
Plant-Based Biostimulant as Sustainable Alternative to Synthetic Growth Regulators in Two Sweet Cherry Cultivars

 ,
, 
 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Results
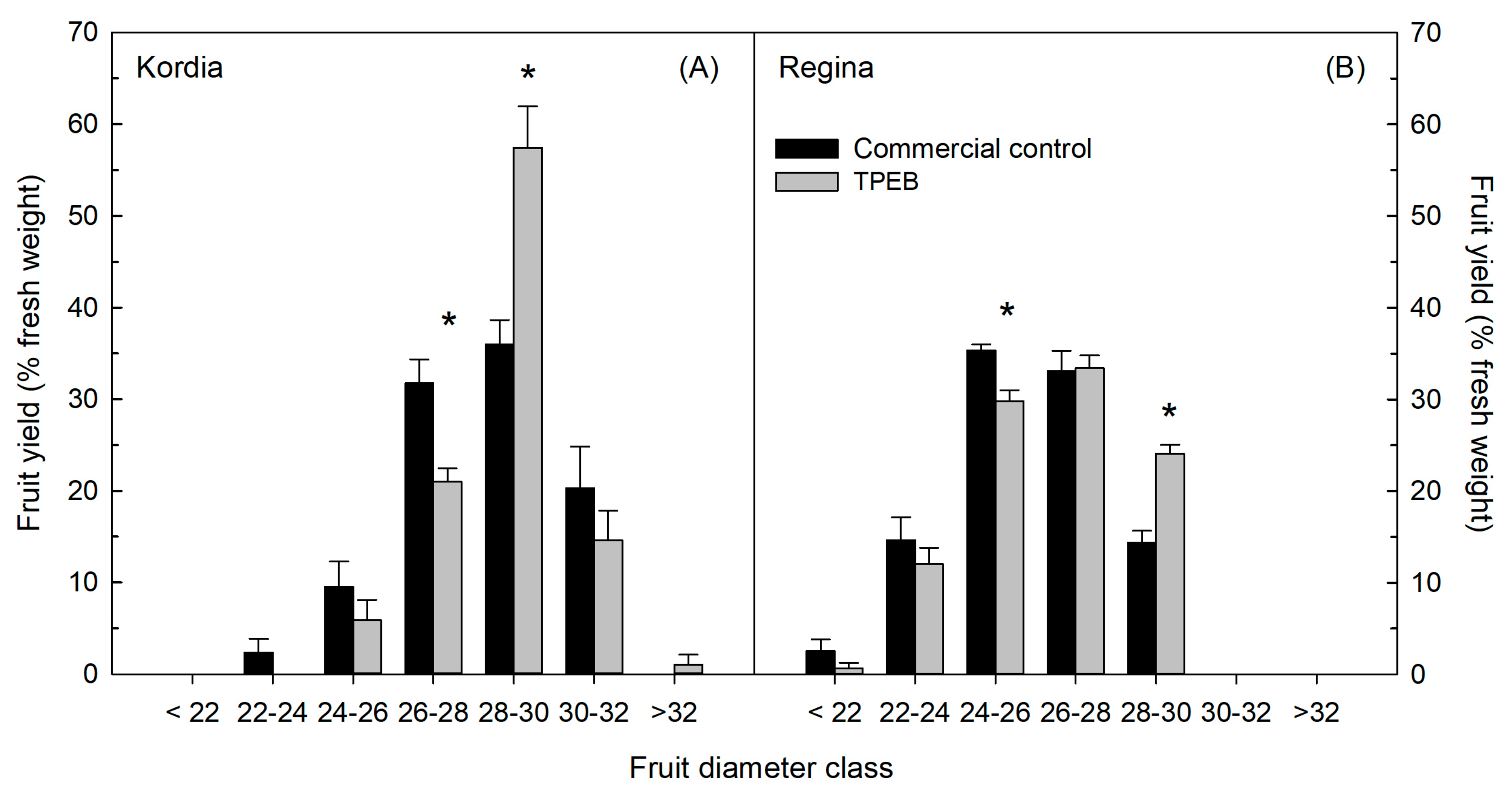
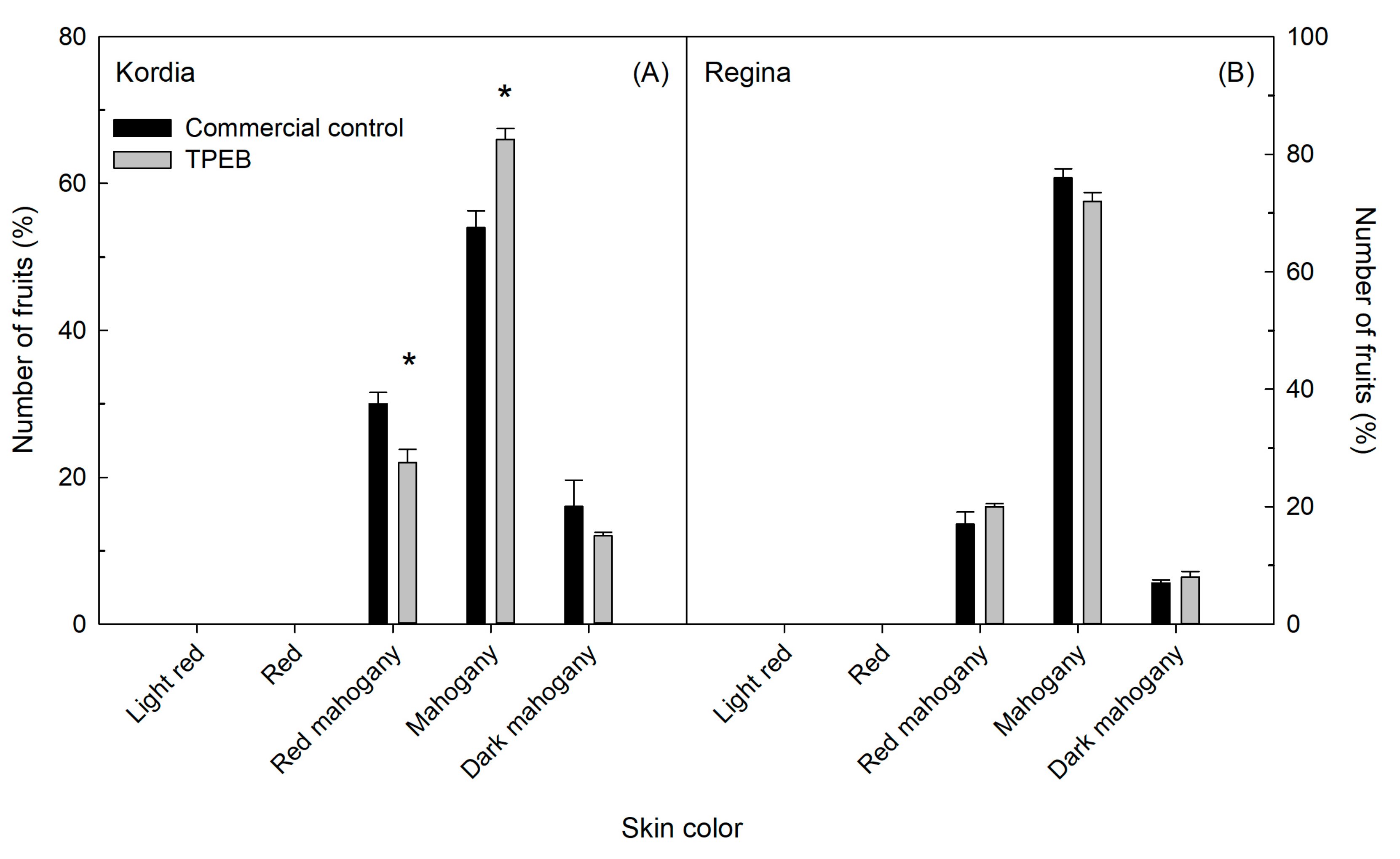
2.1. Fruit Yield, Fruit Fresh Weight, Size and Qualitative Traits
2.2. Composition of Leaf Petiole Sap
3. Discussion
4. Materials and Methods
4.1. Experimental Site, Plant Material and Experimental Design
4.2. Harvest and Fruit Yield
4.3. Fruit Fresh Weight, Size and Qualitative Traits
4.4. Composition of Leaf Petiole Sap
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAOSTAT. Agriculture Data. Available online: www.fao.org/faostat/en/#data/QC (accessed on 25 January 2021).
