invasive Alien Species; Invasive Species Specialist Group (ISSG): Auckland, New Zealand, 2000. [Google Scholar]
- Dale, J.L. Banana bunchy top: An economically important tropical plant virus disease. Adv. Virus Res. 1987, 33, 301–325. [Google Scholar] [CrossRef]
- Jekayinoluwa, T.; Tripathi, L.; Tripathi, J.N.; Ntui, V.O.; Obiero, G.; Muge, E.; Dale, J. RNAi Technology for Management of Banana Bunchy Top Disease (BBTD). Food Energy Secur. 2020. [Google Scholar] [CrossRef] [PubMed]
- Selvarajan, R.; Balasubramanian, V. Host-virus interactions in banana-infecting viruses. In Plant Virus—Ost Interaction: Molecular Approaches and Viral Evolution; Gaur, R., Hohn, T., Sharma, P., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 57–72. [Google Scholar]
- Jekayinoluwa, T.; Tripathi, J.N.; Obiero, G.; Muge, E.; Dale, J.; Tripathi, L. Developing Plantain for Resistance to Banana Aphids by RNA Interference. Proceedings 2020, 36, 54. [Google Scholar] [CrossRef] [Green Version]
- Shekhawat, U.K.S.; Ganapathi, T.R.; Hadapad, A.B. Transgenic banana plants expressing small interfering RNAs targeted against viral replication initiation gene display high-level resistance to banana bunchy top virus infection. J. Gen. Virol. 2012, 93, 1804–1813. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, J.N.; Ntui, V.O.; Ron, M.; Muiruri, S.K.; Britt, A.; Tripathi, L. CRISPR/Cas9 editing of endogenous banana streak virus in the B genome of Musaspp. overcomes a major challenge in banana breeding. Commun. Biol. 2019, 2, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dale, J.; James, A.; Paul, J.-Y.; Khanna, H.; Smith, M.; Peraza-Echeverria, S.; Garcia-Bastidas, F.; Kema, G.; Waterhouse, P.; Mengersen, K.; et al. Transgenic Cavendish bananas with resistance to Fusarium wilt tropical race 4. Nat. Commum. 2017, 8, 1496. [Google Scholar] [CrossRef] [Green Version]
- Paul, J.-Y.; Khanna, H.; Kleidon, J.; Hoang, P.; Geijskes, J.; Daniells, J.; Zaplin, E.; Rosenberg, Y.; James, A.; Mlalazi, B.; et al. Golden bananas in the field: Elevated fruit pro-vitamin A from expresion of a single banana transgene. Plant Biotechnol. J. 2017, 15, 520–532. [Google Scholar] [CrossRef] [Green Version]
