
MYC2 Transcription Factors TwMYC2a and TwMYC2b Negatively Regulate Triptolide Biosynthesis in Tripterygium wilfordii Hairy Roots

 and
and 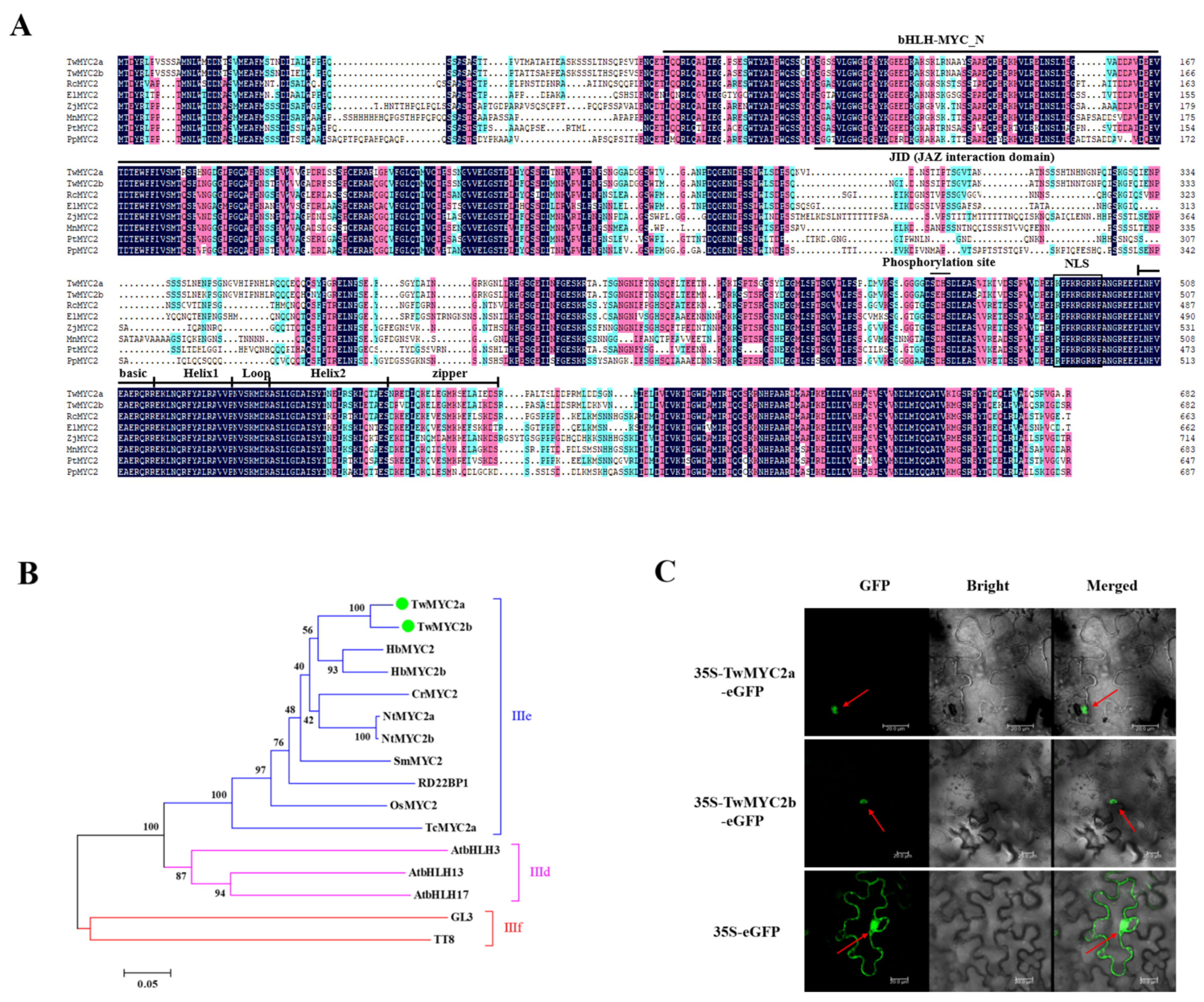{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Isolation and Molecular Characterization of TwMYC2a and TwMYC2b
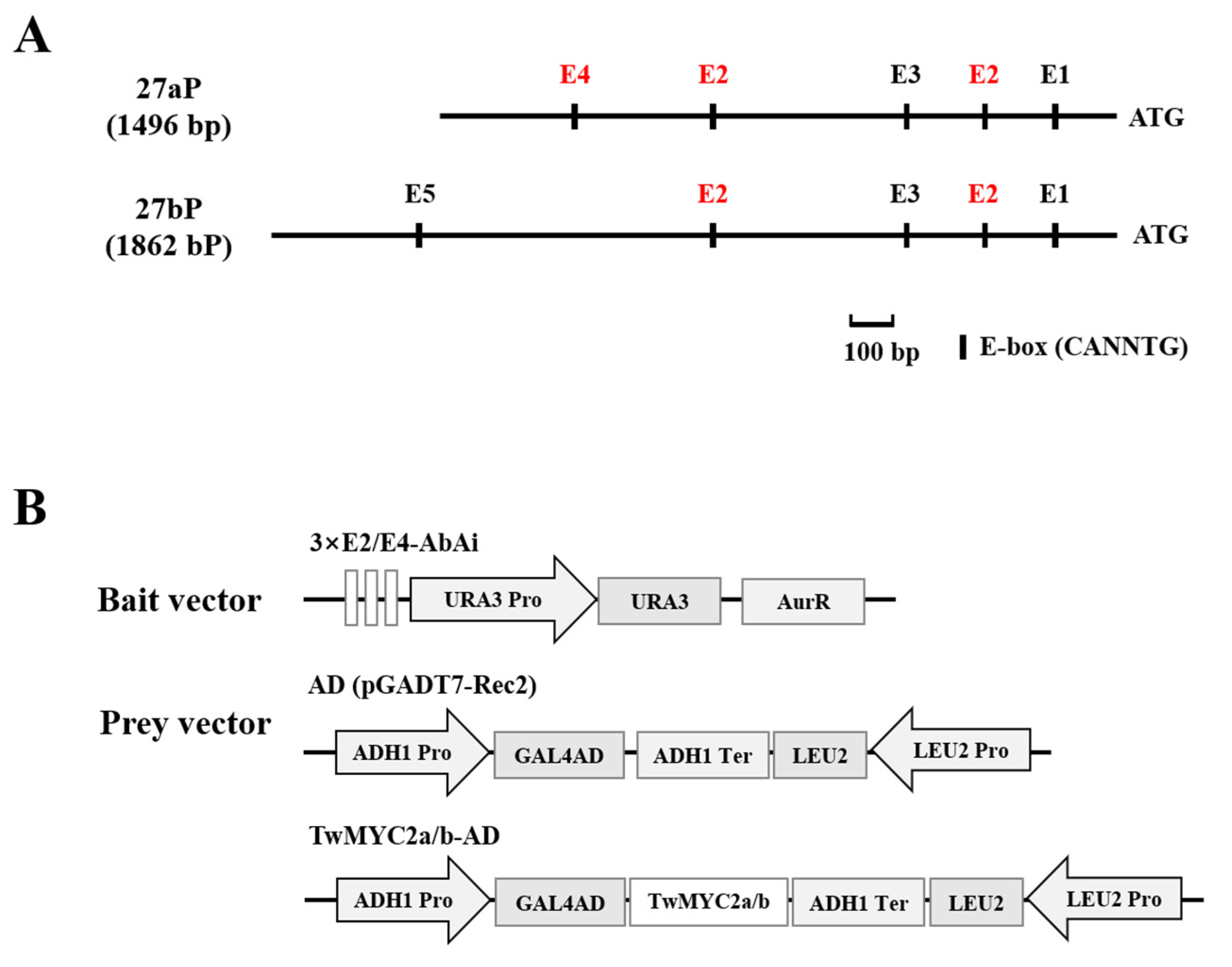
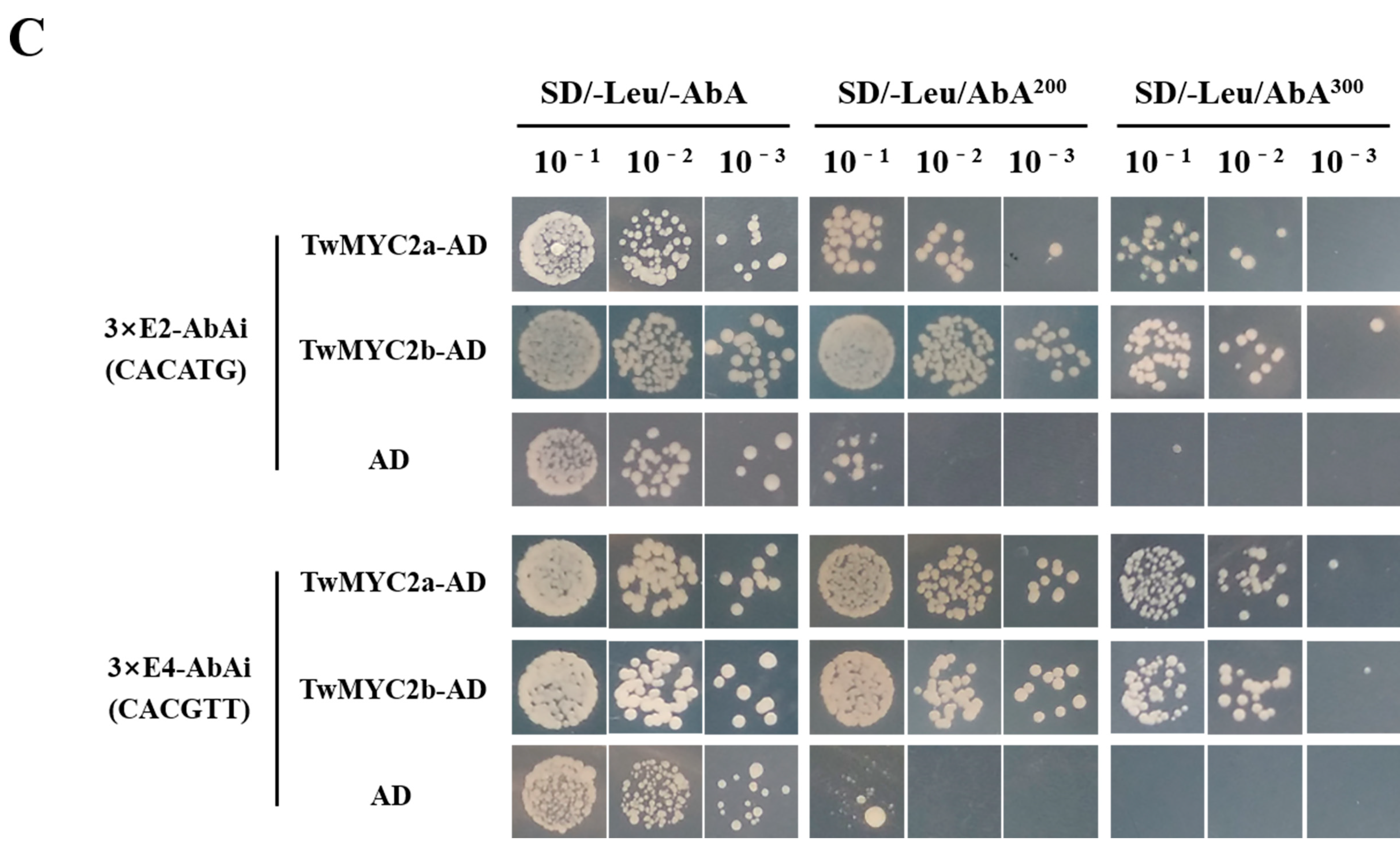
2.2. TwMYC2a and TwMYC2b Specifically Bind the E-Box and T/G-Box Motifs within the TwTPS27a and TwTPS27b Promoters
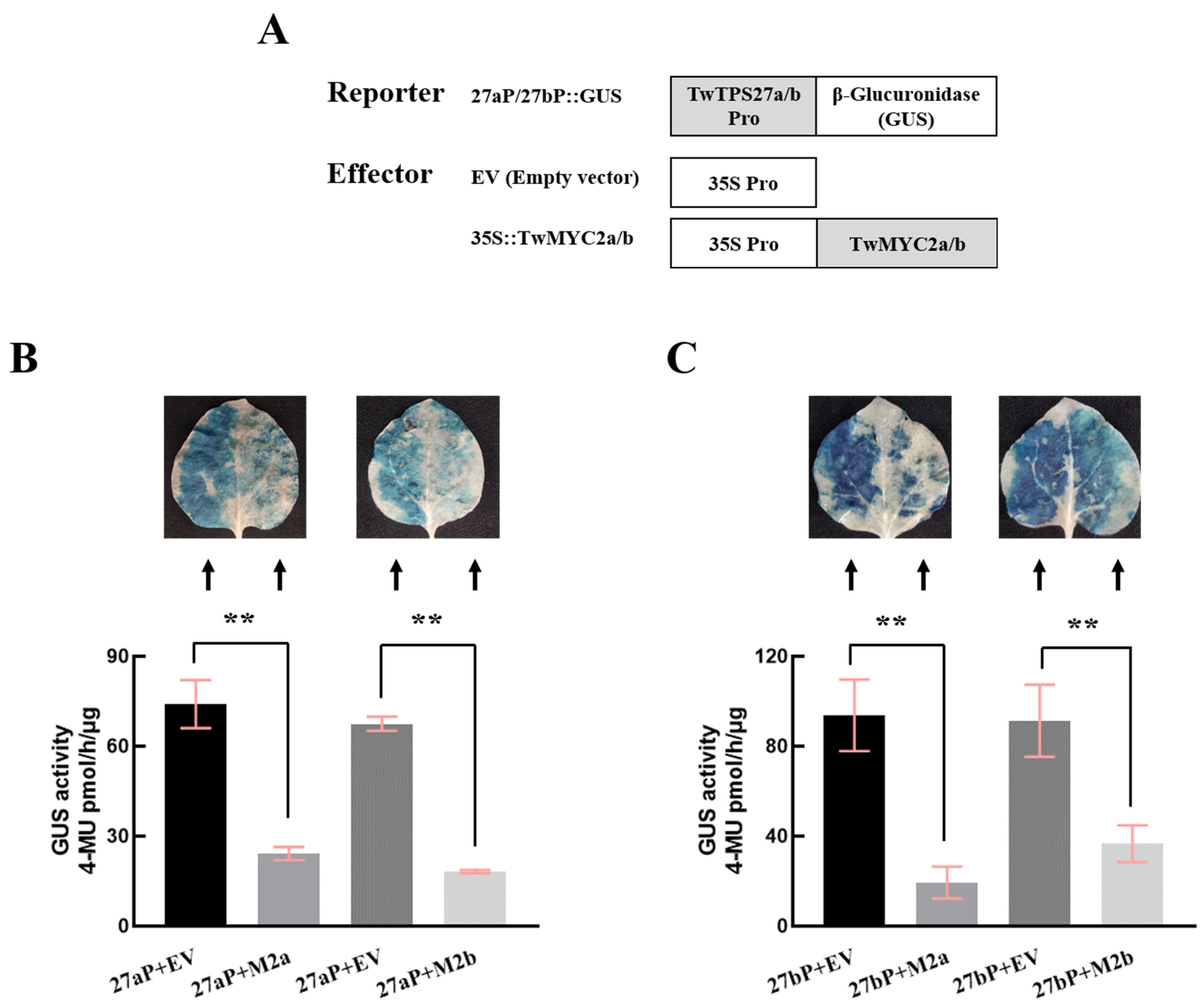
2.3. TwMYC2a and TwMYC2b Could Repress the TwTPS27a and TwTPS27b Promoters
2.4. Relative Expression of TwMYC2a/b and Key Pathway Genes in the RNA Interference (RNAi) Hairy Roots of T. wilfordii
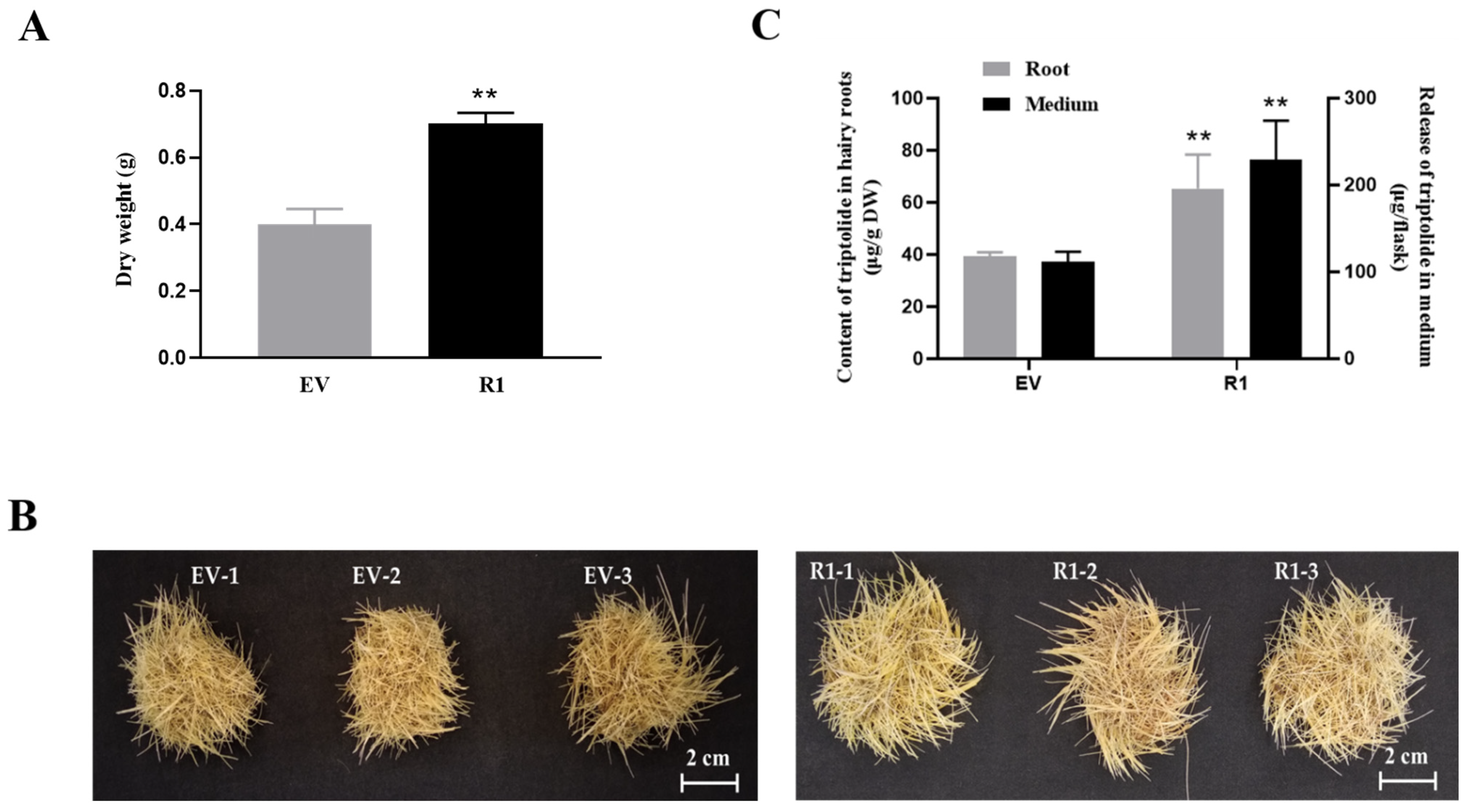
2.5. RNAi of TwMYC2a/b Increases Triptolide Production in the Hairy Roots of T. wilfordii
3. Discussion
4. Materials and Methods
4.1. Plant Materials, Growth Conditions, RNA Isolation, and cDNA Synthesis
4.2. Isolation and Bioinformatics Analyses of the TwMYC2a and TwMYC2b
4.3. Subcellular Localization of TwMYC2a and TwMYC2b
4.4. Quantitative Real-Time PCR (qRT-PCR) Analysis
4.5. Yeast One-Hybrid (Y1H) Assay
4.6. GUS Transactivation Assay
4.7. Construction of RNAi Vector and Generation of TwMYC2a/b Transgenic RNAi Hairy Roots
4.8. Transformant Selection and Transcript Analysis
4.9. High-Performance Liquid Chromatography (HPLC) Analysis
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sylvester, J.; Liacini, A.; Li, W.Q.; Dehnade, F.; Zafarullah, M. Tripterygium wilfordii Hook F Extract Suppresses Proinflammatory Cytokine-Induced Expression of Matrix Metalloproteinase Genes in Articular Chondrocytes by Inhibiting Activating Protein-1 and Nuclear Factor-KappaB Activities. Mol. Pharmacol. 2001, 59, 1196–1205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Dey, M.; Yang, H.; Poulev, A.; Pouleva, R.; Dorn, R.; Lipsky, P.E.; Kennelly, E.J.; Raskin, I. Anti-Inflammatory and Immunosuppressive Compounds from Tripterygium wilfordii. Phytochemistry 2007, 68, 1172–1178. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Li, C.; Ma, J.; Ni, L.; Huang, J.; Li, L.; Lin, M.; Hou, Q.; Zhang, D. Diterpenoids and Lignans from the Leaves of Tripterygium wilfordii. Fitoterapia 2018, 129, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Shen, S.-X.; Liu, L.-X.; Xu, J.-H.; Ma, X.-H.; Shi, D.-M.; Zhang, Z.-Q. A New Demethyl Abietane Diterpenoid from the Roots of Tripterygium wilfordii. Nat. Prod. Res. 2020, 34, 3094–3100. [Google Scholar] [CrossRef]
