
Citrus Cell Suspension Culture Establishment, Maintenance, Efficient Transformation and Regeneration to Complete Transgenic Plant

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
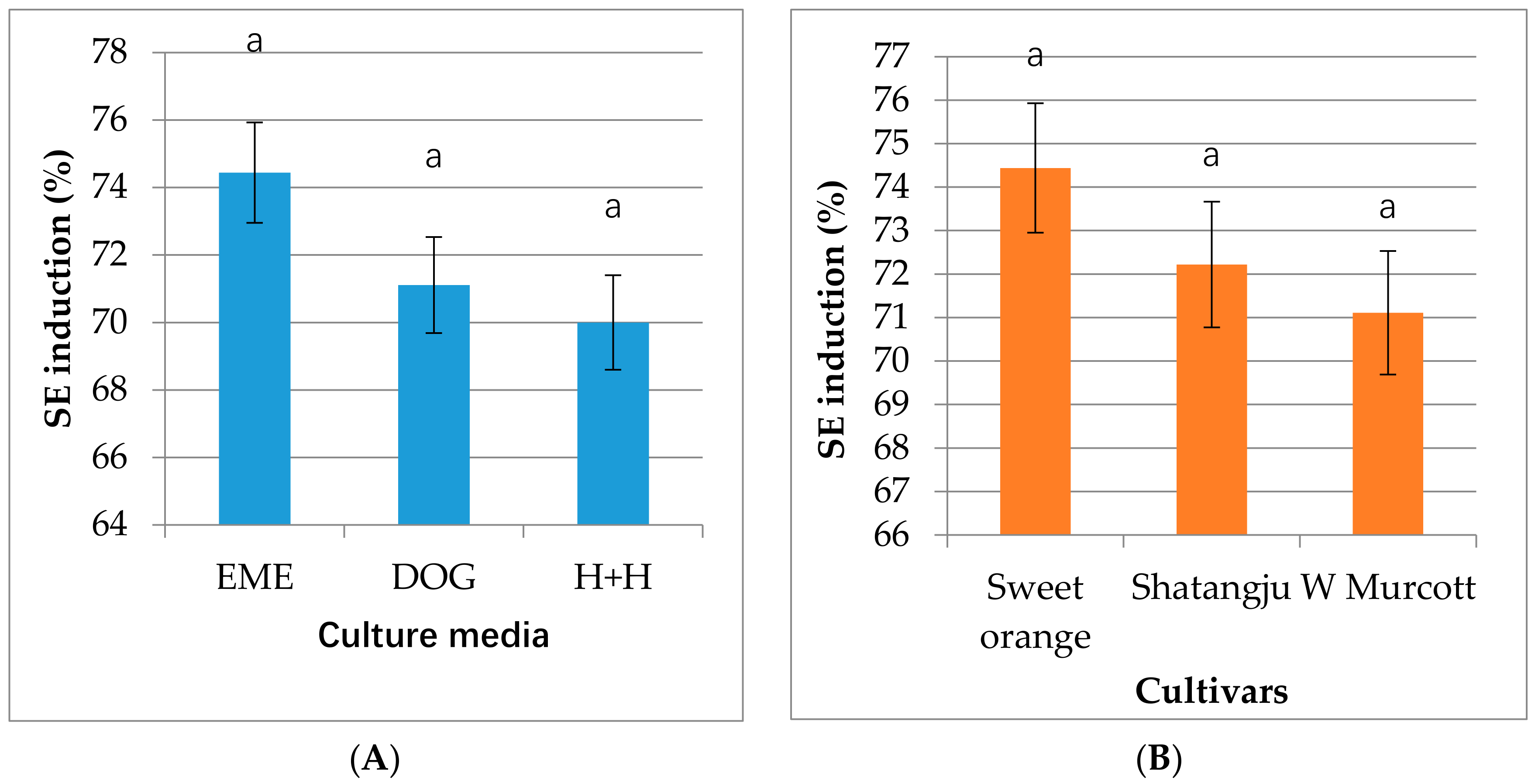
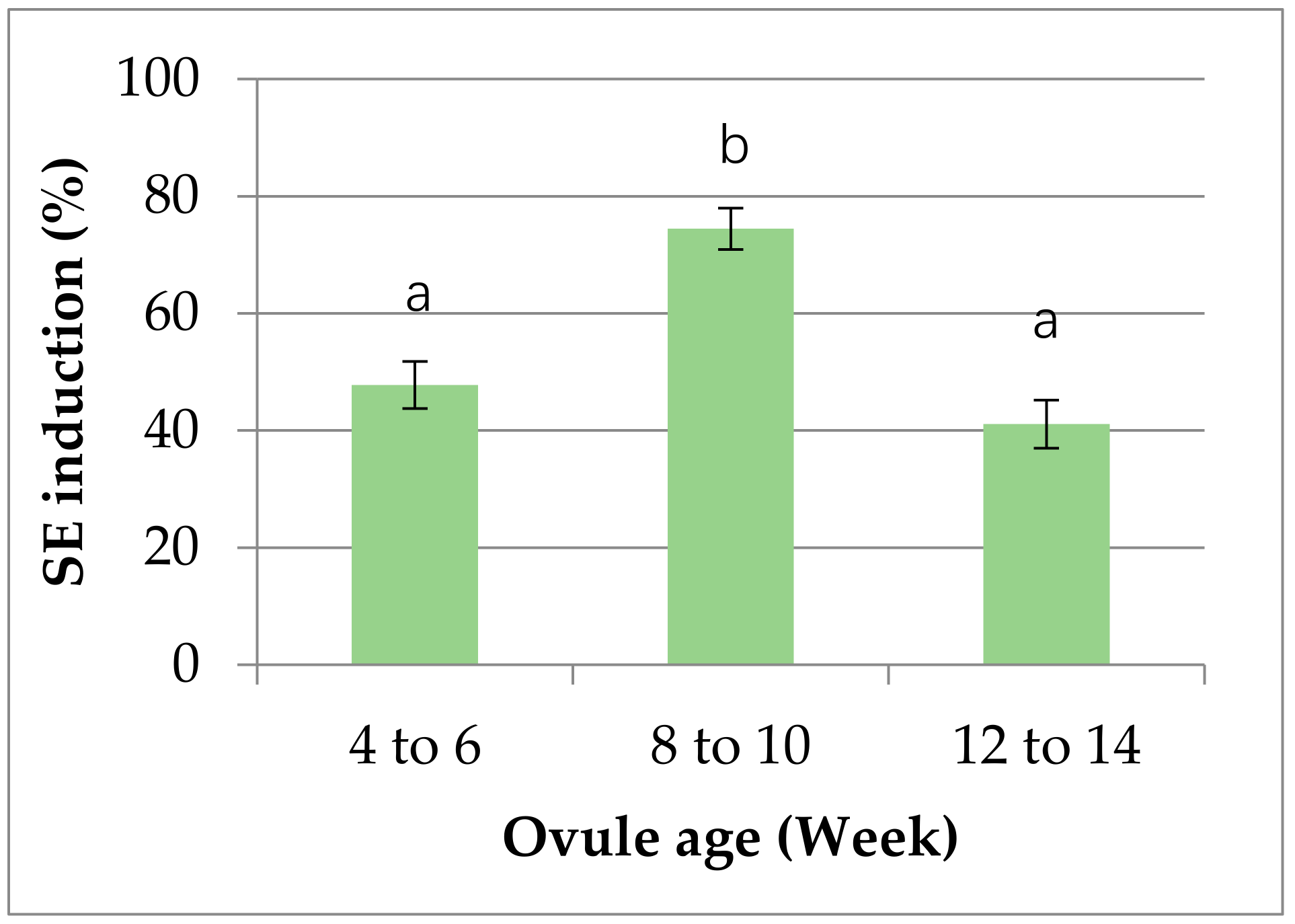
2.1. Embryogenic Callus Induction
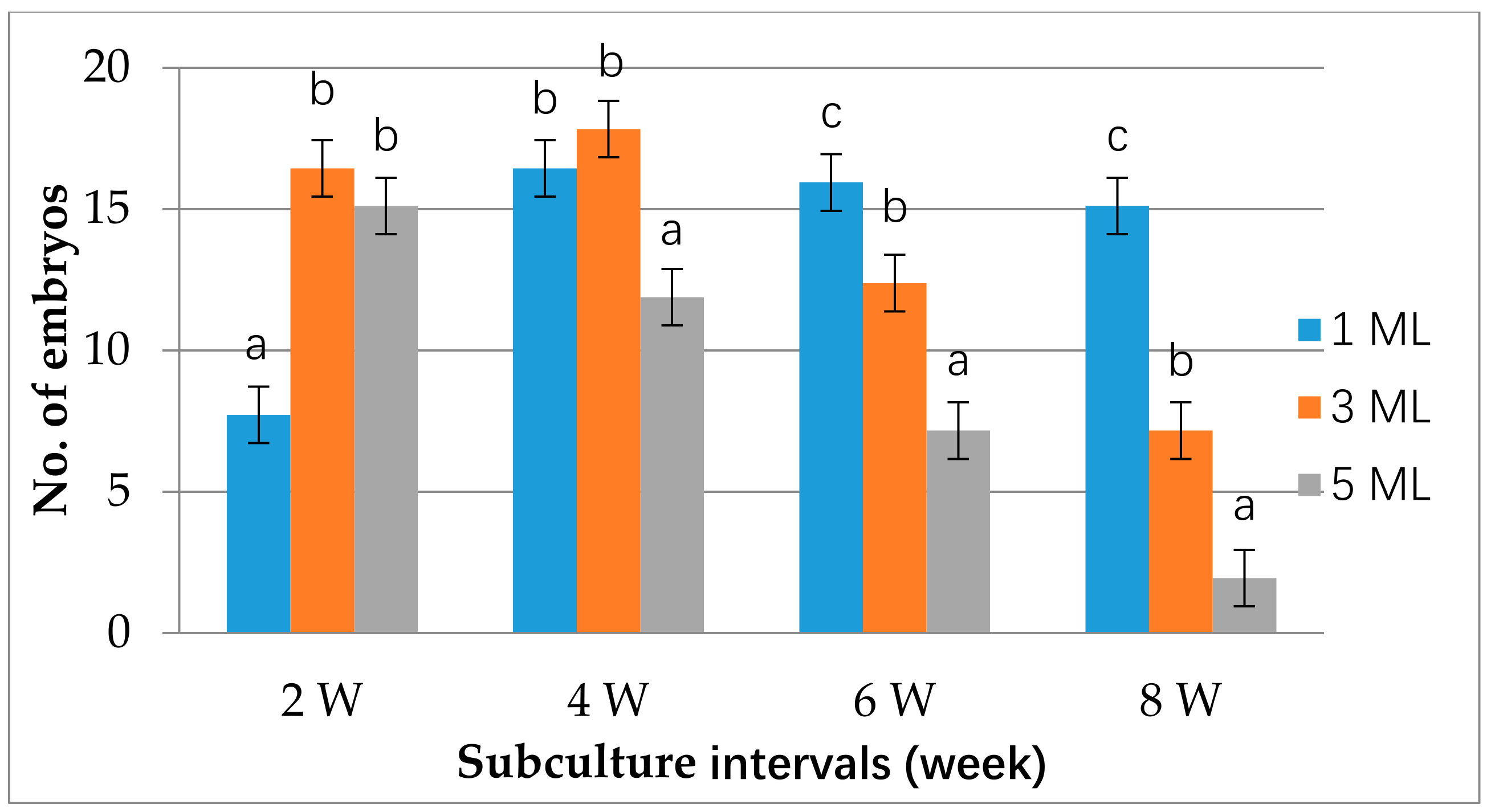
2.2. Suspension Cell Culture and Plant Regeneration
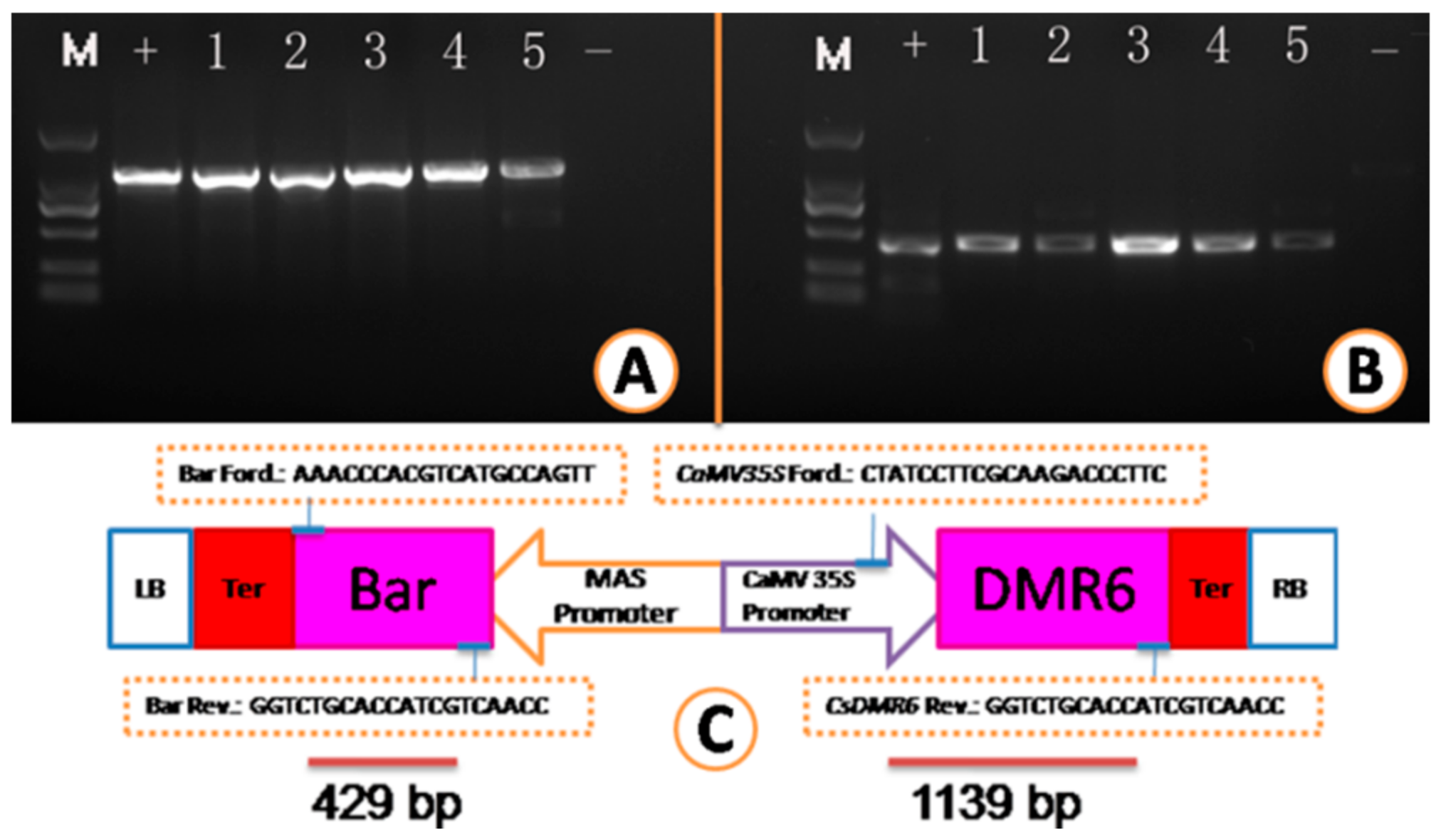
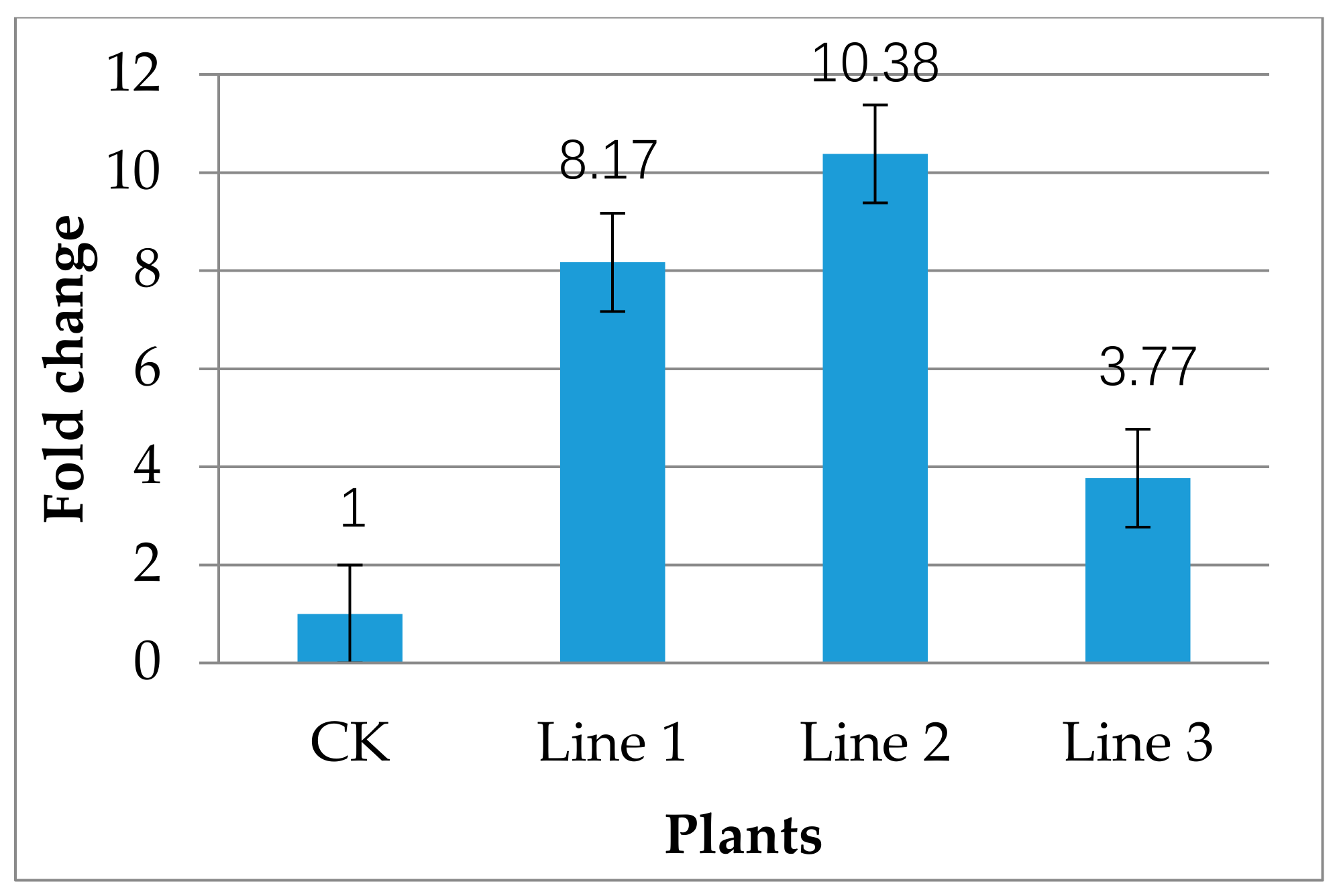
2.3. Transgenic Plant Recovery and Molecular Analysis of Transgenic Plants
3. Materials and Methods
3.1. Plant Materials and Embryonic Callus Formation
3.2. Establishment of Cell Suspension Culture and Plant Organogenesis
3.3. Vector Construction, Agrobacterium Transformation and Plant Regeneration
3.4. PCR and qPCR Analysis
3.5. Data Recording and Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhong, G.; Nicolosi, E. Citrus origin, diffusion, and economic importance. In The Citrus Genome. Compendium of Plant Genomes; Gentile, A., La Malfa, S., Deng, Z., Eds.; Springer: Cham, Switzerland, 2020; pp. 5–21. [Google Scholar]
- Martínez-Blay, V.; Taberner, V.; Pérez-Gago, M.B.; Palou, L. Control of major citrus postharvest diseases by sulfur-containing food additives. Int. J. Food Microbiol. 2020, 330, 108713. [Google Scholar] [CrossRef]
- Moniruzzaman, M.; Zhong, Y.; Yan, H.; Yuanda, L.; Jiang, B.; Zhong, G. Exploration of Susceptible Genes with Clustered Regularly Interspaced Short Palindromic Repeats–Tissue-Specific Knockout (CRISPR-TSKO) to Enhance Host Resistance. Crit. Rev. Plant Sci. 2020, 39, 387–417. [Google Scholar] [CrossRef]
- Pang, Z.; Zhang, L.; Coaker, G.L.; Ma, W.; He, S.-Y.; Wang, N. Citrus CsACD2 Is a Target of CandidatusLiberibacterAsiaticus in Huanglongbing Disease. Plant Physiol. 2020, 184, 792–805. [Google Scholar] [CrossRef]
- Dala-Paula, B.M.; Plotto, A.; Bai, J.; Manthey, J.A.; Baldwin, E.A.; Ferrarezi, R.S.; Gloria, M.B.A. Effect of Huanglongbing or Greening Disease on Orange Juice Quality, a Review. Front. Plant Sci. 2019, 9, 1976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soost, R.; Cameron, J. Tree and fruit characters of Citrus triploids from tetraploid by diploid crosses. Hilgardia 1969, 39, 569–579. [Google Scholar] [CrossRef] [Green Version]
