
The Distribution of Minerals in Crucial Plant Parts of Various Elderberry (Sambucus spp.) Interspecific Hybrids



Abstract
:1. Introduction
2. Results and Discussion
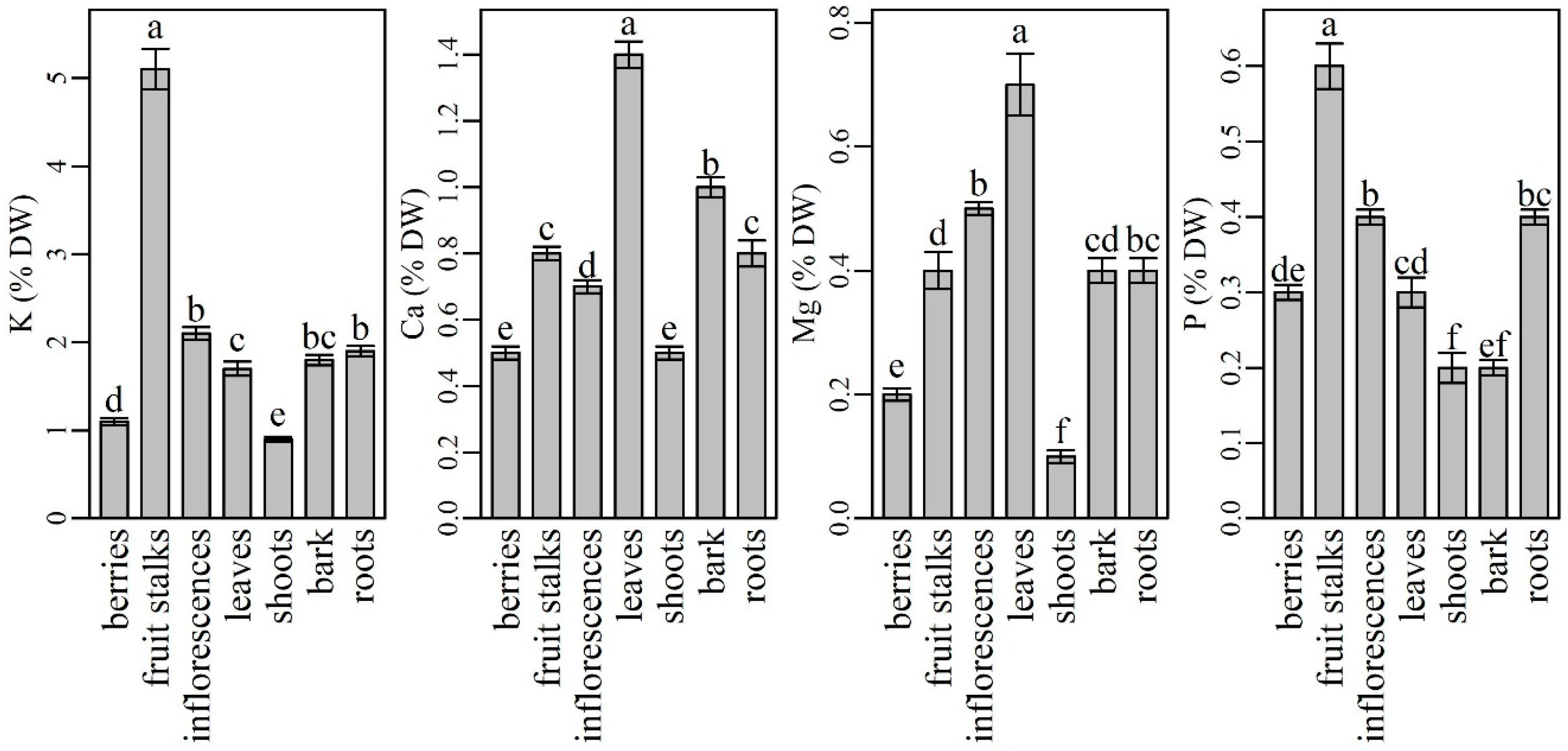
2.1. The Content and Distribution of Macronutrients in Various Plant Parts of Elderberry Interspecific Hybrids
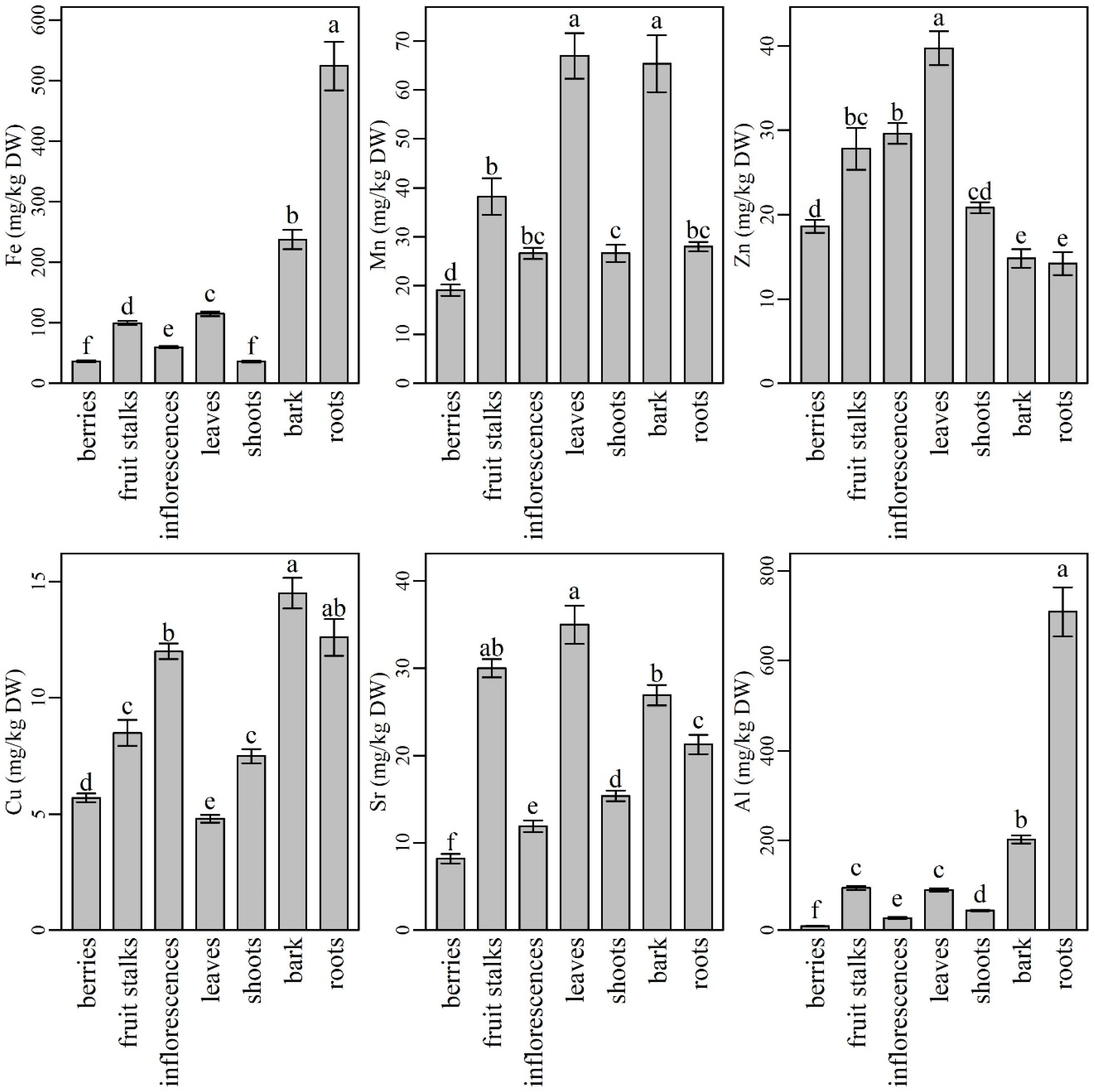
2.2. The Content and Distribution of Micronutrients, Aluminium and Strontium in Various Plant Parts of Elderberry Interspecific Hybrids
2.3. Clustering of Elderberry Interspecific Hybrids and Correlation for Minerals between Plant Parts
3. Materials and Methods
3.1. Plant Material—Elderberry Genotypes and Samples
3.2. Chemical Analysis
3.3. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marschner, H. Mineral Nutrition of Higher Plants, 2nd ed.; Academic Press: Cambridge, MA, USA, 1995. [Google Scholar]
- Barroso, M.F.; Silva, A.; Ramos, S.; Oliva-Teles, M.T.; Delerue-Matos, C.; Sales, M.G.F.; Oliveira, M. Flavoured versus natural waters: Macromineral (Ca, Mg, K, Na) and micromineral (Fe, Cu, Zn) contents. Food Chem. 2009, 116, 580–589. [Google Scholar] [CrossRef] [Green Version]
- Soetan, K.O.; Olaiya, C.O.; Oyewole, O.E. The importance of mineral elements for humans, domestic animals and plants-A review. Afr. J. Food Sci. 2010, 4, 200–222. [Google Scholar]
- Gharibzahedi, S.M.T.; Jafari, S.M. The importance of minerals in human nutrition: Bioavailability, food fortification, processing effects and nanoencapsulation. Trends Food Sci. Technol. 2017, 62, 119–132. [Google Scholar] [CrossRef]
- Stipanuk, M.H.; Caudill, M.A. Biochemical, Physiological, and Molecular Aspects of Human Nutrition, 3rd ed.; Elsevier/Saunders: St. Louis, MO, USA, 2013. [Google Scholar]
- White, P.J.; Broadley, M.R. Biofortification of crops with seven mineral elements often lacking in human diets–iron, zinc, copper, calcium, magnesium, selenium and iodine. New Phytol. 2009, 182, 49–84. [Google Scholar] [CrossRef]
- Gupta, U.C.; Gupta, S.C. Sources and deficiency diseases of mineral nutrients in human health and nutrition: A review. Pedosphere 2014, 24, 13–38. [Google Scholar] [CrossRef]
- Welch, R.M.; Graham, R.D. Breeding for micronutrients in staple food crops from a human nutrition perspective. J. Exp. Bot. 2004, 55, 353–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welch, R.M. Effects of nutrient deficiencies on seed production and quality. Adv. Plant Nutr. 1986, 2, 205–247. [Google Scholar]
- Welch, R.M. Importance of seed mineral nutrient reserves in crop growth and development. In Mineral Nutrition of Crops: Fundamental Mechanisms and Implications; Food Products Press: New York, NY, USA, 1999; pp. 205–226. [Google Scholar]
- Seabra, I.J.; Braga, M.E.M.; Batista, M.T.P.; de Sousa, H.C. Fractioned high pressure extraction of anthocyanins from elderberry (Sambucus nigra L.) pomace. Food Bioprocess Technol. 2010, 3, 674–683. [Google Scholar] [CrossRef] [Green Version]
- Jakobek, L.; Šeruga, M.; Novak, I.; Medvidović-Kosanović, M. Flavonols, phenolic acids and antioxidant activity of some red fruits. Dtsch. Lebensm. Rundsch. 2007, 103, 369–377. [Google Scholar]
- Rimpapa, Z.; Toromanovic, J.; Tahirovic, I.; Šapčanin, A.; Sofic, E. Total content of phenols and anthocyanins in edible fruits from Bosnia. Bosn. J. Basic Med. Sci. 2007, 7, 119–122. [Google Scholar] [CrossRef] [Green Version]