- Baum, J.A.; Bogaert, T.; Clinton, W.; Heck, G.R.; Feldmann, P.; Ilagan, O.; Johnson, S.; Plaetinck, G.; Munyikwa, T.; Pleau, M.; et al. Control of coleopteran insect pests through RNA interference. Nat. Biotechnol. 2007, 25, 1322–1326. [Google Scholar] [CrossRef]
- Zha, W.; Peng, X.; Chen, R.; Du, B.; Zhu, L.; He, G. Knockdown of midgut genes by dsRNA-transgenic plant-mediated RNA interference in the Hemipteran insect Nilaparvata lugens. PLoS ONE 2011, 6, e20504. [Google Scholar] [CrossRef] [Green Version]
- Laudani, F.; Strano, C.P.; Edwards, M.G.; Malacrinò, A.; Campolo, O.; Abd El Halim, H.M.; Gatehouse, A.M.R.; Palmeri, V. RNAi-mediated gene silencing in Rhynchophorus ferrugineus (Oliver) (Coleoptera: Curculionidae). Open Life Sci. 2017, 12, 214–222. [Google Scholar] [CrossRef]
- Poreddy, S.; Li, J.; Baldwin, I.T. Plant-mediated RNAi silences midgut-expressed genes in congeneric lepidopteran insects in nature. Bmc Plant Biol. 2017, 17, 1–10. [Google Scholar] [CrossRef]
- Knorr, E.; Fishilevich, E.; Tenbusch, L.; Frey, M.L.F.; Rangasamy, M.; Billion, A.; Worden, S.E.; Gandra, P.; Arora, K.; Lo, W.; et al. Gene silencing in Tribolium castaneum as a tool for the targeted identification of candidate RNAi targets in crop pests. Sci. Rep. 2018, 8, 1–15. [Google Scholar] [CrossRef]
- Yoon, J.S.; Mogilicherla, K.; Gurusamy, D.; Chen, X.; Chereddy, S.C.R.R.; Palli, S.R. Double-stranded RNA binding protein, Staufen, is required for the initiation of RNAi in coleopteran insects. Proc. Natl. Acad. Sci. 2018, 115, 8334–8339. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Song, X.; Wang, G.; Yang, K.; Wang, Y.; Niu, L.; Chen, X.; Fang, R. Plant-generated artificial small RNAs mediated aphid resistance. PLoS ONE 2014, 9, e97410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whyard, S.; Singh, A.D.; Wong, S. Ingested double-stranded RNAs can act as species-specific insecticides. Insect Biochem. Mol. Biol. 2009, 39, 824–832. [Google Scholar] [CrossRef]
- Soreq, H.; Seidman, S. Acetylcholinesterase-new roles for an old actor. Perspective. Nat. Rev. 2001, 2, 294. [Google Scholar] [CrossRef] [PubMed]