- Salonia, F.; Ciacciulli, A.; Poles, L.; Pappalardo, H.D.; La Malfa, S.; Licciardello, C. New Plant Breeding Techniques in Citrus for the Improvement of Important Agronomic Traits. A Review. Front. Plant Sci. 2020, 11, 1234. [Google Scholar] [CrossRef]
- Qaim, M.; Kouser, S. Genetically modified crops and food security. PLoS ONE 2013, 8, e64879. [Google Scholar] [CrossRef] [Green Version]
- Gasser, C.S.; Fraley, R.T. Genetically engineering plants for crop improvement. Science 1989, 244, 1293–1299. [Google Scholar] [CrossRef] [Green Version]
- Song, G.; Jia, M.; Chen, K.; Kong, X.; Khattak, B.; Xie, C.; Li, A.; Mao, L. Crispr/cas9: A powerful tool for crop genome editing. Crop. J. 2016, 4, 75–82. [Google Scholar] [CrossRef] [Green Version]
- Talon, M.; Gmitter, F.G., Jr. Citrus genomics. Int. J. Plant Genom. 2008, 2008, 528361. [Google Scholar] [CrossRef] [PubMed]
- Gambino, G.; Gribaudo, I. Genetic transformation of fruit trees: Current status and remaining challenges. Transgenic Res. 2012, 21, 1163–1181. [Google Scholar] [CrossRef]
- Van Nocker, S.; Gardiner, S.E. Breeding better cultivars, faster: Applications of new technologies for the rapid deployment of superior horticultural tree crops. Hortic. Res. 2014, 1, 14022. [Google Scholar] [CrossRef] [Green Version]
- Peña, L.; Pérez, R.M.; Cervera, M.; Juárez, J.A.; Navarro, L. Early events in agrobacterium-mediated genetic transformation of Citrus explants. Ann. Bot. 2004, 94, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Dutt, M.; Grosser, J.W. Evaluation of parameters affecting Agrobacterium-mediated transformation of Citrus. Plant Cell Tissue Organ Cult. 2009, 98, 331–340. [Google Scholar] [CrossRef]
- De Almeida, W.A.B.; Filho, F.D.A.A.M.; Mendes, B.M.J.; Pavan, A.; Rodriguez, A.P.M. Agrobacterium-mediated transformation of Citrus sinensis and Citrus limonia epicotyl segments. Sci. Agric. 2003, 60, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Molinari, H.B.C.; Bespalhok, J.C.; Kobayashi, A.K.; Pereira, L.F.P.; Vieira, L.G.E. Agrobacterium tumefaciens-mediated transformation of Swingle citrumelo (Citrus paradisi macf.×Poncirus trifoliata l. Raf.) using thin epicotyl sections. Sci. Hortic. 2004, 99, 379–385. [Google Scholar] [CrossRef]