- Titov, D.V.; Gilman, B.; He, Q.-L.; Bhat, S.; Low, W.-K.; Dang, Y.; Smeaton, M.; Demain, A.L.; Miller, P.S.; Kugel, J.F.; et al. XPB, a Subunit of TFIIH, Is a Target of the Natural Product Triptolide. Nat. Chem. Biol. 2011, 7, 182–188. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.-L.; Yang, Y.-X.; Ding, J.; Li, Y.-C.; Miao, Z.-H. Triptolide: Structural Modifications, Structure-Activity Relationships, Bioactivities, Clinical Development and Mechanisms. Nat. Prod. Rep. 2012, 29, 457–475. [Google Scholar] [CrossRef]
- Zhou, H.; Liu, Y.; Wang, C.; Liu, L.; Wang, H.; Zhang, Y.; Long, C.; Sun, X. Triptolide Inhibits Epstein-Barr Nuclear Antigen 1 Expression by Increasing Sensitivity of Mitochondria Apoptosis of Nasopharyngeal Carcinoma Cells. J. Exp. Clin. Cancer Res. 2018, 37, 192. [Google Scholar] [CrossRef]
- Shi, Y.-F.; Liu, L.; He, L.-L.; Ye, J.; Lin, Z.-J.; Yuan, D.-L.; Deng, M.-M.; Fang, Z.-H.; Carter, B.Z.; Xu, B. Combining Triptolide with ABT-199 Is Effective against Acute Myeloid Leukemia through Reciprocal Regulation of Bcl-2 Family Proteins and Activation of the Intrinsic Apoptotic Pathway. Cell Death Dis. 2020, 11, 555. [Google Scholar] [CrossRef]
- Zhang, B.; Huo, Y.; Zhang, J.; Zhang, X.; Zhu, C. Agrobacterium Rhizogenes-Mediated RNAi of Tripterygium wilfordii and Application for Functional Study of Terpenoid Biosynthesis Pathway Genes. Ind. Crops Prod. 2019, 139, 111509. [Google Scholar] [CrossRef]
- Liu, Y.-J.; Zhao, Y.-J.; Zhang, M.; Su, P.; Wang, X.-J.; Zhang, X.-N.; Gao, W.; Huang, L.-Q. Cloning and Characterisation of the Gene Encoding 3-Hydroxy-3-Methylglutaryl-CoA Synthase in Tripterygium wilfordii. Molecules 2014, 19, 19696–19707. [Google Scholar] [CrossRef] [Green Version]
- Tong, Y.-R.; Zhang, Y.-F.; Zhao, Y.-J.; Hu, T.-Y.; Wang, J.-D.; Huang, L.-Q.; Gao, W. Differential Expression of the TwHMGS Gene and Its Effect on Triptolide Biosynthesis in Tripterygium wilfordii. Chin. J. Nat. Med. 2019, 17, 575–584. [Google Scholar] [CrossRef]
- Tong, Y.; Su, P.; Zhao, Y.; Zhang, M.; Wang, X.; Liu, Y.; Zhang, X.; Gao, W.; Huang, L. Molecular Cloning and Characterization of DXS and DXR Genes in the Terpenoid Biosynthetic Pathway of Tripterygium wilfordii. Int. J. Mol. Sci. 2015, 16, 25516–25535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zhao, Y.; Wang, J.; Hu, T.; Tong, Y.; Zhou, J.; Song, Y.; Gao, W.; Huang, L. Overexpression and RNA Interference of TwDXR Regulate the Accumulation of Terpenoid Active Ingredients in Tripterygium wilfordii. Biotechnol. Lett. 2018, 40, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhao, Y.; Wang, J.; Hu, T.; Tong, Y.; Zhou, J.; Gao, J.; Huang, L.; Gao, W. The Expression of TwDXS in the MEP Pathway Specifically Affects the Accumulation of Triptolide. Physiol. Plant. 