- Fremaux, I.; Mazeres, S.; Brisson-Lougarre, A.; Arnaud, M.; Ladurantie, C.; Fournier, D. Improvement of Drosophila acetylcholinesterase stability by elimination of a free cysteine. BMC Biochem. 2002. Available online: http://www.biomedcentral.com/1471-2091/3/21 (accessed on 8 January 2021).
- Pang, Y.P.; Singh, S.K.; Gao, Y.; Lassiter, T.L.; Mishra, R.K.; Zhu, K.Y.; Brimijoin, S. Selective and Irreversible Inhibitors of Aphid Acetylcholinesterases: Steps Toward Human-Safe Insecticides. PLoS ONE 2009, 4, e4349. [Google Scholar] [CrossRef] [Green Version]
- Dou, D.; Park, J.G.; Rana, S.; Madden, B.J.; Jiang, H.; Pang, Y.-P. Novel selective and irreversible mosquito aetylcholinesterase inhibitors for controlling malaria and other mosquito-borne diseases. Sci. Rep. 2013, 3, 1068. [Google Scholar] [CrossRef] [Green Version]
- Sabri, A.; Vandermoten, S.; Leroy, P.D.; Haubruge, E.; Hance, T.; Thonart, P.; Francis, F. Proteomic Investigation of Aphid Honeydew Reveals an Unexpected Diversity of Proteins. PLoS ONE 2013, 8, e74656. [Google Scholar] [CrossRef] [Green Version]
- Akey, D.H.; Beck, S.D. Nutrition of the pea aphid, Acyrthosiphon pisum: Requirements for trace metals, sulphur, and cholesterol. J. Insect Physiol. 1972, 18, 1901–1914. [Google Scholar] [CrossRef]
- Dadd, R.H.; Mittler, T.E. Permanent culture of an aphid on a totally synthetic diet. Experientia 1966, 22, 832–833. [Google Scholar] [CrossRef]
- Srivastava, P.N.; Auclair, J.L. Influence of Sucrose Concentration on Diet Uptake and Performance by the Pea Aphid, Acyrthosiphon pisum. Ann. Entomol. Soc. Am. 1971, 64, 739–743. [Google Scholar] [CrossRef]
- Dvir, H.; Silman, I.; Harel, M.; Rosenberry, T.L.; Sussman, J.L. Acetylcholinesterase: From 3D structure to function. Chem. Biol. Interact. 2010, 187, 10–22. [Google Scholar] [CrossRef] [Green Version]
- Colovic, M.B.; Krstic, D.Z.; Lazarevic-Pasti, T.D.; Bondzic, A.M.; Vasic, V.M. Acetylcholinesterase Inhibitors: Pharmacology and Toxicology. Curr. Neuropharmaccol. 2013, 11, 315–335. [Google Scholar] [CrossRef] [Green Version]