- Chauvin, M.A.; Whiting, M.; Ross, C.F. The Influence of Harvest Time on Sensory Properties and Consumer Acceptance of Sweet Cherries. HortTechnology 2009, 19, 748–754. [Google Scholar] [CrossRef] [Green Version]
- Cliff, M.A.; Dever, M.C.; Hall, J.W.; Girard, B. Development and Evaluation of Multiple Regression Models for Prediction of Sweet Cherry Liking. Int. Food Res. J. 1995, 28, 583–589. [Google Scholar] [CrossRef]
- Crisosto, C.H.; Crisosto, G.M.; Metheney, P. Consumer Acceptance of “Brooks” and “Bing” Cherries Is Mainly Dependent on Fruit SSC and Visual Skin Color. Postharvest Biol. Technol. 2003, 28, 159–167. [Google Scholar] [CrossRef]
- Hampson, C.R.; Stanich, K.; McKenzie, D.L.; Herbert, L.; Lu, R.; Li, J.; Cliff, M.A. Determining the Optimum Firmness for Sweet Cherries Using Just-About-Right Sensory Methodology. Postharvest Biol. Technol. 2014, 91, 104–111. [Google Scholar] [CrossRef]
- Belge, B.; Goulao, L.F.; Comabella, E.; Graell, J.; Lara, I. Refrigerated Storage and Calcium Dips of Ripe ‘Celeste’ Sweet Cherry Fruit: Combined Effects on Cell Wall Metabolism. Sci. Hortic. 2017, 219, 182–190. [Google Scholar] [CrossRef] [Green Version]
- Correia, S.; Queirós, F.; Ribeiro, C.; Vilela, A.; Aires, A.; Barros, A.I.; Schouten, R.; Silva, A.P.; Gonçalves, B. Effects of Calcium and Growth Regulators on Sweet Cherry (Prunus Avium L.) Quality and Sensory Attributes at Harvest. Sci. Hortic. 2019, 248, 231–240. [Google Scholar] [CrossRef]
- Romano, G.S.; Cittadini, E.D. Sweet Cherry Quality in the Horticultural Production Chain. Stewart Postharvest Rev. 2006, 6, 1–9. [Google Scholar] [CrossRef]
- Zhang, C.; Whiting, M.D. Improving “Bing” Sweet Cherry Fruit Quality with Plant Growth Regulators. Sci. Hortic. 2011, 127, 341–346. [Google Scholar] [CrossRef]
- Whiting, M.D.; Lang, G.; Ophardt, D. Rootstock and Training System Affect Sweet Cherry Growth, Yield, and Fruit Quality. HortScience 2005, 40, 582–586. [Google Scholar] [CrossRef] [Green Version]
- Simon, G. Review on Rain Induced Fruit Cracking of Sweet Cherries (Prunus Avium L.), Its Causes and the Possibilities of Prevention. Int. J. Hortic. 2006, 12, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Guyer, D.; Sinha, N.; Chang, T.-S.; Cash, J.N. Physicochemical and Sensory Characteristics of Fish Gelatin. J. Food Qual. 1993, 16, 355–370. [Google Scholar] [CrossRef]
- Tsantili, E.; Rouskas, D.; Christopoulos, M.V.; Stanidis, V.; Akrivos, J.; Papanikolaou, D. Effects of Two Pre-Harvest Calcium Treatments on Physiological and Quality Parameters in “Vogue” Cherries during Storage. J. Hortic. Sci. Biotechnol. 2007, 82, 657–663. [Google Scholar] [CrossRef]
- Einhorn, T.C.; Wang, Y.; Turner, J. Sweet Cherry Fruit Firmness and Postharvest Quality of Late-Maturing Cultivars Are Improved with Low-Rate, Single Applications of Gibberellic Acid. HortScience 2013, 48, 1010–1017. [Google Scholar] [CrossRef]
- Choi, C.; Wiersma, P.A.; Toivonen, P.; Kappel, F. Fruit Growth, Firmness and Cell Wall Hydrolytic Enzyme Activity during Development of Sweet Cherry Fruit Treated with Gibberellic Acid (GA3). J. Hortic. Sci. Biotechnol. 2002, 77, 615–621. [Google Scholar] [CrossRef]