- Viskelis, P.; Rubinskiene, M.; Bobinaite, R.; Dambrauskiene, E. Bioactive compounds and antioxidant activity of small fruits in Lithuania. J. Food Agric. Environ. 2010, 8, 259–263. [Google Scholar]
- Mikulic-Petkovsek, M.; Slatnar, A.; Stampar, F.; Veberic, R. HPLC–MSn identification and quantification of flavonol glycosides in 28 wild and cultivated berry species. Food Chem. 2012, 135, 2138–2146. [Google Scholar] [CrossRef] [PubMed]
- Kiss, A.; Rapi, S.; Korozs, M.; Forgo, P. Elaboration of novel extraction procedure to reveal bioactive component profile of anthocyanin-rich plants. Czech J. Food Sci. 2014, 32, 384–390. [Google Scholar] [CrossRef] [Green Version]
- Dudonne, S.; Dube, P.; Anhe, F.F.; Pilon, G.; Marette, A.; Lemire, M.; Harris, C.; Dewailly, E.; Desjardins, Y. Comprehensive analysis of phenolic compounds and abscisic acid profiles of twelve native Canadian berries. J. Food Compos. Anal. 2015, 44, 214–224. [Google Scholar] [CrossRef]
- Klensporf-Pawlik, D.; Przybylski, R. Antioxidant activity of selected wild Canadian prairie fruits. Acta Sci. Pol. Technol. Aliment. 2015, 14, 357–366. [Google Scholar] [CrossRef]
- Cata, A.; Stefanut, M.N.; Pop, R.; Tanasie, C.; Mosoarca, C.; Zamfir, A.D. Evaluation of antioxidant activities of some small fruits containing anthocyanins using electrochemical and chemical methods. Croat. Chem. Acta 2016, 89, 37–48. [Google Scholar] [CrossRef]
- Tarko, T.; Duda-Chodak, A.; Sroka, P.; Satora, P.; Semik-Szczurak, D.; Wajda, L. Diversity and bioavailability of fruit polyphenols. J. Food Nutr. Res. 2017, 56, 167–178. [Google Scholar]
- Sidor, A.; Gramza-Michalowska, A. Advanced research on the antioxidant and health benefit of elderberry (Sambucus nigra) in food—A review. J. Funct. Foods 2015, 18, 941–958. [Google Scholar] [CrossRef]
- Młynarczyk, K.; Walkowiak-Tomczak, D.; Lysiak, G.P. Bioactive properties of Sambucus nigra L. as a functional ingredient for food and pharmaceutical industry. J. Funct. Foods 2018, 40, 377–390. [Google Scholar] [CrossRef]
- Bolli, R. Revision of the Genus Sambucus; J. Cramer: Berlin, Germany, 1994. [Google Scholar]
- Anton, A.M.; Pintea, A.M.; Rugina, D.O.; Sconta, Z.M.; Hanganu, D.; Vlase, L.; Benedec, D. Preliminary studies on the chemical characterization and antioxidant capacity of polyphenols from Sambucus sp. Dig. J. Nanomater. Biostruct. 2013, 8, 973–980. [Google Scholar]
- Veberic, R.; Jakopic, J.; Stampar, F.; Schmitzer, V. European elderberry (Sambucus nigra L.) rich in sugars, organic acids, anthocyanins and selected polyphenols. Food Chem. 2009, 114, 511–515. [Google Scholar] [CrossRef]
- Fernald, M.L. Gray’s Manual of Botany, 8th ed.; American Book Co.: New York, NY, USA, 1950. [Google Scholar]
- Akbulut, M.; Ercisli, S.; Tosun, M. Physico-chemical characteristics of some wild grown European elderberry (Sambucus nigra L.) genotypes. Pharmacogn. Mag. 2009, 5, 320–323. [Google Scholar]
- Imbrea, I.M.; Radulov, I.; Nicolin, A.L.; Imbrea, F. Analysis of macroelements content of some medicinal and aromatic plants using flame atomic absorption spectrometry (FAAS). Rom. Biotechnol. Lett. 2016, 21, 11641–11649. [Google Scholar]
- Mikulic-Petkovsek, M.; Ivancic, A.; Todorovic, B.; Veberic, R.; Stampar, F. Fruit phenolic composition of different elderberry species and hybrids. J. Food Sci. 2015, 80, 2180–2190. [Google Scholar] [CrossRef] [PubMed]