- Kishk, A.; Hijaz, F.; Anber, H.A.I.; AbdEl-Raof, T.K.; El-Sherbeni, A.E.H.D.; Hamed, S.; Killiny, N. RNA interference of acetylcholinesterase in the Asian citrus psyllid, Diaphorina citri, increases its susceptibility to carbamate and organophosphate insecticides. Pestic. Biochem. Physiol. 2017, 143, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Wuriyanghan, H.; Rosa, C.; Falk, B.W. Oral delivery of double-stranded RNAs and siRNAs induces RNAi effects in the potato/tomato psyllid, Bactericerca cockerelli. PLoS ONE 2011, 6, e27736. [Google Scholar] [CrossRef] [Green Version]
- Majidiani, S.; PourAbad, R.F.; Laudani, F.; Campolo, O.; Zappala, L.; Rahmani, S.; Mohammadi, S.A.; Palmeri, V. RNAi in Tuta absoluta management: Effects of injection and root delivery of dsRNAs. J. Pest Sci. 2019, 92, 1409–1419. [Google Scholar] [CrossRef]
- Zhang, J.; Khan, S.A.; Hasse, C.; Ruf, S.; Heckel, D.G.; Bock, R. Full crop protection from an insect pest by expression of long double-stranded RNAs in plastids. Science 2015, 347, 991–994. [Google Scholar] [CrossRef]
- Christiaens, O.; Smagghe, G. The challenge of RNAi-mediated control of hemipterans. Insect Sci. 2014, 6, 15–21. [Google Scholar] [CrossRef]
- Ghodke, A.B.; Good, R.T.; Golz, J.F.; Russell, D.A.; Edwards, O.; Robin, C. Exracellular endonucleases in the midgut of Myzus persicae may limit the efficacy of orally delivered RNAi. Sci. Rep. 2019. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Wang, K.; Fu, W.; Sheng, C.; Han, Z. Biochemial comparison of dsRNA degradation nucleases in four different insects. Front. Physiol. 2018, 9, 624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, S.; Qian, J.; Cai, C.; Ma, Z.; Li, J.; Yin, M.; Ren, B.; Shen, J. Spary method application of transdermal dsRNA delivery system for efficient gene silencing and pest control on soybean aphid Aphis Glycines. J. Pest Sci. 2020, 93, 449–459. [Google Scholar] [CrossRef]
- Douglas, A.E.; van Emden, H.F. Nutrition and Symbiosis. In Aphid as Crop Pests, CABI, 2nd ed.; Van Emden, H.F., Harrington, R., Eds.; CABI: Wallingford, UK, 2007; pp. 115–130. [Google Scholar] [CrossRef]