- Ghorbel, R.; Navarro, L.; Duran-Vila, N. Morphogenesis and regeneration of whole plants of grapefruit (Citrus paradisi), sour orange (C. Aurantium) and alemow (C. Macrophylla). J. Hortic. Sci. Biotechnol. 1998, 73, 323–327. [Google Scholar] [CrossRef]
- Moore, G.A.; Jacono, C.C.; Neidigh, J.L.; Lawrence, S.D.; Cline, K. Agrobacterium-mediated transformation of citrus stem segments and regeneration of transgenic plants. Plant Cell Rep. 1992, 11, 238–242. [Google Scholar] [CrossRef]
- Febres, V.; Fisher, L.; Khalaf, A.; Moore, G.A. Citrus transformation: Challenges and prospects. In Genetic Transformation; Alvarez, M., Ed.; IntechOpen: London, UK, 2011. [Google Scholar]
- Cervera, M.; López, M.M.; Navarro, L.; Peña, L. Virulence and supervirulence of Agrobacterium tumefaciensin woody fruit plants. Physiol. Mol. Plant Pathol. 1998, 52, 67–78. [Google Scholar] [CrossRef]
- Dutt, M.; Erpen, L.; Grosser, J.W. Genetic transformation of the ‘W Murcott’ tangor: Comparison between different techniques. Sci. Hortic. 2018, 242, 90–94. [Google Scholar] [CrossRef]
- Almeida, W.A.B.; Mourão Filho, F.A.A.; Pino, L.E.; Boscariol, R.L.; Rodriguez, A.P.M.; Mendes, B.M.J. Genetic transformation and plant recovery from mature tissues of Citrus sinensis l. Osbeck. Plant Sci. 2003, 164, 203–211. [Google Scholar] [CrossRef]
- Cervera, M.; Navarro, A.; Navarro, L.; Peña, L. Production of transgenic adult plants from clementine mandarin by enhancing cell competence for transformation and regeneration. Tree Physiol. 2008, 28, 55–66. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, A.K.; Bespalhok, J.C.; Pereira, L.F.P.; Vieira, L.G.E. Plant regeneration of sweet orange (Citrus sinensis) from thin sections of mature stem segments. Plant Cell Tissue Organ Cult. 2003, 74, 99–102. [Google Scholar] [CrossRef]
- Aderkas, P.V.; Bonga, J. Influencing micropropagation and somatic embryogenesis in mature trees by manipulation of phase change, stress and culture environment. Tree Physiol. 2000, 20, 921–928. [Google Scholar] [CrossRef] [Green Version]