- Milena, V.; Tatjana, M.; Gökhan, Z.; Ivana, B.; Aleksandra, C.; Mohammad, M.F.; Marija, R. Advantages of contemporary extraction techniques for the extraction of bioactive constituents from black elderberry (Sambucus nigra L.) flowers. Ind. Crops Prod. 2019, 136, 93–101. [Google Scholar] [CrossRef]
- Mikulic-Petkovsek, M.; Samoticha, J.; Eler, K.; Stampar, F.; Veberic, R. Traditional elderflower beverages: A rich source of phenolic compounds with high antioxidant activity. J. Agric. Food Chem. 2015, 63, 1477–1487. [Google Scholar] [CrossRef]
- Shokrzadeh, M.; Saravi, S.S.S.; Mirzayi, M. Cytotoxic effects of ethyl acetate extract of Sambucus ebulus compared with etoposide on normal and cancer cell lines. Pharmacogn. Mag. 2009, 5, 316–319. [Google Scholar]
- Vlachojannis, J.E.; Cameron, M.; Chrubasik, S. A systematic review on the sambuci fructus effect and efficacy profiles. Phytother. Res. 2010, 24, 1–8. [Google Scholar] [CrossRef]
- Veberic, R.; Jakopic, J.; Stampar, F. Flavonols and Anthocyanins of Elderberry Fruits (Sambucus nigra L.). In Acta Horticulturae, II International Symposium on Human Health Effects of Fruits and Vegetables: Favhealth 2007; Patil, B., Ed.; International Society for Horticultural Science: Houston, TX, USA, 2009; pp. 611–614. [Google Scholar]
- Duymus, H.G.; Goger, F.; Baser, K.H.C. In vitro antioxidant properties and anthocyanin compositions of elderberry extracts. Food Chem. 2014, 155, 112–119. [Google Scholar] [CrossRef]
- Silva, P.; Ferreira, S.; Nunes, F.M. Elderberry (Sambucus nigra L.) by-products a source of anthocyanins and antioxidant polyphenols. Ind. Crops Prod. 2017, 95, 227–234. [Google Scholar] [CrossRef]
- Dawidowicz, A.L.; Wianowska, D.; Baraniak, B. The antioxidant properties of alcoholic extracts from Sambucus nigra L. (antioxidant properties of extracts). LWT—Food Sci. Technol. 2006, 39, 308–315. [Google Scholar] [CrossRef]
- Caruso, M.C.; Galgano, F.; Tolve, R.; Pecora, M.; Tedesco, I.; Favati, F.; Condelli, N. Nutraceutical properties of wild berry fruits from Southern Italy. J. Berry Res. 2016, 6, 321–332. [Google Scholar] [CrossRef] [Green Version]
- Młynarczyk, K.; Walkowiak-Tomczak, D.; Staniek, H.; Kidoń, M.; Łysiak, G.P. The Content of Selected Minerals, Bioactive Compounds, and the Antioxidant Properties of the Flowers and Fruit of Selected Cultivars and Wildly Growing Plants of Sambucus nigra L. Molecules 2020, 25, 876. [Google Scholar] [CrossRef] [Green Version]
- Tuncay, K.A.N. Some Biochemical Characteristics of Fruit Samples of Sambucus nigra Genotypes from Northeastern Turkey. Not. Bot. Horti Agrobot. Cluj-Napoca 2019, 47, 553–557. [Google Scholar]
- Antolak, H.; Czyzowska, A.; Kregiel, D. Antibacterial and Antiadhesive Activities of Extracts from Edible Plants against Soft Drink Spoilage by Asaia spp. J. Food Prot. 2017, 80, 25–34. [Google Scholar] [CrossRef]
- Vrchotova, N.; Dadakova, E.; Matejicek, A.; Triska, J.; Kaplan, J. Effect of variety on content of bioactive phenolic compounds in common elder (Sambucus nigra L.). Nat. Prod. Res. 2017, 31, 700–703. [Google Scholar] [CrossRef]
- Ozgen, M.; Scheerens, J.C.; Reese, R.N.; Miller, R.A. Total phenolic, anthocyanin contents and antioxidant capacity of selected elderberry (Sambucus canadensis L.) accessions. Pharmacogn. Mag. 2010, 6, 198–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Finn, C.E. Anthocyanins and other polyphenolics in American elderberry (Sambuclus canadensis) and European elderberry (S. nigra) cultivars. J. Sci. Food Agric. 2007, 87, 2665–2675. [Google Scholar] [CrossRef] [Green Version]