2019. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Q.; Tong, Y.; Wang, Z.; Su, P.; Gao, W.; Huang, L. Molecular Cloning and Functional Identification of a CDNA Encoding 4-Hydroxy-3-Methylbut-2-Enyl Diphosphate Reductase from Tripterygium wilfordii. Acta Pharm. Sin. B 2017, 7, 208–214. [Google Scholar] [CrossRef]
- Tong, Y.; Zhang, M.; Su, P.; Zhao, Y.; Wang, X.; Zhang, X.; Gao, W.; Huang, L. Cloning and Functional Characterization of an Isopentenyl Diphosphate Isomerase Gene from Tripterygium wilfordii. Biotechnol. Appl. Biochem. 2016, 63, 863–869. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, Y.; Zhang, Y.; Su, P.; Hu, T.; Lu, Y.; Zhang, R.; Zhou, J.; Ma, B.; Gao, W.; et al. Overexpression and RNAi-Mediated Downregulation of TwIDI Regulates Triptolide and Celastrol Accumulation in Tripterygium wilfordii. Gene 2018, 679, 195–201. [Google Scholar] [CrossRef]
- Zhang, M.; Su, P.; Zhou, Y.-J.J.; Wang, X.-J.J.; Zhao, Y.-J.J.; Liu, Y.-J.J.; Tong, Y.-R.R.; Hu, T.-Y.Y.; Huang, L.-Q.Q.; Gao, W. Identification of Geranylgeranyl Diphosphate Synthase Genes from Tripterygium wilfordii. Plant Cell Rep. 2015, 34, 2179–2188. [Google Scholar] [CrossRef] [PubMed]
- Su, P.; Gao, L.; Tong, Y.; Guan, H.; Liu, S.; Zhang, Y.; Zhao, Y.; Wang, J.; Hu, T.; Tu, L.; et al. Analysis of the Role of Geranylgeranyl Diphosphate Synthase 8 from Tripterygium wilfordii in Diterpenoids Biosynthesis. Plant Sci. 2019, 285, 184–192. [Google Scholar] [CrossRef]
- Hansen, N.L.; Heskes, A.M.; Hamberger, B.; Olsen, C.E.; Hallström, B.M.; Andersen-Ranberg, J.; Hamberger, B. The Terpene Synthase Gene Family in Tripterygium wilfordii Harbors a Labdane-Type Diterpene Synthase among the Monoterpene Synthase TPS-b Subfamily. Plant J. 2017, 89, 429–441. [Google Scholar] [CrossRef] [Green Version]
- Su, P.; Guan, H.; Zhao, Y.; Tong, Y.; Xu, M.; Zhang, Y.; Hu, T.; Yang, J.; Cheng, Q.; Gao, L.; et al. Identification and Functional Characterization of Diterpene Synthases for Triptolide Biosynthesis from Tripterygium wilfordii. Plant J. Cell Mol. Biol. 2018, 93, 50–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, L.; Su, P.; Zhang, Z.; Gao, L.; Wang, J.; Hu, T.; Zhou, J.; Zhang, Y.; Zhao, Y.; Liu, Y.; et al. Genome of Tripterygium wilfordii and Identification of Cytochrome P450 Involved in Triptolide Biosynthesis. Nat. Commun. 2020, 11, 971. [Google Scholar] [CrossRef] [Green Version]