- Henry, R. Molecular and biochemical characterization of somaclonal variation. In Somaclonal Variation and Induced Mutations in Crop Improvement. Current Plant Science and Biotechnology in Agriculture; Jain, S.M., Brar, D.S., Ahloowalia, B.S., Eds.; Springer: Dordrecht, The Netherlands, 1998; Volume 32. [Google Scholar]
- Omar, A.A.; Dutt, M.; Gmitter, F.G.; Grosser, J.W. Somatic embryogenesis: Still a relevant technique in citrus improvement. In In Vitro Embryogenesis in Higher Plants; Humana Press: New York, NY, USA, 2016; Volume 1359, pp. 289–327. [Google Scholar]
- Dutt, M.; Grosser, J.W. An embryogenic suspension cell culture system for Agrobacterium-mediated transformation of citrus. Plant Cell Rep. 2010, 29, 1251–1260. [Google Scholar] [CrossRef]
- Haccius, B. Question of unicellular origin of non-zygotic embryos in callus cultures. Phytomorphology 1978, 28, 74–81. [Google Scholar]
- Toonen, M.A.J.; Hendriks, T.; Schmidt, E.D.L.; Verhoeven, H.A.; van Kammen, A.; de Vries, S.C. Description of somatic-embryo-forming single cells in carrot suspension cultures employing video cell tracking. Planta 1994, 194, 565–572. [Google Scholar] [CrossRef]
- Lee, K.S.; Zapata-Arias, F.J.; Brunner, H.; Afza, R. Histology of somatic embryo initiation and organogenesis from rhizome explants of Musa spp. Plant Cell Tissue Organ Cult. 1997, 51, 1–8. [Google Scholar] [CrossRef]
- Chan, J.L.; Saénz, L.; Talavera, C.; Hornung, R.; Robert, M.; Oropeza, C. Regeneration of Coconut (Cocos nucifera L.) from plumule explants through somatic embryogenesis. Plant Cell Rep. 1998, 17, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Rugkhla, A.; Jones, M.G.K. Somatic embryogenesis and plantlet formation in Santalum album and S. spicatum. J. Exp. Bot. 1998, 49, 563–571. [Google Scholar] [CrossRef]
- Nonohay, J.S.; Mariath, J.E.A.; Winge, H. Histological analysis of somatic embryogenesis in brazilian cultivars of barley, Hordeum vulgare vulgare, poaceae. Plant Cell Rep. 1999, 18, 929–934. [Google Scholar] [CrossRef]
- Roussos, P. Orange (Citrus sinensis (l.) Osbeck). In Nutritional Composition of Fruit Cultivars; Academic Press: Cambridge, UK, 2016; pp. 469–496. [Google Scholar]
- Cao, J.; Kang, C.; Chen, Y.; Karim, N.; Wang, Y.; Sun, C. Physiochemical changes in Citrus reticulata cv. Shatangju fruit during vesicle collapse. Postharvest Biol. Technol. 2020, 165, 111180. [Google Scholar] [CrossRef]