- Mudge, E.; Applequist, W.L.; Finley, J.; Lister, P.; Townesmith, A.K.; Walker, K.M.; Brown, P.N. Variation of select flavonols and chlorogenic acid content of elderberry collected throughout the Eastern United States. J. Food Compos. Anal. 2016, 47, 52–59. [Google Scholar] [CrossRef] [Green Version]
- Thomas, A.L.; Perkins-Veazie, P.; Byers, P.L.; Finn, C.E.; Lee, J. A comparison of fruit characteristics among diverse elderberry genotypes grown in Missouri and Oregon. J. Berry Res. 2013, 3, 159–168. [Google Scholar] [CrossRef] [Green Version]
- Diviš, P.; Porizka, J.; Vespalcová, M.; Matejiček, A.; Kaplan, J. Elemental composition of fruits from different Black elder (Sambucus nigra L.) cultivars grown in the Czech Republic. J. Elem. 2015, 20, 549–557. [Google Scholar] [CrossRef]
- Llorent-Martinez, E.J.; Spinola, V.; Castilho, P.C. Evaluation of the inorganic content of six underused wild berries from Portugal: Potential new sources of essential minerals. J. Food Compos. Anal. 2017, 59, 153–160. [Google Scholar] [CrossRef]
- Senica, M.; Stampar, F.; Veberic, R.; Mikulic-Petkovsek, M. The higher the better? Differences in phenolics and cyanogenic glycosides in Sambucus nigra leaves, flowers and berries from different altitudes. J. Sci. Food Agric. 2017, 97, 2623–2632. [Google Scholar] [CrossRef]
- Thomas, A.L.; Chen, Y.C.; Rottinghaus, G.E.; Malone, A.M.; Byers, P.L.; Applequist, W.L.; Finn, C.E. Occurrence of rutin and chlorogenic acid in elderberry leaf, flower, and stem in response to genotype, environment, and season. In Proceedings of the Acta Horticulturae, XXVII International Horticultural Congress-IHC2006: International Symposium on Plants as Food and Medicine: The Utilization and Development of Horticultural Plants for Human Health, Seoul, Korea, 13–19 August 2006; Gardner, L.E.C.G., Ed.; International Society for Horticultural Science: Houston, TX, USA, 2008; pp. 197–206. [Google Scholar]
- Prajapati, K.; Modi, H.A. The importance of potassium in plant growth-a review. Indian J. Plant Sci. 2012, 1, 177–186. [Google Scholar]
- Tagliavini, M.; Baldi, E.; Lucchi, P.; Antonelli, M.; Sorrenti, G.; Baruzzi, G.; Faedi, W. Dynamics of nutrients uptake by strawberry plants (Fragaria× Ananassa Dutch.) grown in soil and soilless culture. Eur. J. Agron. 2005, 23, 15–25. [Google Scholar] [CrossRef]
- Twyford, I.T.; Walmsley, D. The mineral composition of the Robusta banana plant II. The concentration of mineral constituents. Plant Soil 1974, 41, 459–470. [Google Scholar] [CrossRef]
- Kolodziej, B.; Maksymiec, N.; Drozdzal, K.; Antonkiewicz, J. Effect of traffic pollution on chemical composition of raw elderberry (Sambucus nigra L.). J. Elem. 2012, 17, 67–78. [Google Scholar] [CrossRef]
- He, F.J.; MacGregor, G.A. Beneficial effects of potassium on human health. Physiol. Plant 2008, 133, 725–735. [Google Scholar] [CrossRef] [PubMed]
- Bluhm, W.L.; Catalfomo, P. Poisonous Plants Encountered in Oregon; Oregon State University Extension Service: Corvallis, OR, USA, 1981. [Google Scholar]
- Kumar, A.; Singh, U.M.; Manohar, M.; Gaur, V.S. Calcium transport from source to sink: Understanding the mechanism (s) of acquisition, translocation, and accumulation for crop biofortification. Acta Physiol. Plant 2015, 37, 1722. [Google Scholar] [CrossRef]