- Stewart, C.N.; Via, L.E. A rapid CTAB DNA isolation technique useful for RAPD fingerprinting and other PCR applications. Biotechniques 1993, 14, 751–752, 754. [Google Scholar]
- Dugdale, B.; Mortimer, C.L.; Kato, M.; James, T.A.; Harding, R.M.; Dale, J.L. In plant activation: An inducible, hyperexpression platform for recombinant protein production in plants. Plant Cell 2013, 25, 2429–2443. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
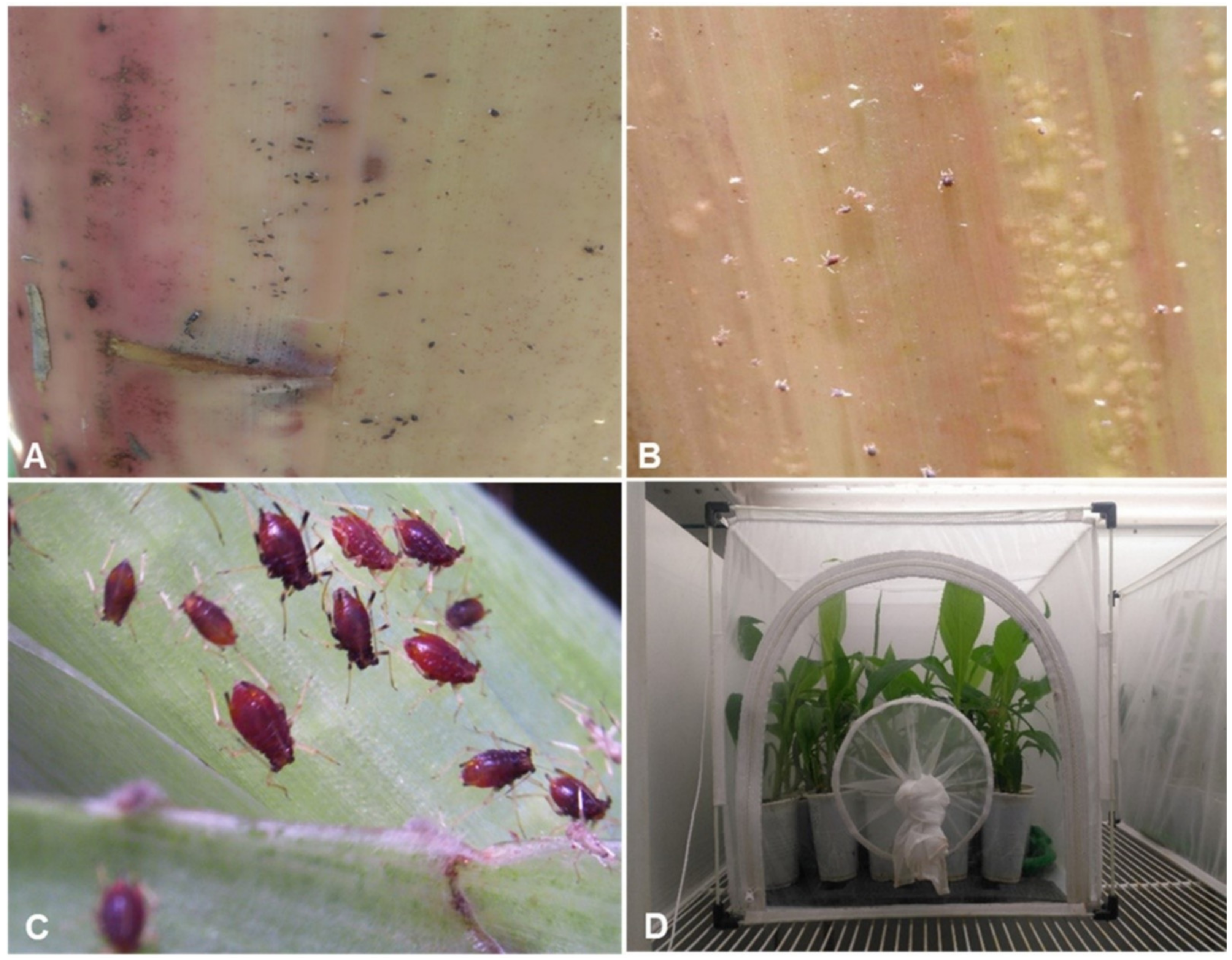{kind=link}
| Day | Sucrose Level (g/100 mL) | Diet Type | NDA | NLA | NN |
|---|---|---|---|---|---|
| 5 | 0 | Diet 1 | 4.0 ± 0 ns | 0 ± 0 ns | 0.08 ± 0.3 ns |
| Diet 2 | 4.0 ± 0 ns | 0 ± 0 ns | 0.05 ± 0.2 ns | ||
| Diet 3 | 4.0 ± 0 ns | 0 ± 0 ns | 0.13 ± 0.4 ns | ||
| Diet 4 | 4.0 ± 0 ns | 0.08 ± 0.3 ns | 0.33 ± 0.7 ns | ||
| Control | 4.0 ± 0 | 0 ± 0 | 0.08 ± 0.3 | ||
| 5 | Diet 1 | 2.0 ± 1.0 ** | 1.75 ± 1.6 ** | 3.5 ± 1.7 *** | |
| Diet 2 | 1.0 ± 1.1 *** | 3.2 ± 1.2 *** | 5.73 ± 2.7 *** | ||
| Diet 3 | 1.0 ± 1 *** | 3.13 ± 1.1 *** | 3.73 ± 1.0 *** | ||
| Diet 4 | 1.0 ± 1 *** | 3.25 ± 1.1 *** | 3.66 ± 1.8 *** | ||
| Control | 4.0 ± 1.0 | 0.16 ± 0.6 | 0.25 ± 0.5 | ||
| 7.5 | Diet 1 | 1.5 ± 1.5 *** | 2.5 ± 1.5 *** | 3.08 ± 1.8 ** | |
| Diet 2 | 0.17 ± 0.5 *** | 3.83 ± 0.5 *** | 8.44 ± 1.8 *** | ||
| Diet 3 | 0.71 ± 0.9 *** | 3.28 ± 0.9 *** | 4.21 ± 0.9 *** | ||
| Diet 4 | 1.25 ± 1.4 *** | 2.75 ± 1.4 *** | 3.66 ± 1.9 *** | ||
| Control | 3.33 ± 1.1 | 0.66 ± 1.1 | 0.83 ± 1.3 | ||
| 15 | Diet 1 | 1.5 ± 1.7 * | 2.5 ± 1.7 * | 3.08 ± 1.9 ** | |
| Diet 2 | 0.78 ± 0.7 *** | 3.21 ± 0.7 *** | 5.35 ± 2.2 *** | ||
| Diet 3 | 1.2 ± 1.5 ** | 2.8 ± 1.5 ** | 3.26 ± 1.6 *** | ||
| Diet 4 | 0.66 ± 0.8 *** | 3.33 ± 0.8 *** | 3.0 ± 1.6 ** | ||
| Control | 2.8 ± 1.3 | 1.16 ± 1.3 | 0.75 ± 1.1 | ||
| 20 | Diet 1 | 2.3 ± 1.9 ns | 1.67 ± 1.9 ns | 2.16 ± 1.9 ns | |
| Diet 2 | 1.4 ± 1.2 ns | 2.6 ± 1.2 ns | 3.0 ± 1.8 *** | ||
| Diet 3 | 1.33 ± 1.3 ns | 2.66 ± 1.3 ns | 2.53 ± 1.5 ** | ||
| Diet 4 | 1.16 ± 1.3 * | 2.83 ± 1.3 * | 2.58 ± 0.7 ** | ||
| Control | 2.66 ± 1.6 | 1.33 ± 1.6 | 0.75 ± 0.9 | ||
| 30 | Diet 1 | 3.08 ± 1.7 ns | 0.83 ± 1.6 ns | 1.08 ± 1.3 ns | |