- Yan, H.; Zhou, B.; Jiang, B.; Lv, Y.; Moniruzzaman, M.; Zhong, G.; Zhong, Y. Comparative analysis of bacterial and fungal endophytes responses to Candidatus liberibacterasiaticus infection in leaf midribs of Citrus reticulata cv. Shatangju. Physiol. Mol. Plant. Pathol. 2021, 113, 101590. [Google Scholar] [CrossRef]
- Omar, A.A.; Murata, M.M.; El-Shamy, H.A.; Graham, J.H.; Grosser, J.W. Enhanced resistance to citrus canker in transgenic mandarin expressing xa21 from rice. Transgenic Res. 2018, 27, 179–191. [Google Scholar] [CrossRef]
- Grosser, J.W.; Calović, M.; Louzada, E.S. Protoplast fusion technology—Somatic hybridization and cybridization. In Plant Cell Culture; Davey, M.R., Anthony, P., Eds.; John Wiley & Sons, Ltd.: New York, NY, USA, 2010; pp. 175–198. [Google Scholar]
- Kayim, M.; Koc, N.K. The effects of some carbohydrates on growth and somatic embryogenesis in citrus callus culture. Sci. Hortic. 2006, 109, 29–34. [Google Scholar] [CrossRef]
- Rangan, T.S.; Murashige, T.; Bitters, W.P. In vitro initiation of nucellar embryos in monoembryonic Citrus. HortScience 1968, 3, 226–227. [Google Scholar]
- Bitters, W.P.; Murashigi, T.; Rangan, T.S.; Nauer, E.M. Investigations on established virusfree plants through tissue culture. Calif. Citrus Nurs. Soc. 1970, 9, 27–30. [Google Scholar]
- Button, J.; Bornman, C.H. Development of nucellar plants from unpollinated and unfertilized ovules of the washington navel orange in vitro. J. S. Afr. Bot. 1971, 37, 127–134. [Google Scholar]
- Gmitter, F.G.; Moore, G.A. Plant regeneration from undeveloped ovules and embryogenic calli of Citrus: Embryo production, germination, and plant survival. Plant Cell Tissue Organ Cult. 1986, 6, 139–147. [Google Scholar] [CrossRef]
- Carimi, F.; Tortorici, M.C.; De Pasquale, F.; Crescimanno, F.G. Somatic embryogenesis and plant regeneration from undeveloped ovules and stigma/style explants of sweet orange navel group [Citrus sinensis (l.) osb.]. Plant Cell Tissue Organ Cult. 1998, 54, 183–189. [Google Scholar] [CrossRef]
- Litz, R.E.; Moore, G.A.; Srinivasan, C. In vitro systems for propagation and improvement of tropical fruits and palms. Hortic. Rev. 1985, 7, 157–200. [Google Scholar]