- Hong, G.-J.; Xue, X.-Y.; Mao, Y.-B.; Wang, L.-J.; Chen, X.-Y. Arabidopsis MYC2 Interacts with DELLA Proteins in Regulating Sesquiterpene Synthase Gene Expression. Plant Cell 2012, 24, 2635–2648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aslam, M.Z.; Lin, X.; Li, X.; Yang, N.; Chen, L. Molecular Cloning and Functional Characterization of CpMYC2 and CpbHLH13 Transcription Factors from Wintersweet (Chimonanthus praecox L.). Plants 2020, 9, 785. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Sun, W.; Chen, J.; Tan, H.; Xiao, Y.; Li, Q.; Ji, Q.; Gao, S.; Chen, L.; Chen, S.; et al. SmMYC2a and SmMYC2b Played Similar but Irreplaceable Roles in Regulating the Biosynthesis of Tanshinones and Phenolic Acids in Salvia miltiorrhiza. Sci. Rep. 2016, 6, 22852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, A.; Rather, G.A.; Misra, P.; Dhar, M.K.; Lattoo, S.K. Jasmonate Responsive Transcription Factor WsMYC2 Regulates the Biosynthesis of Triterpenoid Withanolides and Phytosterol via Key Pathway Genes in Withania somnifera (L.) Dunal. Plant Mol. Biol. 2019, 100, 543–560. [Google Scholar] [CrossRef]
- Dombrecht, B.; Xue, G.P.; Sprague, S.J.; Kirkegaard, J.A.; Ross, J.J.; Reid, J.B.; Fitt, G.P.; Sewelam, N.; Schenk, P.M.; Manners, J.M.; et al. MYC2 Differentially Modulates Diverse Jasmonate-Dependent Functions in Arabidopsis. Plant Cell 2007, 19, 2225–2245. [Google Scholar] [CrossRef] [Green Version]
- Lenka, S.K.; Nims, N.E.; Vongpaseuth, K.; Boshar, R.A.; Roberts, S.C.; Walker, E.L. Jasmonate-Responsive Expression of Paclitaxel Biosynthesis Genes in Taxus cuspidata Cultured Cells Is Negatively Regulated by the bHLH Transcription Factors TcJAMYC1, TcJAMYC2, and TcJAMYC4. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Atchley, W.R.; Fitch, W.M. A Natural Classification of the Basic Helix-Loop-Helix Class of Transcription Factors. Proc. Natl. Acad. Sci. USA 1997, 94, 5172–5176. [Google Scholar] [CrossRef] [Green Version]
- Abe, H.; Urao, T.; Ito, T.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) Function as Transcriptional Activators in Abscisic Acid Signaling. Plant Cell 2003, 15, 63–78. [Google Scholar] [CrossRef] [Green Version]
- Carretero-Paulet, L.; Galstyan, A.; Roig-Villanova, I.; Martínez-García, J.F.; Bilbao-Castro, J.R.; Robertson, D.L. Genome-Wide Classification and Evolutionary Analysis of the bHLH Family of Transcription Factors in Arabidopsis, Poplar, Rice, Moss, and Algae. Plant Physiol. 2010, 153, 1398–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, D.-G.; Yu, J.-Q.; Han, P.-L.; Xie, X.-B.; Sun, C.-H.; Zhang, Q.-Y.; Wang, J.-H.; Hao, Y.-J. The Regulatory Module MdPUB29-MdbHLH3 Connects Ethylene Biosynthesis with Fruit Quality in Apple. New Phytol. 2019, 221, 1966–1982. [Google Scholar] [CrossRef] [PubMed]