| Diet 2 | 3.13 ± 1.12 ns | 0.86 ± 1.1 ns | 0.46 ± 0.8 ns | ||
| Diet 3 | 3.00 ± 1.5 ns | 0.86 ± 1.2 ns | 0.86 ± 1.3 ns | ||
| Diet 4 | 3.66 ± 0.5 ns | 0.33 ± 0.5 ns | 0.42 ± 0.7 ns | ||
| Control | 3.58 ± 0.8 | 0.42 ± 0.8 | 0.25 ± 0.5 | ||
| 7 | 0 | Diet 1 | 4.0 ± 0 ns | 0 ± 0 ns | 0 ± 0 ns |
| Diet 2 | 4.0 ± 0 ns | 0.0 ± 0 ns | 0 ± 0 ns | ||
| Diet 3 | 4.0 ± 0 ns | 0 ± 0 ns | 0.33 ± 0.5 ns | ||
| Diet 4 | 3.83 ± 0.4 ns | 0.16 ± 0.4 ns | 0.66 ± 0.8 * | ||
| Control | 4.0 ± 0 | 0 ± 0 | 0 ± 0 | ||
| 5 | Diet 1 | 2.5 ± 1.5 ns | 1.66 ± 1.9 ns | 3.0 ± 2.1 ns | |
| Diet 2 | 1.0 ± 1.5 ** | 3.0 ± 1.5 ** | 6 ± 3.7 *** | ||
| Diet 3 | 0.5 ± 0.8 *** | 3.5 ± 0.8 *** | 4.16 ± 0.8 ** | ||
| Diet 4 | 1.0 ± 1.5 ** | 3.0 ± 1.5 ** | 4.0 ± 1.1 ** | ||
| Control | 3.66 ± 0.8 | 0.33 ± 0.8 | 0.5 ± 0.5 | ||
| 7.5 | Diet 1 | 0.83 ± 1.0 * | 3.16 ± 1.0 ** | 4.5 ± 1.2 ** | |
| Diet 2 | 0.0 ± 0 ** | 4.0 ± 0 *** | 9.8 ± 1.2 *** | ||
| Diet 3 | 1.16 ± 1.2 ns | 2.83 ± 1.2 ns | 4.33 ± 1.0 ** | ||
| Diet 4 | 0.66 ± 1.6 ** | 3.33 ± 1.6 ** | 4.83 ± 2.0 *** | ||
| Control | 2.83 ± 1.3 | 1.16 ± 1.3 | 1.5 ± 1.6 | ||
| 15 | Diet 1 | 0.66 ± 1.2 ** | 3.33 ± 1.2 ** | 3.66 ± 1.6* | |
| Diet 2 | 0.4 ± 0.5** | 3.6 ± 0.5 ** | 7.2 ± 0.8 *** | ||
| Diet 3 | 2.16 ± 1.8 ns | 1.83 ± 1.8 ns | 3.66 ± 2.0* | ||
| Diet 4 | 0.5 ± 0.8 ** | 3.5 ± 0.8 ** | 3.66 ± 1.9 * | ||
| Control | 3.16 ± 1.3 | 0.8 ± 1.3 | 1.17 ± 1.2 | ||
| 20 | Diet 1 | 2.5 ± 2.0 ns | 1.5 ± 2.0 ns | 3.16 ± 2.1* | |
| Diet 2 | 1.16 ± 1.2 ns | 2.83 ± 1.2 ns | 4.5 ± 2.0 *** | ||
| Diet 3 | 1.83 ± 1.2 ns | 2.16 ± 1.2 ns | 3.33 ± 1.0 * | ||
| Diet 4 | 1.5 ± 1.5 ns | 2.5 ± 1.5 ns | 2.66 ± 0.8 ns | ||
| Control | 2.16 ± 2.0 | 1.83 ± 2.0 | 1.0 ± 0.9 | ||
| 30 | Diet 1 | 4.0 ± 0 ns | 0 ± 0 ns | 0.83 ± 1.2 ns | |
| Diet 2 | 3.33 ± 0.8 ns | 0.66 ± 0.8 ns | 0.33 ± 0.8 ns | ||
| Diet 3 | 2.83 ± 1.5 ns | 1.16 ± 1.5 ns | 1.83 ± 1.5 * | ||
| Diet 4 | 3.83 ± 0.4 ns | 0.16 ± 0.4 ns | 0.66 ± 0.8 ns | ||
| Control | 3.44 ± 0.9 | 0.55 ± 0.9 | 0.33 ± 0.5 | ||
| Main effect p-value | |||||
| SL | *** | *** | *** | ||
| DT | *** | *** | *** | ||
| Day | ns | ns | *** | ||
| Day*SL*DT | *** | *** | *** |
| Treatment (ng) | Day | NDA | NLA | NN |