- Nito, N.; Iwamasa, M. In vitro plantlet formation from juice vesicle callus of satsuma (Citrus unshiu Marc.). Plant Cell Tissue Organ Cult. 1990, 20, 137–140. [Google Scholar] [CrossRef]
- Hidaka, T.; Yamada, Y.; Shichijo, T. Plantlet formation by anther culture of Citrus aurantium L. Ikushugaku Zasshi 1982, 32, 247–252. [Google Scholar] [CrossRef] [Green Version]
- Carimi, F.; De Pasquale, F.; Crescimanno, F.G. Somatic embryogenesis from styles of lemon (Citrus limon). Plant Cell Tissue Organ Cult. 1994, 37, 209–211. [Google Scholar] [CrossRef]
- Gill, M.I.S.; Singh, Z.; Dhillon, B.S.; Gosal, S.S. Somatic embryogenesis and plantlet regeneration in mandarin (Citrus reticulata blanco). Sci. Hortic. 1995, 63, 167–174. [Google Scholar] [CrossRef]
- Navarro, L.; Ortíz, J.; Juárez, J. Aberrant citrus plants obtained by somatic embryogenesis of nucelli cultured in vitro. HortScience 1985, 20, 214–215. [Google Scholar]
- Grosser, J.W.; Gmitter, F.G. Protoplast fusion for production of tetraploids and triploids: Applications for scion and rootstock breeding in citrus. Plant Cell Tissue Organ Cult. 2011, 104, 343–357. [Google Scholar] [CrossRef]
- Moscatiello, R.; Baldan, B.; Navazio, L. Plant cell suspension cultures. In Plant Mineral Nutrients; Maathuis, F., Ed.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2013; Volume 953, pp. 77–93. [Google Scholar]
- Mathur, J.; Koncz, C. Establishment and maintenance of cell suspension cultures. In Arabidopsis Protocols; Martinez-Zapater, J.M., Salinas, J., Eds.; Methods in Molecular Biology; Humana Press: New York, NY, USA, 1988; Volume 82. [Google Scholar]
- Guo, W.; Li, D.; Duan, Y. Citrus transgenics: Current status and prospects. Transgenic Plant J. 2007, 1, 202–209. [Google Scholar]
- Ghaderi, I.; Sohani, M.M.; Mahmoudi, A. Efficient genetic transformation of sour orange, Citrus aurantium l. Using agrobacterium tumefaciens containing the coat protein gene of Citrus tristeza virus. Plant Gene 2018, 14, 7–11. [Google Scholar] [CrossRef]
- Zhang, D.-M.; Zhong, Y.; Chang, X.-J.; Hu, M.-L.; Cheng, C.-Z.; Zhang, Y.-Y. A simple and efficient in planta transformation method for pommelo (Citrus maxima) using Agrobacterium tumefaciens. Sci. Hortic. 2017, 214, 174–179. [Google Scholar] [CrossRef]