- Xing, B.; Yang, D.; Yu, H.; Zhang, B.; Yan, K.; Zhang, X.; Han, R.; Liang, Z. Overexpression of SmbHLH10 Enhances Tanshinones Biosynthesis in Salvia miltiorrhiza Hairy Roots. Plant Sci. 2018, 276, 229–238. [Google Scholar] [CrossRef]
- Xu, Y.-H.; Liao, Y.-C.; Lv, F.-F.; Zhang, Z.; Sun, P.-W.; Gao, Z.-H.; Hu, K.-P.; Sui, C.; Jin, Y.; Wei, J.-H. Transcription Factor AsMYC2 Controls the Jasmonate-Responsive Expression of ASS1 Regulating Sesquiterpene Biosynthesis in Aquilaria sinensis (Lour.) Gilg. Plant Cell Physiol. 2017, 58, 1924–1933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huo, Y.; Zhang, B.; Chen, L.; Zhang, J.; Zhang, X.; Zhu, C. Isolation and Functional Characterization of the Promoters of Miltiradiene Synthase Genes, TwTPS27a and TwTPS27b, and Interaction Analysis with the Transcription Factor TwTGA1 from Tripterygium wilfordii. Plants 2021, 10, 418. [Google Scholar] [CrossRef] [PubMed]
- Hilson, P.; Allemeersch, J.; Altmann, T.; Aubourg, S.; Avon, A.; Beynon, J.; Bhalerao, R.P.; Bitton, F.; Caboche, M.; Cannoot, B.; et al. Versatile Gene-Specific Sequence Tags for Arabidopsis Functional Genomics: Transcript Profiling and Reverse Genetics Applications. Genome Res. 2004, 14, 2176–2189. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Hedhili, S.; Montiel, G.; Zhang, Y.; Chatel, G.; Pré, M.; Gantet, P.; Memelink, J. The Basic Helix-Loop-Helix Transcription Factor CrMYC2 Controls the Jasmonate-Responsive Expression of the ORCA Genes That Regulate Alkaloid Biosynthesis in Catharanthus roseus. Plant J. Cell Mol. Biol. 2011, 67, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Todd, A.T.; Liu, E.; Polvi, S.L.; Pammett, R.T.; Page, J.E. A Functional Genomics Screen Identifies Diverse Transcription Factors That Regulate Alkaloid Biosynthesis in Nicotiana benthamiana. Plant J. Cell Mol. Biol. 2010, 62, 589–600. [Google Scholar] [CrossRef]
- Yamada, Y.; Kokabu, Y.; Chaki, K.; Yoshimoto, T.; Ohgaki, M.; Yoshida, S.; Kato, N.; Koyama, T.; Sato, F. Isoquinoline Alkaloid Biosynthesis Is Regulated by a Unique bHLH-Type Transcription Factor in Coptis japonica. Plant Cell Physiol. 2011, 52, 1131–1141. [Google Scholar] [CrossRef] [Green Version]
- Hao, X.; Xie, C.; Ruan, Q.; Zhang, X.; Wu, C.; Han, B.; Qian, J.; Zhou, W.; Nützmann, H.-W.; Kai, G. The Transcription Factor OpWRKY2 Positively Regulates the Biosynthesis of the Anticancer Drug Camptothecin in Ophiorrhiza pumila. Hortic. Res. 2021, 8, 7. [Google Scholar] [CrossRef] [PubMed]