- White, P.J. Long-distance transport in the xylem and phloem. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marshner, P., Ed.; Academic Press: Cambridge, MA, USA, 2012; pp. 49–70. [Google Scholar]
- Dichio, B.; Remorini, D.; Lang, S. Developmental changes in xylem functionality in kiwifruit fruit: Implications for fruit calcium accumulation. In Proceedings of the Acta Horticulturae, V International Symposium on Kiwifruit, Wuhan, China, 15–20 September 2002; International Society for Horticultural Science: Houston, TX, USA, 2003; pp. 191–195. [Google Scholar]
- Pliszka, B. Content and correlation of polyphenolic compounds, bioelements and antiradical activity in black elder berries (Sambucus nigra L.). J. Elem. 2020, 25, 595–605. [Google Scholar] [CrossRef]
- Wilkinson, S.R.; Welch, R.M.; Mayland, H.F.; Grunes, D.L. Magnesium in plants: Uptake, distribution, function, and utilization by man and animals. In Metal Ions in Biological Systems; Sigel, H., Sigel, A., Eds.; Marcel Dekker Inc.: New York, NY, USA; Basel, Switzerland, 1990; Volume 26, pp. 33–56. [Google Scholar]
- Karley, A.J.; White, P.J. Moving cationic minerals to edible tissues: Potassium, magnesium, calcium. Curr. Opin. Plant Biol. 2009, 12, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Rosanoff, A.; Dai, Q.; Shapses, S.A. Essential nutrient interactions: Does low or suboptimal magnesium status interact with vitamin D and/or calcium status? Adv. Nutr. 2016, 7, 25–43. [Google Scholar] [CrossRef] [Green Version]
- Veneklaas, E.J.; Lambers, H.; Bragg, J.; Finnegan, P.M.; Lovelock, C.E.; Plaxton, W.C.; Price, C.A.; Scheible, W.R.; Shane, M.W.; White, P.J. Opportunities for improving phosphorus-use efficiency in crop plants. New Phytol. 2012, 195, 306–320. [Google Scholar] [CrossRef] [PubMed]
- Page, V.; Feller, U. Heavy metals in crop plants: Transport and redistribution processes on the whole plant level. Agronomy 2015, 5, 447–463. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Sinha, S.; Saxena, R.; Pandey, K.; Bhatt, K. Translocation of metals and its effects in the tomato plants grown on various amendments of tannery waste: Evidence for involvement of antioxidants. Chemosphere 2004, 57, 91–99. [Google Scholar] [CrossRef]
- Nile, S.H.; Park, S.W. Edible berries: Bioactive components and their effect on human health. Nutrition 2014, 30, 134–144. [Google Scholar] [CrossRef]
- Page, V.; Weisskopf, L.; Feller, U. Heavy metals in white lupin: Uptake, root-to-shoot transfer and redistribution within the plant. New Phytol. 2006, 171, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos Utmazian, M.N.; Wenzel, W.W. Cadmium and zinc accumulation in willow and poplar species grown on polluted soils. J. Plant Nutr. Soil Sci. 2007, 170, 265–272. [Google Scholar] [CrossRef]
- Deng, H.; Ye, Z.H.; Wong, M.H. Accumulation of lead, zinc, copper and cadmium by 12 wetland plant species thriving in metal-contaminated sites in China. Environ. Pollut. 2004, 132, 29–40. [Google Scholar] [CrossRef]
- Yadav, A.K.; Abbassi, R.; Kumar, N.; Satya, S.; Sreekrishnan, T.R.; Mishra, B.K. The removal of heavy metals in wetland microcosms: Effects of bed depth, plant species, and metal mobility. Chem. Eng. J. 2012, 211, 501–507. [Google Scholar] [CrossRef]