- Donmez, D.; Simsek, O.; Izgu, T.; Kacar, Y.A.; Mendi, Y.Y. Genetic transformation in citrus. Sci. World J. 2013, 2013, 491207. [Google Scholar] [CrossRef]
- Kong, Q.; Wu, M.; Huan, Y.; Zhang, L.; Liu, H.; Bou, G.; Luo, Y.; Mu, Y.; Liu, Z. Transgene expression is associated with copy number and cytomegalovirus promoter methylation in transgenic pigs. PLoS ONE 2009, 4, e6679. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.; Hwang, G.; Abdul Razak, S.; Sohm, F.; Maclean, N. Copy number related transgene expression and mosaic somatic expression in hemizygous and homozygous transgenic tilapia (Oreochromis niloticus). Transgenic Res. 2001, 9, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Hobbs, S.L.A.; Warkentin, T.D.; DeLong, C.M.O. Transgene copy number can be positively or negatively associated with transgene expression. Plant Mol. Biol. 1993, 21, 17–26. [Google Scholar] [CrossRef]
- Collier, R.; Dasgupta, K.; Xing, Y.P.; Hernandez, B.T.; Shao, M.; Rohozinski, D.; Kovak, E.; Lin, J.; de Oliveira, M.L.P.; Stover, E.; et al. Accurate measurement of transgene copy number in crop plants using droplet digital PCR. Plant J. 2017, 90, 1014–1025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prols, F.; Meyer, P. The methylation patterns of chromosomal integration regions influence gene activity of transferred DNA in petunia hybrida. Plant J. 1992, 2, 465–475. [Google Scholar] [PubMed]
- Ottaviani, M.-P.; Smits, T.; Cate, C.H.H.T. Differential methylation and expression of the β-glucuronidase and neomycin phosphotransferase genes in transgenic plants of potato cv. Bintje. Plant Sci. 1993, 88, 73–81. [Google Scholar] [CrossRef]
- Murashige, T.; Tucker, D.P. Growth factor requirements of citrus tissue culture. Proc. 1st Intl. Citrus Symp. 1969, 3, 1155–1161. [Google Scholar]
- Zeilmaker, T.; Ludwig, N.R.; Elberse, J.; Seidl, M.F.; Berke, L.; Van Doorn, A.; Schuurink, R.C.; Snel, B.; Ackerveken, G.V.D. DOWNY MILDEW RESISTANT 6 and DMR6-LIKE OXYGENASE 1 are partially redundant but distinct suppressors of immunity in Arabidopsis. Plant J. 2014, 81, 210–222. [Google Scholar] [CrossRef]