- Ozkan, Y.; Ucar, M.; Yildiz, K.; Ozturk, B. Pre-Harvest Gibberellic Acid (GA3) Treatments Play an Important Role on Bioactive Compounds and Fruit Quality of Sweet Cherry Cultivars. Sci. Hortic. 2016, 211, 358–362. [Google Scholar] [CrossRef]
- Li, M.; Cheng, S.; Wang, Y.; Dong, Y. Improving Fruit Coloration, Quality Attributes, and Phenolics Content in ‘Rainier’ and ‘Bing’ Cherries by Gibberellic Acid Combined with Homobrassinolide. J. Plant Nutr. 2020, 39, 1130–1139. [Google Scholar] [CrossRef]
- Dong, Y.; Zhi, H.; Wang, Y. Cooperative Effects of Pre-Harvest Calcium and Gibberellic Acid on Tissue Calcium Content, Quality Attributes, and in Relation to Postharvest Disorders of Late-Maturing Sweet Cherry. Sci. Hortic. 2019, 246, 123–128. [Google Scholar] [CrossRef]
- Wójcik, P.; Akgül, H.; Demirtaş, I.; Sarisu, C.; Aksu, M.; Gubbuk, H. Effect of Preharvest Sprays of Calcium Chloride and Sucrose on Cracking and Quality of “Burlat” Sweet Cherry Fruit. J. Plant Nutr. 2013, 36, 1453–1465. [Google Scholar] [CrossRef]
- Stern, R.A.; Flaishman, M.; Applebaum, S.; Ben-Arie, R. Effect of Synthetic Auxins on Fruit Development of “Bing” Cherry (Prunus Avium L.). Sci. Hortic. 2007, 114, 275–280. [Google Scholar] [CrossRef]
- Hocking, B.; Tyerman, S.D.; Burton, R.A.; Gilliham, M. Fruit Calcium: Transport and Physiology. Front. Plant Sci. 2016, 7, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Peșteanu, A.; Balan, V.; Ivanov, I. Effect of Auxiger Growth Regulator on Fruits Development, Production and Chracing Index of “Regina” Cherry Variety. Sci. Papers. Series B Hortic. 2017, 11, 137–142. [Google Scholar]
- Correia, S.; Santos, M.; Glińska, S.; Gapińska, M.; Matos, M.; Carnide, V.; Schouten, R.; Silva, A.P.; Gonçalves, B. Effects of Exogenous Compound Sprays on Cherry Cracking: Skin Properties and Gene Expression. J. Sci. Food Agric. 2020, 100, 2911–2921. [Google Scholar] [CrossRef]
- Bhattacharyya, C.; Roy, R.; Tribedi, P.; Ghosh, A.; Ghosh, A. Biofertilizers as Substitute to Commercial Agrochemicals; Butterworth-Heinemann: Oxford, UK, 2020; ISBN 9780081030172. [Google Scholar]
- Srivastav, A.L. Chemical Fertilizers and Pesticides: Role in Groundwater Contamination; Butterworth-Heinemann: Oxford, UK, 2020; ISBN 9780081030172. [Google Scholar]
- Bulgari, R.; Cocetta, G.; Trivellini, A.; Vernieri, P.; Ferrante, A. Biostimulants and Crop Responses: A Review. Biol. Agric. Hortic. 2015, 31, 1–17. [Google Scholar] [CrossRef]
- De Pascale, S.; Rouphael, Y.; Colla, G. Plant Biostimulants: Innovative Tool for Enhancing Plant Nutrition in Organic Farming. Eur. J. Hortic. Sci. 2017, 82, 277–285. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Front. Plant Sci. 2020, 11, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Mataffo, A.; Scognamiglio, P.; Dente, A.; Strollo, D.; Colla, G.; Rouphael, Y.; Basile, B. Foliar Application of an Amino Acid-Enriched Urea Fertilizer on ‘Greco’ Grapevines at Full Veraison Increases Berry Yeast-Assimilable Nitrogen Content. Plants 2020, 9, 619. [Google Scholar] [CrossRef]
- Basile, B.; Rouphael, Y.; Colla, G.; Soppelsa, S.; Andreotti, C. Appraisal of Emerging Crop Management Opportunities in Fruit Trees, Grapevines and Berry Crops Facilitated by the Application of Biostimulants. Sci. Hortic. 2020, 267, 109330. [Google Scholar] [CrossRef]