- Soraya, R.K.; Takarina, N.D.; Soedjiarti, T. Metals accumulation (Cu, Zn and Pb) in mangrove-associated plants from Blanakan brackish water Pond, Subang district, West Java. In Proceedings of the AIP Conference Proceedings, 4th International Symposium on Current Progress in Mathematics and Sciences, Depok, Indonesia, 30–31 October 2018; Mart, T., Triyono, D., Anggraningrum, I.T., Eds.; AIP Publishing LLC: Melville, NY, USA, 2019; pp. 1–9. [Google Scholar]
- Dahmani-Muller, H.; van Oort, F.; Gélie, B.; Balabane, M. Strategies of heavy metal uptake by three plant species growing near a metal smelter. Environ. Pollut. 2000, 109, 231–238. [Google Scholar] [CrossRef]
- Gouthu, S.; Weginwar, R.; Arie, T.; Ambe, S.; Ozaki, T.; Enomoto, S.; Ambe, F.; Yamaguchi, I. Subcellular distribution and translocation of radionuclides in plants. Environ. Toxicol. Chem. 1999, 18, 2023–2027. [Google Scholar] [CrossRef]
- Dow, E.C.; Stanbury, J.B. Strontium and calcium metabolism in metabolic bone diseases. J. Clin. Investig. 1960, 39, 885–903. [Google Scholar] [CrossRef]
- Sasmaz, M.; Sasmaz, A. The accumulation of strontium by native plants grown on Gumuskoy mining soils. J. Geochem. Explor. 2017, 181, 236–242. [Google Scholar] [CrossRef]
- Watts, P.; Howe, P. Strontium and Strontium Compounds; World Health Organization: Geneva, Switzerland, 2010. [Google Scholar]
- Shen, R.; Ma, J.F. Distribution and mobility of aluminium in an Al-accumulating plant, Fagopyrum esculentum Moench. J. Exp. Bot. 2001, 52, 1683–1687. [Google Scholar] [PubMed]
- Wagatsuma, T. Characteristics of upward translocation of aluminum in plants. Soil Sci. Plant Nutr. 1984, 30, 345–358. [Google Scholar] [CrossRef] [Green Version]
- Exley, C. The toxicity of aluminium in humans. Morphologie 2016, 100, 51–55. [Google Scholar] [CrossRef]
- Stachiw, S.; Bicalho, B.; Grant-Weaver, I.; Noernberg, T.; Shotyk, W. Trace elements in berries collected near upgraders and open pit mines in the Athabasca Bituminous Sands Region (ABSR): Distinguishing atmospheric dust deposition from plant uptake. Sci. Total Environ. 2019, 670, 849–864. [Google Scholar] [CrossRef]
- Schaller, K. Praktikum zur Bodenkunde und Pflanzenernährung, 2nd ed.; Gesellschaft zur Förderung der Forschungsanstalt Geisenheim: Geisenhein, Germay, 1988. [Google Scholar]
- Team, R.C. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 8 March 2021).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2014, 67, 1–48. [Google Scholar]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest package: Tests in linear mixed effects models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Maechler, M.; Rousseeuw, P.J.; Struyf, A.; Hubert, M.; Hornik, K. Cluster: Cluster Analysis Basics and Extensions. R Package Version 2.1.0. 2012, 1, 56. [Google Scholar]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Clusters | Group 1 | Group 2 | Group 3 | Group 4 | |
|---|---|---|---|---|---|
| NI VIR LAC BB | JA × VIR (JA × NI) × BB C1 (JA × NI) × BB ((JA × NI) × BB) × (JA × (JA × NI)) JA × (JA × EB) JA × (((JA × NI) × NI) × MIQ) ((JA × NI) × BB) × (JA × MIQ) ((JA × NI) × SIB) × (JA × NI) JA × (JA × CER) ((CER × NI) × JA) × (CER × NI) (JA × RAC) × (CER × NI) ((JA × NI) × MIQ) × (CER × MIQ) ((JA × CER) × MIQ) × ((JA × NI) × CER) ((JA × NI) × SIB) × (JA × CER) No 3 | ((JA × NI) × NI) × ((JA × NI) × BB) ((JA × NI) × RAC) × ((JA × NI) × BB) (JA × (JA × MIQ)) × ((JA × NI) × BB) | JA × CER No 3 C1 ((JA × NI) × SIB) × CER | ||