- De Toledo Thomazella, D.P.; Brail, Q.; Thomazella, D.; Dahlbeck, D.; Staskawicz, B. Crispr-cas9 mediated mutagenesis of a DMR6 ortholog in tomato confers broad-spectrum disease resistance. bioRxiv 2016, 064824. [Google Scholar] [CrossRef] [Green Version]
- Van Damme, M.; Huibers, R.P.; Elberse, J.; Ackerveken, G.V.D. Arabidopsis DMR6 encodes a putative 2OG-Fe(II) oxygenase that is defense-associated but required for susceptibility to downy mildew. Plant J. 2008, 54, 785–793. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.C.D.L.; Shimo, H.M.; De Felício, R.; Mercaldi, G.F.; Rocco, S.A.; Benedetti, C.E. Structure-function relationship of a citrus salicylate methylesterase and role of salicylic acid in citrus canker resistance. Sci. Rep. 2019, 9, 3901. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivars | Embryos on Non-Selection Media | Embryos on BASTA (R) Selection Media | Transformation Percentage (%) |
|---|---|---|---|
| “Sweet orange” | 51.94 ± 1.94 | 17.83 ± 0.39 | 35.09 ± 1.43 |
| “Shatangju” | 53.06 ±1.78 | 16.88 ± 0.63 | 32.42 ± 1.56 |
| “W. Murcott” | 53.66 ± 1.82 | 17.11 ± 0.8 | 32.38 ± 1.68 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moniruzzaman, M.; Zhong, Y.; Huang, Z.; Yan, H.; Yuanda, L.; Jiang, B.; Zhong, G. Citrus Cell Suspension Culture Establishment, Maintenance, Efficient Transformation and Regeneration to Complete Transgenic Plant. Plants 2021, 10, 664. https://doi.org/10.3390/plants10040664
Moniruzzaman M, Zhong Y, Huang Z, Yan H, Yuanda L, Jiang B, Zhong G. Citrus Cell Suspension Culture Establishment, Maintenance, Efficient Transformation and Regeneration to Complete Transgenic Plant. Plants. 2021; 10(4):664. https://doi.org/10.3390/plants10040664
Chicago/Turabian StyleMoniruzzaman, M., Yun Zhong, Zhifeng Huang, Huaxue Yan, Lv Yuanda, Bo Jiang, and Guangyan Zhong. 2021. "Citrus Cell Suspension Culture Establishment, Maintenance, Efficient Transformation and Regeneration to Complete Transgenic Plant" Plants 10, no. 4: 664. https://doi.org/10.3390/plants10040664
APA StyleMoniruzzaman, M., Zhong, Y., Huang, Z., Yan, H., Yuanda, L., Jiang, B., & Zhong, G. (2021). Citrus Cell Suspension Culture Establishment, Maintenance, Efficient Transformation and Regeneration to Complete Transgenic Plant. Plants, 10(4), 664. https://doi.org/10.3390/plants10040664