- Kazan, K.; Manners, J.M. MYC2: The Master in Action. Mol. Plant 2013, 6, 686–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Jin, X.; Chen, Y.; Wei, M.; Liao, W.; Zhao, S.; Fu, C.; Yu, L. TcMYC2a, a Basic Helix–Loop–Helix Transcription Factor, Transduces JA-Signals and Regulates Taxol Biosynthesis in Taxus chinensis. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef]
- Zhang, H.-B.; Bokowiec, M.T.; Rushton, P.J.; Han, S.-C.; Timko, M.P. Tobacco Transcription Factors NtMYC2a and NtMYC2b Form Nuclear Complexes with the NtJAZ1 Repressor and Regulate Multiple Jasmonate-Inducible Steps in Nicotine Biosynthesis. Mol. Plant 2012, 5, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.; Guo, D.; Yang, S.; Shi, M.; Chao, J.; Li, H.; Peng, S.; Tian, W. Jasmonate Signalling in the Regulation of Rubber Biosynthesis in Laticifer Cells of Rubber Tree, Hevea brasiliensis. J. Exp. Bot. 2018, 69, 3559–3571. [Google Scholar] [CrossRef] [PubMed]
- Guo, D.; Li, H.-L.; Wang, Y.; Zhu, J.-H.; Peng, S.-Q. A Myelocytomatosis Transcription Factor from Hevea brasiliensis Positively Regulates the Expression of the Small Rubber Particle Protein Gene. Ind. Crops Prod. 2019, 133, 90–97. [Google Scholar] [CrossRef]
- Yang, N.; Zhou, W.; Su, J.; Wang, X.; Li, L.; Wang, L.; Cao, X.; Wang, Z. Overexpression of SmMYC2 Increases the Production of Phenolic Acids in Salvia miltiorrhiza. Front. Plant Sci. 2017, 8, 1804. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Salasini, B.C.; Khan, M.; Devi, B.; Bush, M.; Subramaniam, R.; Hepworth, S.R. Clade I TGACG-Motif Binding Basic Leucine Zipper Transcription Factors Mediate BLADE-ON-PETIOLE-Dependent Regulation of Development. Plant Physiol. 2019, 180, 937–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Budimir, J.; Treffon, K.; Nair, A.; Thurow, C.; Gatz, C. Redox-Active Cysteines in TGACG-BINDING FACTOR 1 (TGA1) Do Not Play a Role in Salicylic Acid or Pathogen-Induced Expression of TGA1-Regulated Target Genes in Arabidopsis Thaliana. New Phytol. 2020. [Google Scholar] [CrossRef]
- Sun, T.; Busta, L.; Zhang, Q.; Ding, P.; Jetter, R.; Zhang, Y. TGACG-BINDING FACTOR 1 (TGA1) and TGA4 Regulate Salicylic Acid and Pipecolic Acid Biosynthesis by Modulating the Expression of SYSTEMIC ACQUIRED RESISTANCE DEFICIENT 1 (SARD1) and CALMODULIN-BINDING PROTEIN 60g (CBP60g). New Phytol. 2018, 217, 344–354. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.; Sun, Q.; Li, J.; Ahammed, G.J.; Yu, J.; Fang, H.; Xia, X. Glutaredoxin S25 and Its Interacting TGACG Motif-Binding Factor TGA2 Mediate Brassinosteroid-Induced Chlorothalonil Metabolism in Tomato Plants. Environ. Pollut. 2019, 255, 113256. [Google Scholar] [CrossRef]