- Vercammen, J.; Van Daele, G.; Vanrykel, T. Cracking of Sweet Cherries: Past Tense? Acta Hortic. 2008, 795 Pt 2, 463–468. [Google Scholar] [CrossRef]
- Martínez-Gil, A.M.; Angenieux, M.; Pardo-García, A.I.; Alonso, G.L.; Ojeda, H.; Rosario Salinas, M. Glycosidic Aroma Precursors of Syrah and Chardonnay Grapes after an Oak Extract Application to the Grapevines. Food Chem. 2013, 138, 956–965. [Google Scholar] [CrossRef]
- Pardo-García, A.I.; Martínez-Gil, A.M.; Cadahía, E.; Pardo, F.; Alonso, G.L.; Salinas, M.R. Oak Extract Application to Grapevines as a Plant Biostimulant to Increase Wine Polyphenols. Int. Food Res. J. 2014, 55, 150–160. [Google Scholar] [CrossRef]
- Sánchez-Gómez, R.; Garde-Cerdán, T.; Zalacain, A.; Garcia, R.; Cabrita, M.J.; Salinas, M.R. Vine-Shoot Waste Aqueous Extract Applied as Foliar Fertilizer to Grapevines: Effect on Amino Acids and Fermentative Volatile Content. Food Chem. 2016, 197, 132–140. [Google Scholar] [CrossRef]
- Sánchez-Gómez, R.; Zalacain, A.; Pardo, F.; Alonso, G.L.; Salinas, M.R. An Innovative Use of Vine-Shoots Residues and Their “Feedback” Effect on Wine Quality. Innov. Food Sci. Emerg. Technol. 2016, 37, 18–26. [Google Scholar] [CrossRef]
- Sánchez-Gómez, R.; Alonso, G.L.; Salinas, M.R.; Zalacain, A. Reuse of Vine-Shoots Wastes for Agricultural Purposes. In Handbook of Grape Processing by-Products-Sustainable Solutions; Elsevier: London, UK, 2017; pp. 79–104. [Google Scholar]
- Nasir, M.; Khan, A.S.; Basra, S.M.A.; Malik, A.U. Foliar Application of Moringa Leaf Extract, Potassium and Zinc Influence Yield and Fruit Quality of ‘Kinnow’ Mandarin. Sci. Hortic. 2016, 210, 227–235. [Google Scholar] [CrossRef]
- Colla, G.; Cardarelli, M.; Bonini, P.; Rouphael, Y. Foliar Applications of Protein Hydrolysate, Plant and Seaweed Extracts Increase Yield but Differentially Modulate Fruit Quality of Greenhouse Tomato. HortScience 2017, 52, 1214–1220. [Google Scholar] [CrossRef]
- Gillaspy, G.; Ben-david, H.; Gruissem, W.; Darwin, C. Fruits: A Developmental Perspective. Plant Cell 1993, 5, 1439–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, R.; Khurana, A.; Sharma, A.K. Role of Plant Hormones and Their Interplay in Development and Ripening of Fleshy Fruits. J. Exp. Bot. 2014, 65, 4561–4575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenn, M.A.; Giovannoni, J.J. Phytohormones in Fruit Development and Maturation. Plant J. 2020, 446–458. [Google Scholar] [CrossRef]
- Wen, B.; Song, W.; Sun, M.; Chen, M.; Mu, Q.; Zhang, X.; Chen, X.; Gao, D.; Xiao, W. Identification and Characterization of Cherry (Cerasus Pseudocerasus G. Don) Genes in Response to Parthenocarpy Induced by GA3 through Transcriptome Analysis. BMC Genet. 2019, 20, 65. [Google Scholar] [CrossRef]
- Colla, G.; Nardi, S.; Cardarelli, M.; Ertani, A.; Lucini, L.; Canaguier, R.; Rouphael, Y. Protein Hydrolysates as Biostimulants in Horticulture. Sci. Hortic. 2015, 196, 28–38. [Google Scholar] [CrossRef]