|---|---|---|---|---|
| 100 | 2 | 0 ± 0 ns | 4 ± 0 ns | 2.5 ± 0.7 ns |
| 3 | 0 ± 0 ns | 4 ± 0 ns | 3.5 ± 0.7 ns | |
| 7 | 0.5 ± 0.7 ns | 3.5 ± 0.7 *** | 6 ± 1.4 ns | |
| 200 | 2 | 0.5 ± 0.7 ns | 3.5 ± 0.7 ns | 2.5 ± 2.1 ns |
| 3 | 0.6 ± 0.9 ns | 3.6 ± 1.1 ns | 3.4 ± 1.5 ns | |
| 7 | 1.0 ± 1.4 ns | 3.0 ± 1.4 *** | 7.0 ± 4.2 ns | |
| 300 | 2 | 0 ± 0 ns | 4.0 ± 0 ns | 3.0 ± 1.4 ns |
| 3 | 1.0 ± 0 ns | 3.0 ± 0 ns | 2.5 ± 0.7 ns | |
| 7 | 1.0 ± 1.4 ns | 3.0 ± 1.4 *** | 4.5 ± 2.1 *** | |
| 500 | 2 | 1.0 ± 1.4 ns | 3.0 ± 1.4 ns | 1.5 ± 0.7 ns |
| 3 | 2.83 ± 1.8 *** | 1.16 ± 1.8 *** | 0.66 ± 1.2 *** | |
| 7 | 3.5 ± 0.7 ** | 0.5 ± 0.7 ** | 3.5 ± 2.1 * | |
| Control | 2 | 0 ± 0 | 4.0 ± 0 | 4.0 ± 0.7 |
| 3 | 0.11 ± 0.3 | 3.88 ± 0.3 | 5.33 ± 2.5 | |
| 7 | 0.43 ± 0.5 | 3.57 ± 0.5 | 8.86 ± 1.9 | |
| Main effect p-value | ||||
| Treatment | *** | *** | *** | |
| Day | * | * | *** | |
| Treatment * Day | ns | ns | ns |
| Composition | Quantity (mg)/100 mL Diet | |||
|---|---|---|---|---|
| Diet 1 | Diet 2 | Diet 3 | Diet 4 | |
| di-potassium hydrogen orthophosphate | 750.0 | 750.0 | 750.0 | 750.0 |
| magnesium sulfate | - | 123.0 | - | 123.0 |
| magnesium chloride | 123.0 | - | 123.0 | - |
| L-tyrosine | 40.0 | 40.0 | 40.0 | 40.0 |
| L-asparagine hydrate | 550.0 | 550.0 | 550.0 | 550.0 |
| L-aspartic acid | 140.0 | 140.0 | 140.0 | 140.0 |
| L-tryptophan | 80.0 | 80.0 | 80.0 | 80.0 |
| L-alanine | 100.0 | 100.0 | 100.0 | 100.0 |
| L-arginine monohydrochloride | 270.0 | 270.0 | 270.0 | 270.0 |
| L-cysteine hydrochloride, hydrate | 40.0 | 40.0 | 40.0 | 40.0 |
| L-glutamic acid | 140.0 | 140.0 | 140.0 | 140.0 |
| L-glutamine | 150.0 | 150.0 | 150.0 | 150.0 |
| L-glycine | 80.0 | 80.0 | 80.0 | 80.0 |
| L-histidine | 80.0 | 80.0 | 80.0 | 80.0 |
| L-isoleucine (allo free) | 80.0 | 80.0 | 80.0 | 80.0 |
| L-Leucine | 80.0 | 80.0 | 80.0 | 80.0 |
| L-Lysine monohydrochloride | 120.0 | 120.0 | 120.0 | 120.0 |
| L-methionine | 40.0 | 40.0 | 40.0 | 40.0 |
| L-phenylalanine | 40.0 | 40.0 | 40.0 | 40.0 |
| L-proline | 80.0 | 80.0 | 80.0 | 80.0 |
| L-serine | 80.0 | 80.0 | 80.0 | 80.0 |
| L-threonine | 140.0 | 140.0 | 140.0 | 140.0 |