- Gatz, C. From Pioneers to Team Players: TGA Transcription Factors Provide a Molecular Link between Different Stress Pathways. Mol. Plant-Microbe Interact. 2013, 26, 151–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.H.; Yoo, C.M.; Hong, A.P.; Kim, S.H.; Jeong, H.J.; Shin, S.Y.; Kim, H.J.; Yun, D.-J.; Lim, C.O.; Bahk, J.D.; et al. DNA-Binding Study Identifies C-Box and Hybrid C/G-Box or C/A-Box Motifs as High-Affinity Binding Sites for STF1 and LONG HYPOCOTYL5 Proteins. Plant Physiol. 2008, 146, 1862–1877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boter, M.; Ruíz-Rivero, O.; Abdeen, A.; Prat, S. Conserved MYC Transcription Factors Play a Key Role in Jasmonate Signaling Both in Tomato and Arabidopsis. Genes Dev. 2004, 18, 1577–1591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenzo, O.; Chico, J.M.; Sánchez-Serrano, J.J.; Solano, R. JASMONATE-INSENSITIVE1 Encodes a MYC Transcription Factor Essential to Discriminate between Different Jasmonate-Regulated Defense Responses in Arabidopsis. Plant Cell 2004, 16, 1938–1950. [Google Scholar] [CrossRef] [Green Version]
- Shen, Q.; Lu, X.; Yan, T.; Fu, X.; Lv, Z.; Zhang, F.; Pan, Q.; Wang, G.; Sun, X.; Tang, K. The Jasmonate-Responsive AaMYC2 Transcription Factor Positively Regulates Artemisinin Biosynthesis in Artemisia annua. New Phytol. 2016, 210, 1269–1281. [Google Scholar] [CrossRef] [Green Version]
- An, J.-P.; Li, H.-H.; Song, L.-Q.; Su, L.; Liu, X.; You, C.-X.; Wang, X.-F.; Hao, Y.-J. The Molecular Cloning and Functional Characterization of MdMYC2, a bHLH Transcription Factor in Apple. Plant Physiol. Biochem. 2016, 108, 24–31. [Google Scholar] [CrossRef]
- Zhou, Y.; Feng, J.; Li, Q.; Huang, D.; Chen, X.; Du, Z.; Lv, Z.; Xiao, Y.; Han, Y.; Chen, J.; et al. SmMYC2b Enhances Tanshinone Accumulation in Salvia miltiorrhiza by Activating Pathway Genes and Promoting Lateral Root Development. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- Goossens, J.; Mertens, J.; Goossens, A. Role and Functioning of bHLH Transcription Factors in Jasmonate Signalling. J. Exp. Bot. 2017, 68, 1333–1347. [Google Scholar] [CrossRef] [PubMed]
- Nakata, M.; Mitsuda, N.; Herde, M.; Koo, A.J.K.; Moreno, J.E.; Suzuki, K.; Howe, G.A.; Ohme-Takagi, M. A bHLH-Type Transcription Factor, ABA-INDUCIBLE bHLH-TYPE TRANSCRIPTION FACTOR/JA-ASSOCIATED MYC2-LIKE1, Acts as a Repressor to Negatively Regulate Jasmonate Signaling in Arabidopsis. Plant Cell 2013, 25, 1641–1656. [Google Scholar] [CrossRef] [Green Version]