- Colla, G.; Hoagland, L.; Ruzzi, M.; Cardarelli, M.; Bonini, P.; Canaguier, R.; Rouphael, Y. Biostimulant Action of Protein Hydrolysates: Unraveling Their Effects on Plant Physiology and Microbiome. Front. Plant Sci. 2017, 8, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Sestili, F.; Rouphael, Y.; Cardarelli, M.; Pucci, A.; Bonini, P.; Canaguier, R.; Colla, G. Protein Hydrolysate Stimulates Growth in Tomato Coupled with N-Dependent Gene Expression Involved in N Assimilation. Front. Plant Sci. 2018, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Schiavon, M.; Ertani, A.; Nardi, S. Effects of an Alfalfa Protein Hydrolysate on the Gene Expression and Activity of Enzymes of the Tricarboxylic Acid (TCA) Cycle and Nitrogen Metabolism in Zea Mays L. J. Agric. Food Chem. 2008, 56, 11800–11808. [Google Scholar] [CrossRef]
- Ertani, A.; Cavani, L.; Pizzeghello, D.; Brandellero, E.; Altissimo, A.; Ciavatta, C.; Nardi, S. Biostimulant Activity of Two Protein Hydrolyzates in the Growth and Nitrogen Metabolism of Maize Seedlings. J. Plant Nutr. Soil Sci. 2009, 172, 237–244. [Google Scholar] [CrossRef]
- Ertani, A.; Schiavon, M.; Muscolo, A.; Nardi, S. Alfalfa Plant-Derived Biostimulant Stimulate Short-Term Growth of Salt Stressed Zea Mays L. Plants. Plant Soil. 2013, 364, 145–158. [Google Scholar] [CrossRef]
- Ertani, A.; Schiavon, M.; Nardi, S. Transcriptome-Wide Identification of Differentially Expressed Genes in Solanum Lycopersicon L. In Response to an Alfalfa-Protein Hydrolysate Using Microarrays. Front. Plant Sci. 2017, 8, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouphael, Y.; Cardarelli, M.; Bonini, P.; Colla, G. Synergistic Action of a Microbial-Based Biostimulant and a Plant Derived-Protein Hydrolysate Enhances Lettuce Tolerance to Alkalinity and Salinity. Front. Plant Sci. 2017, 8, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagredo, K.X.; Cassasa, V.; Vera, R.; Carroza, I. Pollination and Fruit Set for “Kordia” and “Regina” Sweet Cherry Trees in the South of Chile. Acta Hortic. 2017, 1161, 353–359. [Google Scholar] [CrossRef]
- Whiting, M.D.; Lang, G.A. “Bing” Sweet Cherry on the Dwarfing Rootstock “Gisela 5”: Thinning Affects Fruit Quality and Vegetative Growth but Not Net CO2 Exchange. J. Am. Soc. Hortic. Sci. 2004, 129, 407–415. [Google Scholar] [CrossRef] [Green Version]
- Azarenko, A.N.; Chozinski, A.; Brewer, L.J. Fruit Growth Curve Analysis of Seven Sweet Cherry Cultivars. Acta Hortic. 2008, 795, 561–566. [Google Scholar] [CrossRef]
- Basile, B.; Mariscal, M.J.; Day, K.R.; Johnson, R.S.; DeJong, T.M. Japanese Plum (Prunus Salicina L.) Fruit Growth: Seasonal Pattern of Source/Sink Limitations. J. Am. Pomol. Soc. 2002, 56, 86–93. [Google Scholar]
- Tukey, H.B.; Young, J.O. Histological Study of the Developing Fruit of the Sour Cherry. Bot. Gaz 1939, 100, 723–749. [Google Scholar] [CrossRef]
- Coombe, B.G. The Development of Fleshy Fruits. Annu. Rev. Plant Biol 1976, 27, 207–228. [Google Scholar] [CrossRef]
- Basile, B.; Solari, L.I.; Dejong, T.M. Intra-Canopy Variability of Fruit Growth Rate in Peach Trees Grafted on Rootstocks with Different Vigour-Control Capacity. J. Hortic. Sci. Biotechnol. 2007, 82, 243–256. [Google Scholar] [CrossRef]
- Iwahori, S.; Tominaga, S.; Yamasaki, T. Stimulation of Fruit Growth of Kiwifruit, Actinidia Chinensis Planch., by N-(2-Chloro-4-Pyridyl)-N’-Phenylurea, a Diphenylurea-Derivative Cytokinin. Sci. Hortic. 1988, 35, 109–115. [Google Scholar] [CrossRef]
- Zhang, C.; Lee, U.; Tanabe, K. Hormonal Regulation of Fruit Set, Parthenogenesis Induction and Fruit Expansion in Japanese Pear. Plant Growth Regul. 2008, 55, 231–240. [Google Scholar] [CrossRef]