| L-valine | 80.0 | 80.0 | 80.0 | 80.0 |
| ascorbic acid (vitamin C) | 100.0 | 100.0 | 100.0 | 100.0 |
| aneurine hydrochloride (vitamin B) | 2.5 | 2.5 | 2.5 | 2.5 |
| nicotinic acid | 10.0 | 10.0 | 10.0 | 10.0 |
| folic acid | 0.5 | 0.5 | 0.5 | 0.5 |
| (+)-pathothenic acid (calcium salt) | 5.0 | 5.0 | 5.0 | 5.0 |
| myo-inositol | 50.0 | 50.0 | 50.0 | 50.0 |
| choline chloride | 50.0 | 50.0 | 50.0 | 50.0 |
| EDTA Fe(III)-Na chelate pure | 1.5 | - | - | - |
| EDTA Zn-Na2 chelate pure | 0.8 | - | - | - |
| MnCl2.4H2O | 0.8 | - | - | - |
| EDTA Cu-Na2 chelate pure | 0.4 | - | - | - |
| FeSO4.7H2O | - | 1.5 | - | - |
| ZnSO4.7H2O | - | 0.8 | - | - |
| MnSO4.H2O | - | 0.8 | - | - |
| CuSO4.5H2O | - | 0.4 | - | - |
| pyridoxine hydrochloride (vitamin B6) | 2.5 | 2.5 | 2.5 | 2.5 |
| D-biotin, crystalline | 0.1 | 0.1 | 0.1 | 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jekayinoluwa, T.; Tripathi, J.N.; Dugdale, B.; Obiero, G.; Muge, E.; Dale, J.; Tripathi, L. Transgenic Expression of dsRNA Targeting the Pentalonia nigronervosa acetylcholinesterase Gene in Banana and Plantain Reduces Aphid Populations. Plants 2021, 10, 613. https://doi.org/10.3390/plants10040613
Jekayinoluwa T, Tripathi JN, Dugdale B, Obiero G, Muge E, Dale J, Tripathi L. Transgenic Expression of dsRNA Targeting the Pentalonia nigronervosa acetylcholinesterase Gene in Banana and Plantain Reduces Aphid Populations. Plants. 2021; 10(4):613. https://doi.org/10.3390/plants10040613
Chicago/Turabian StyleJekayinoluwa, Temitope, Jaindra Nath Tripathi, Benjamin Dugdale, George Obiero, Edward Muge, James Dale, and Leena Tripathi. 2021. "Transgenic Expression of dsRNA Targeting the Pentalonia nigronervosa acetylcholinesterase Gene in Banana and Plantain Reduces Aphid Populations" Plants 10, no. 4: 613. https://doi.org/10.3390/plants10040613
APA StyleJekayinoluwa, T., Tripathi, J. N., Dugdale, B., Obiero, G., Muge, E., Dale, J., & Tripathi, L. (2021). Transgenic Expression of dsRNA Targeting the Pentalonia nigronervosa acetylcholinesterase Gene in Banana and Plantain Reduces Aphid Populations. Plants, 10(4), 613. https://doi.org/10.3390/plants10040613