| Mineral content | High | Ca (inflorescences) Ca (berries) Mg (inflorescences) Fe (inflorescences) Zn (inflorescences) Zn (berries) Sr (berries) Sr (inflorescences) Al (inflorescences) | K (inflorescences) K (berries) Mg (berries) Fe (berries) Mn (inflorescences) Mn (berries) Cu (inflorescences) Cu (berries) | K (inflorescences) K (berries) | Ca (inflorescences) Ca (berries) Zn (inflorescences) Zn (berries) Sr (inflorescences) Sr (berries) Al (berries) |
| Low | K (berries) K (inflorescences) Mn (berries) Al (berries) | Ca (inflorescences) Mg (berries) Fe (berries) Mn (berries) Mn (inflorescences) Cu (berries) Cu (inflorescences) Zn (berries) Sr (berries) Sr (inflorescences) | K (berries) K (inflorescences) Mg (inflorescences) Fe (inflorescences) Mn (berries) Mn (inflorescences) Al (inflorescences) | ||
| K | shoots | leaves | Berries | inflorescences | Cu | shoots | leaves | berries | Inflorescences |
| shoots | 1.00 | 0.61 *** | 0.44 ** | 0.39 ** | shoots | 1.00 | 0.31 * | 0.54 *** | 0.46 ** |
| leaves | 0.61 *** | 1.00 | 0.71 *** | 0.62 *** | leaves | 0.31 * | 1.00 | 0.40 ** | 0.40 ** |
| berries | 0.44 ** | 0.71 *** | 1.00 | 0.57 *** | berries | 0.54 *** | 0.40 ** | 1.00 | 0.60 *** |
| inflorescences | 0.39 ** | 0.62 *** | 0.57 *** | 1.00 | inflorescences | 0.46 ** | 0.40 ** | 0.60 *** | 1.00 |
| Ca | shoots | leaves | Berries | inflorescences | Mn | shoots | leaves | berries | inflorescences |
| shoots | 1.00 | 0.47 *** | 0.41 ** | 0.23 | shoots | 1.00 | 0.83 *** | 0.85 *** | 0.68 *** |
| leaves | 0.47 *** | 1.00 | 0.69 *** | 0.42 ** | leaves | 0.83 *** | 1.00 | 0.77 *** | 0.83 *** |
| berries | 0.41 ** | 0.69 *** | 1.00 | 0.50 *** | berries | 0.85 *** | 0.77 *** | 1.00 | 0.69 *** |
| inflorescences | 0.23 | 0.42 ** | 0.50 *** | 1.00 | inflorescences | 0.68 *** | 0.83 *** | 0.69 *** | 1.00 |
| Mg | shoots | leaves | Berries | inflorescences | Sr | shoots | leaves | berries | inflorescences |
| shoots | 1.00 | 0.54 *** | 0.25 | 0.45 ** | shoots | 1.00 | 0.76 *** | 0.77 *** | 0.71 *** |
| leaves | 0.54 *** | 1.00 | 0.41 ** | 0.62 *** | leaves | 0.76 *** | 1.00 | 0.78 *** | 0.73 *** |
| berries | 0.25 | 0.41 ** | 1.00 | 0.48 *** | berries | 0.77 *** | 0.78 *** | 1.00 | 0.78 *** |
| inflorescences | 0.45 ** | 0.62 *** | 0.48 *** | 1.00 | inflorescences | 0.71 *** | 0.73 *** | 0.78 *** | 1.00 |
| P | shoots | leaves | Berries | inflorescences | Zn | shoots | leaves | berries | inflorescences |
| shoots | 1.00 | 0.36 * | −0.22 | −0.14 | shoots | 1.00 | 0.42 ** | 0.51 *** | 0.45 ** |
| leaves | 0.36 * | 1.00 | 0.01 | 0.47 *** | leaves | 0.42 ** | 1.00 | 0.38 ** | 0.22 |
| berries | −0.22 | 0.01 | 1.00 | 0.38 ** | berries | 0.51 *** | 0.38 ** | 1.00 | 0.51 *** |
| inflorescences | −0.14 | 0.47 *** | 0.38 ** | 1.00 | inflorescences | 0.45 ** | 0.22 | 0.51 *** | 1.00 |
| Fe | shoots | leaves | Berries | inflorescences | Al | shoots | leaves | berries | inflorescences |
| shoots | 1.00 | 0.15 | 0.03 | 0.37 * | shoots | 1.00 | 0.15 | 0.00 | 0.22 |
| leaves | 0.15 | 1.00 | 0.11 | 0.47 *** | leaves | 0.15 | 1.00 | −0.04 | 0.30 * |
| berries | 0.03 | 0.11 | 1.00 | 0.27 | berries | 0.00 | −0.04 | 1.00 | −0.59 *** |
| inflorescences | 0.37 * | 0.47 *** | 0.27 | 1.00 | inflorescences | 0.22 | 0.30 * | −0.59 *** | 1.00 |
| Material | Abbreviation | No. |
|---|---|---|
| Species and other taxons | ||
| Sambucus nigra | NI | 2 |
| S. nigra var. viridis | VIR | 1 |
| S. nigra var. laciniata | LAC | 1 |
| S. nigra ‘Black Beauty’ | BB | 1 |
| Interspecific hybrids exhibiting combination of traits of S. nigra and S. javanica | ||