- Stirk, W.A.; Van Staden, J. Comparison of Cytokinin- and Auxin-like Activity in Some Commercially Used Seaweed Extracts. J. Appl. Phycol. 1996, 8, 503–508. [Google Scholar] [CrossRef]
- Pizzeghello, D.; Francioso, O.; Ertani, A.; Muscolo, A.; Nardi, S. Isopentenyladenosine and Cytokinin-like Activity of Different Humic Substances. J. Geochem. Explor. 2013, 129, 70–75. [Google Scholar] [CrossRef]
- Wang, Y.; Long, L.E. Physiological and Biochemical Changes Relating to Postharvest Splitting of Sweet Cherries Affected by Calcium Application in Hydrocooling Water. Food Chem. 2015, 181, 241–247. [Google Scholar] [CrossRef]
- Caccavello, G.; Giaccone, M.; Scognamiglio, P.; Forlani, M.; Basile, B. Influence of Intensity of Post-Veraison Defoliation or Shoot Trimming on Vine Physiology, Yield Components, Berry and Wine Composition in Aglianico Grapevines. Aust. J. Grape Wine Res. 2017, 23, 226–239. [Google Scholar] [CrossRef]
- Larronde, F.; Krisa, S.; Decendit, A.; Chèze, C.; Deffieux, G.; Mérillon, J.M. Regulation of Polyphenol Production in Vitis Vinifera Cell Suspension Cultures by Sugars. Plant Cell Reps. 1998, 17, 946–950. [Google Scholar] [CrossRef]
- Vitrac, X.; Larronde, F.; Krisa, S.; Decendit, A.; Deffieux, G.; Mérillon, J.M. Sugar Sensing and Ca2+-Calmodulin Requirement in Vitis Vinifera Cells Producing Anthocyanins. Phytochemistry 2000, 53, 659–665. [Google Scholar] [CrossRef]
- Ertani, A.; Pizzeghello, D.; Francioso, O.; Sambo, P.; Sanchez-Cortes, S.; Nardi, S. Capsicum Chinensis L. Growth and Nutraceutical Properties Are Enhanced by Biostimulants in a Long-Term Period: Chemical and Metabolomic Approaches. Front. Plant Sci. 2014, 5, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landi, M.; Massai, R.; Remorini, D. Effect of Rootstock and Manual Floral Bud Thinning on Organoleptical and Nutraceutical Properties of Sweet Cherry (Prunus Avium L) Cv “Lapins”. Agrochimica 2014, 58, 335–351. [Google Scholar] [CrossRef]
- Kurlus, R.; Rutkowski, K.; Łysiak, G.P. Improving of Cherry Fruit Quality and Bearing Regularity by Chemical Thinning with Fertilizer. Agronomy 2020, 10, 1281. [Google Scholar] [CrossRef]
- Mataffo, A.; Scognamiglio, P.; Basile, B.; Lisanti, M.T.; Tenore, G.C.; Graziani, G.; Ritieni, A.; Giaccone, M. Crop Load Affects the Nutritional Quality of Flat Peaches (Prunus Persica L. Var. Platycarpa (Decne.) L.H. Bailey). Italus Hortus 2020, 27, 41–54. [Google Scholar] [CrossRef]
- Baggiolini, M. Les Stades Repérés Des Arbres Fruitiers à Noyau. Rev. Romande D’Agric. D’Arboric. 1952, 8, 4–6. [Google Scholar]
- Fadón, E.; Herrero, M.; Rodrigo, J. Flower Development in Sweet Cherry Framed in the BBCH Scale. Sci. Hortic. 2015, 192, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Hernandez, A.; Mateos, R.; Garcia-Mesa, J.A.; Beltran, G.; Fernandez-Escobar, R. Determination of Mineral Elements in Fresh Olive Fruits by Flame Atomic Spectrometry. Span. J. Agric. Res. 2010, 8, 1183. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Treatment | Kordia | Regina | ||
|---|---|---|---|---|
| Fruit Yield (tons/ha) | Crop Load (Number Fruits/Tree) | Fruit Yield (tons/ha) | Crop Load (Number Fruits/Tree) | |
| Commercial Control | 12.064 ± 0.308 | 1113 ± 28 | 26.914 ± 0.449 | 3134 ± 54 |