- Sasaki-Sekimoto, Y.; Jikumaru, Y.; Obayashi, T.; Saito, H.; Masuda, S.; Kamiya, Y.; Ohta, H.; Shirasu, K. Basic Helix-Loop-Helix Transcription Factors JASMONATE-ASSOCIATED MYC2-LIKE1 (JAM1), JAM2, and JAM3 Are Negative Regulators of Jasmonate Responses in Arabidopsis. Plant Physiol. 2013, 163, 291–304. [Google Scholar] [CrossRef] [Green Version]
- Bai, Z.; Li, W.; Jia, Y.; Yue, Z.; Jiao, J.; Huang, W.; Xia, P.; Liang, Z. The Ethylene Response Factor SmERF6 Co-Regulates the Transcription of SmCPS1 and SmKSL1 and Is Involved in Tanshinone Biosynthesis in Salvia miltiorrhiza Hairy Roots. Planta 2018. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ji, A.; Xu, Z.; Luo, H.; Song, J. The AP2/ERF Transcription Factor SmERF128 Positively Regulates Diterpenoid Biosynthesis in Salvia miltiorrhiza. Plant Mol. Biol. 2019, 100, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Xing, B.; Yang, D.; Ren, M.; Guo, H.; Yang, S.; Liang, Z. SmbHLH3 Acts as a Transcription Repressor for Both Phenolic Acids and Tanshinone Biosynthesis in Salvia miltiorrhiza Hairy Roots. Phytochemistry 2020, 169, 112183. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Liu, H.; Wang, S.; Wang, C.; Miao, G. A Class I TGA Transcription Factor from Tripterygium wilfordii Hook.f. Modulates the Biosynthesis of Secondary Metabolites in Both Native and Heterologous Hosts. Plant Sci. 2020, 290, 110293. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Miao, G.; Zhu, C.; Feng, J.; Han, J.; Song, X.-W.; Zhang, X. Aggregate Cell Suspension Cultures of Tripterygium wilfordii Hook. f. for Triptolide, Wilforgine, and Wilforine Production. Plant Cell Tissue Organ Cult. PCTOC 2013, 112, 109–116. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huo, Y.; Zhang, J.; Zhang, B.; Chen, L.; Zhang, X.; Zhu, C. MYC2 Transcription Factors TwMYC2a and TwMYC2b Negatively Regulate Triptolide Biosynthesis in Tripterygium wilfordii Hairy Roots. Plants 2021, 10, 679. https://doi.org/10.3390/plants10040679
Huo Y, Zhang J, Zhang B, Chen L, Zhang X, Zhu C. MYC2 Transcription Factors TwMYC2a and TwMYC2b Negatively Regulate Triptolide Biosynthesis in Tripterygium wilfordii Hairy Roots. Plants. 2021; 10(4):679. https://doi.org/10.3390/plants10040679
Chicago/Turabian StyleHuo, Yanbo, Jing Zhang, Bin Zhang, Ling Chen, Xing Zhang, and Chuanshu Zhu. 2021. "MYC2 Transcription Factors TwMYC2a and TwMYC2b Negatively Regulate Triptolide Biosynthesis in Tripterygium wilfordii Hairy Roots" Plants 10, no. 4: 679. https://doi.org/10.3390/plants10040679
APA StyleHuo, Y., Zhang, J., Zhang, B., Chen, L., Zhang, X., & Zhu, C. (2021). MYC2 Transcription Factors TwMYC2a and TwMYC2b Negatively Regulate Triptolide Biosynthesis in Tripterygium wilfordii Hairy Roots. Plants, 10(4), 679. https://doi.org/10.3390/plants10040679