| S. javanica × S. nigra var. viridis | JA × VIR | 1 |
| (S. javanica × S. nigra) × S. nigra ‘Black Beauty’C1 a | (JA × NI) × BB C1 | 5 |
| (S. javanica × S. nigra) × S. nigra ‘Black Beauty’ b | (JA × NI) × BB | 2 |
| ((S. javanica × S. nigra) × S. nigra) × ((S. javanica × S. nigra) × S. nigra ‘Black Beauty’) | ((JA × NI) × NI) × ((JA × NI) × BB) | 6 |
| ((S. javanica × S. nigra) × S nigra ‘Black Beauty’) × (S. javanica × (S. javanica × S. nigra)) | ((JA × NI) × BB) × (JA × (JA × NI)) | 1 |
| Interspecific hybrids exhibiting similarity to S. ebulus | ||
| S. javanica × (S. javanica × S. ebulus) | JA × (JA × EB) | 1 |
| Interspecific hybrids exhibiting similarity to S. racemosa | ||
| ((S. javanica × S. nigra) × S. racemosa) × ((S. javanica × S. nigra) × S. nigra ‘Black Beauty’) | ((JA × NI) × RAC) × ((JA × NI) × BB) | 2 |
| S. javanica × (((S. javanica × S. nigra) × S. nigra) × S. racemosa − miquelii) | JA × (((JA × NI) × NI) × MIQ) | 3 |
| (S javanica × (S. javanica × S. racemosa − miquelii)) × ((S. javanica × S. nigra) × S. nigra ‘Black Beauty’) | (JA × (JA × MIQ)) × ((JA × NI) × BB) | 1 |
| ((S. javanica × S. nigra) × S. nigra ‘Black Beauty’) × (S. javanica × S. racemosa − miquelii) | ((JA × NI) × BB) × (JA × MIQ) | 1 |
| ((S. javanica × S. nigra) × S. racemosa subsp. sibirica) × (S. javanica × S. nigra) | ((JA × NI) × SIB) × (JA × NI) | 1 |
| Interspecific hybrids exhibiting similarity to S. cerulea | ||
| S. javanica × S. cerulea No 3 C1 | JA × CER No 3 C1 | 5 |
| S. javanica × (S. javanica × S. cerulea) | JA × (JA × CER) | 2 |
| ((S. cerulea × S. nigra) × S. javanica) × (S. cerulea × S. nigra) | ((CER × NI) × JA) × (CER × NI) | 1 |
| Interspecific hybrids exhibiting combination of S. cerulea and S. racemosa | ||
| (S. javanica × S. racemosa) × (S. cerulea × S. nigra) | (JA × RAC) × (CER × NI) | 1 |
| ((S. javanica × S. nigra) × S. racemosa − miquelii) × (S. cerulea × S. racemosa − miquelii) | ((JA × NI) × MIQ) × (CER × MIQ) | 1 |
| ((S. javanica × S. cerulea) × S. racemosa − miquelii) × ((S. javanica × S. nigra) × S. cerulea) | ((JA × CER) × MIQ) × ((JA × NI) × CER) | 2 |
| ((S. javanica × S. nigra) × S. racemosa subsp. sibirica) × S. cerulea | ((JA × NI) × SIB) × CER | 1 |
| ((S. javanica × S. nigra) × S. racemosa subsp. sibirica) × (S. javanica × S. cerulea) No 3 | ((JA × NI) × SIB) × (JA × CER) No 3 | 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Imenšek, N.; Sem, V.; Kolar, M.; Ivančič, A.; Kristl, J. The Distribution of Minerals in Crucial Plant Parts of Various Elderberry (Sambucus spp.) Interspecific Hybrids. Plants 2021, 10, 653. https://doi.org/10.3390/plants10040653
Imenšek N, Sem V, Kolar M, Ivančič A, Kristl J. The Distribution of Minerals in Crucial Plant Parts of Various Elderberry (Sambucus spp.) Interspecific Hybrids. Plants. 2021; 10(4):653. https://doi.org/10.3390/plants10040653
Chicago/Turabian StyleImenšek, Nataša, Vilma Sem, Mitja Kolar, Anton Ivančič, and Janja Kristl. 2021. "The Distribution of Minerals in Crucial Plant Parts of Various Elderberry (Sambucus spp.) Interspecific Hybrids" Plants 10, no. 4: 653. https://doi.org/10.3390/plants10040653
APA StyleImenšek, N., Sem, V., Kolar, M., Ivančič, A., & Kristl, J. (2021). The Distribution of Minerals in Crucial Plant Parts of Various Elderberry (Sambucus spp.) Interspecific Hybrids. Plants, 10(4), 653. https://doi.org/10.3390/plants10040653