| TPEB | 12.954 ± 0.267 | 1136 ± 23 | 30.438 ± 0.304 | 3369 ± 34 |
| Significance | * | n.s. | ** | ** |
| Treatment | Kordia | Regina | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Fruit Diameter (mm) | Fruit Fresh Weight (g/Fruit) | SSC (°Brix) | Flesh Firmness (g/mm2) | Calcium Content (mm/100 g) | Fruit Diameter (mm) | Fruit Fresh Weight (g/Fruit) | SSC (°Brix) | Flesh Firmness (g/mm2) | Calcium Content (mm/100 g) | |
| Commercial Control | 27.84 ± 0.26 | 12.08 ± 0.21 | 18.31 ± 0.28 | 325.29 ± 7.29 | 12.47 ± 0.19 | 25.42 ± 0.19 | 9.67 ± 0.19 | 18.86 ± 0.26 | 322.77 ± 6.69 | 12.02 ± 0.29 |
| TPEB | 28.55 ± 0.16 | 11.39 ± 0.19 | 20.47 ± 0.37 | 347.09 ± 7.66 | 15.75 ± 0.18 | 25.97 ± 0.19 | 10.46 ± 0.20 | 17.46 ± 0.22 | 324.45 ± 7.00 | 12.80 ± 0.18 |
| Significance | * | * | *** | * | * | * | ** | *** | n.s. | n.s. |
| Treatment | Phenological Stage | |||
|---|---|---|---|---|
| Early Stage D “Sepals Open” | Stage F “Full Bloom” | Stage G ”Full Petal Fall” | 7 Days after Stage G | |
| Commercial control | Seaweed extract (3.75 L/ha) + Organic fertilizer (14% of calcium complexed with 6% of amino acids; 4.5 L/ha) | Cytokinin promoter (2 L/ha) + Organic fertilizer (14% of calcium complexed with 6% of amino acids; 4.5 L/ha) | Cytokinin promoter (2 L/ha) + Organic fertilizer (14% of calcium complexed with 6% of amino acids; 4.5 L/ha) | Cytokinin promoter (2 L/ha) + Organic fertilizer (14% of calcium complexed with 6% of amino acids; 4.5 L/ha) |
| TPEB | Seaweed extract (3.75 L/ha) + Organic fertilizer (5% of calcium, CaO, and 5% of vegetal amino acids; 3 L/ha) | Tropical-plant extract (1.5 L/ha) + Organic fertilizer (5% of calcium, CaO, and 5% of vegetal amino acids; 3 L/ha) | Tropical-plant extract (1.5 L/ha) + Organic fertilizer (5% of calcium, CaO, and 5% of vegetal amino acids; 3 L/ha) | Tropical-plant extract (1.5 L/ha) + Organic fertilizer (5% of calcium, CaO, and 5% of vegetal amino acids; 3 L/ha) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Basile, B.; Brown, N.; Valdes, J.M.; Cardarelli, M.; Scognamiglio, P.; Mataffo, A.; Rouphael, Y.; Bonini, P.; Colla, G. Plant-Based Biostimulant as Sustainable Alternative to Synthetic Growth Regulators in Two Sweet Cherry Cultivars. Plants 2021, 10, 619. https://doi.org/10.3390/plants10040619
Basile B, Brown N, Valdes JM, Cardarelli M, Scognamiglio P, Mataffo A, Rouphael Y, Bonini P, Colla G. Plant-Based Biostimulant as Sustainable Alternative to Synthetic Growth Regulators in Two Sweet Cherry Cultivars. Plants. 2021; 10(4):619. https://doi.org/10.3390/plants10040619
Chicago/Turabian StyleBasile, Boris, Natalie Brown, José Miguel Valdes, Mariateresa Cardarelli, Pasquale Scognamiglio, Alessandro Mataffo, Youssef Rouphael, Paolo Bonini, and Giuseppe Colla. 2021. "Plant-Based Biostimulant as Sustainable Alternative to Synthetic Growth Regulators in Two Sweet Cherry Cultivars" Plants 10, no. 4: 619. https://doi.org/10.3390/plants10040619
APA StyleBasile, B., Brown, N., Valdes, J. M., Cardarelli, M., Scognamiglio, P., Mataffo, A., Rouphael, Y., Bonini, P., & Colla, G. (2021). Plant-Based Biostimulant as Sustainable Alternative to Synthetic Growth Regulators in Two Sweet Cherry Cultivars. Plants, 10(4), 619. https://doi.org/10.3390/plants10040619