
Uncovering the Neglected Floral Secretory Structures of Rhamnaceae and Their Functional and Systematic Significance


Abstract
:1. Introduction
2. Results
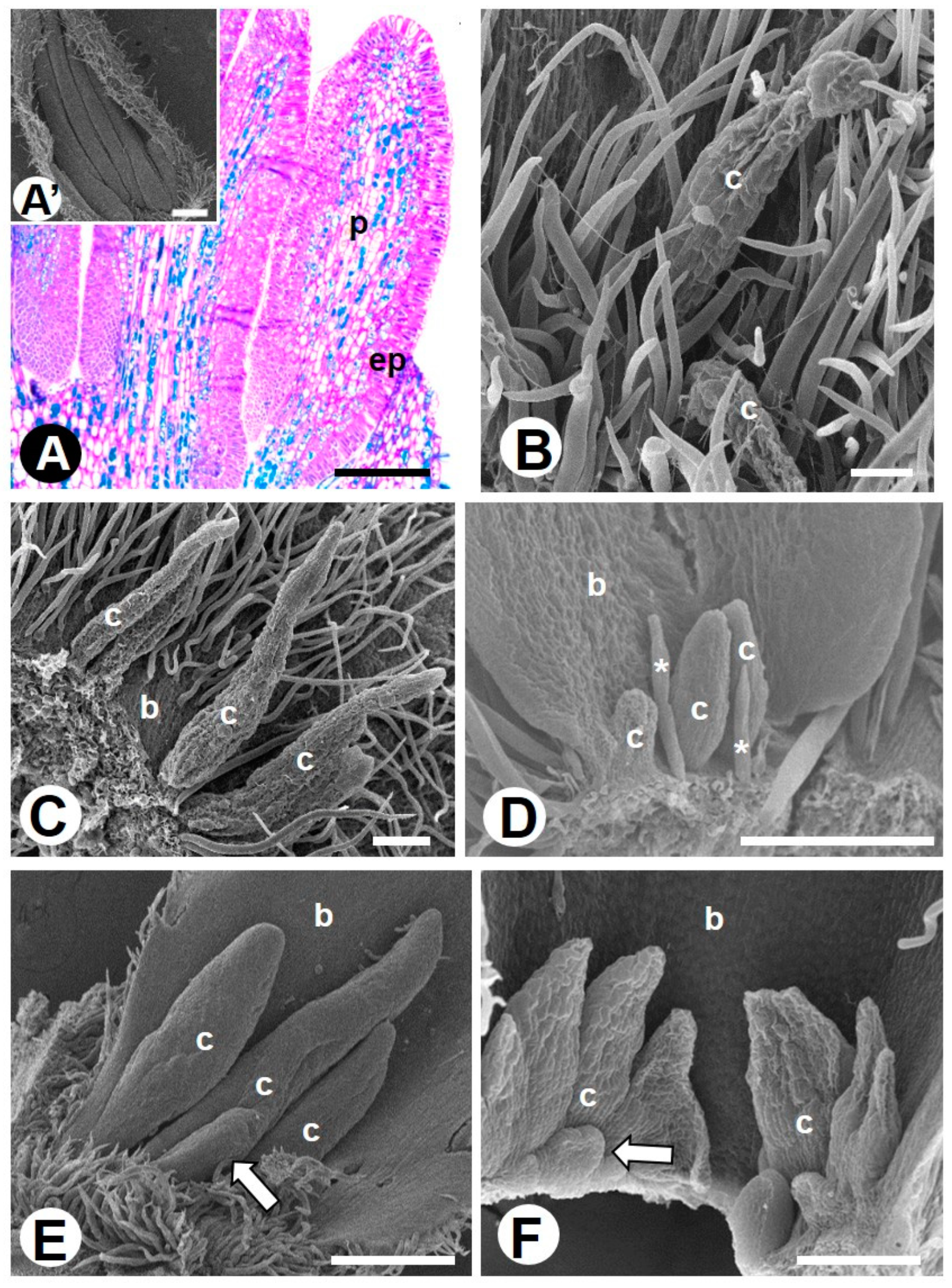
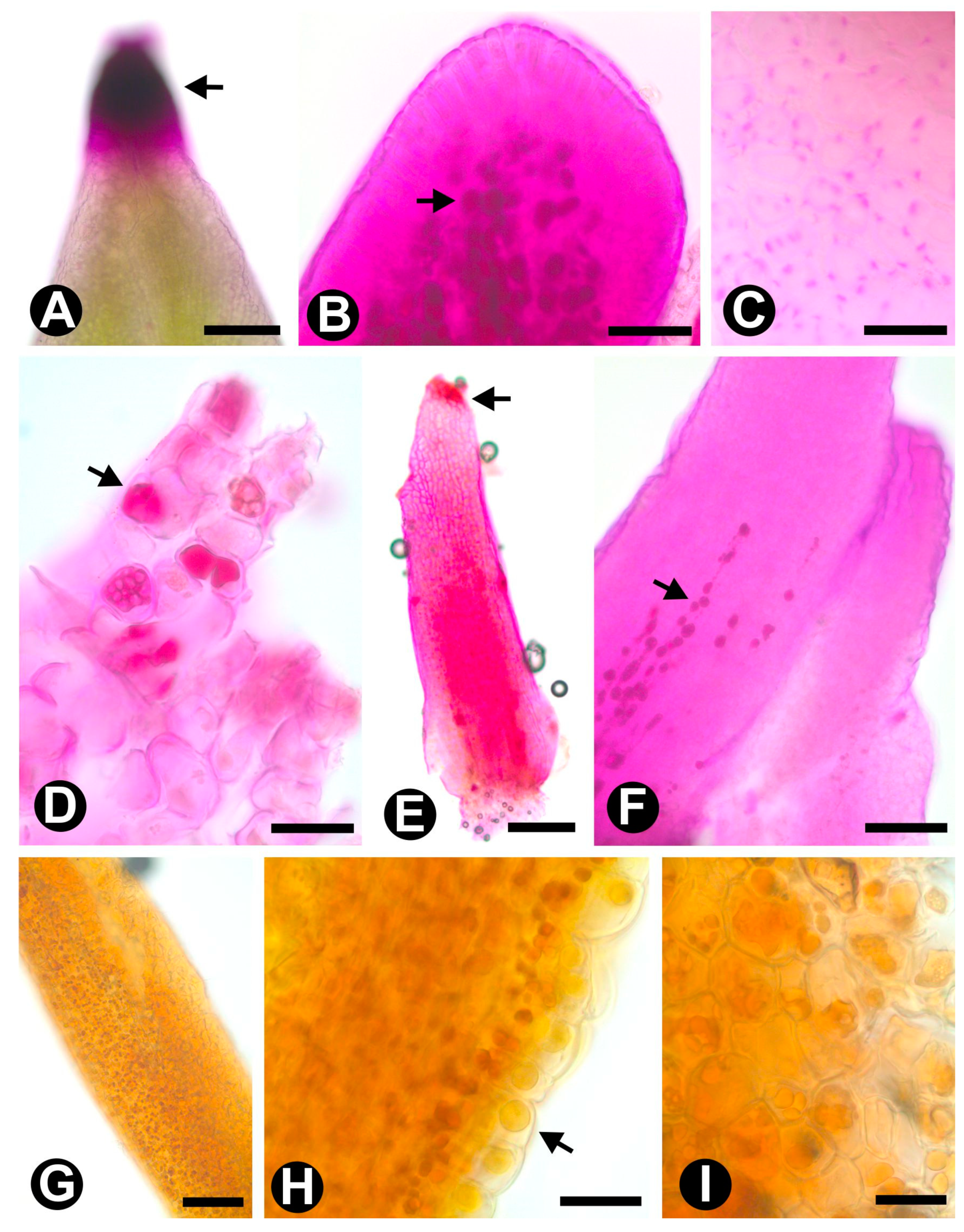
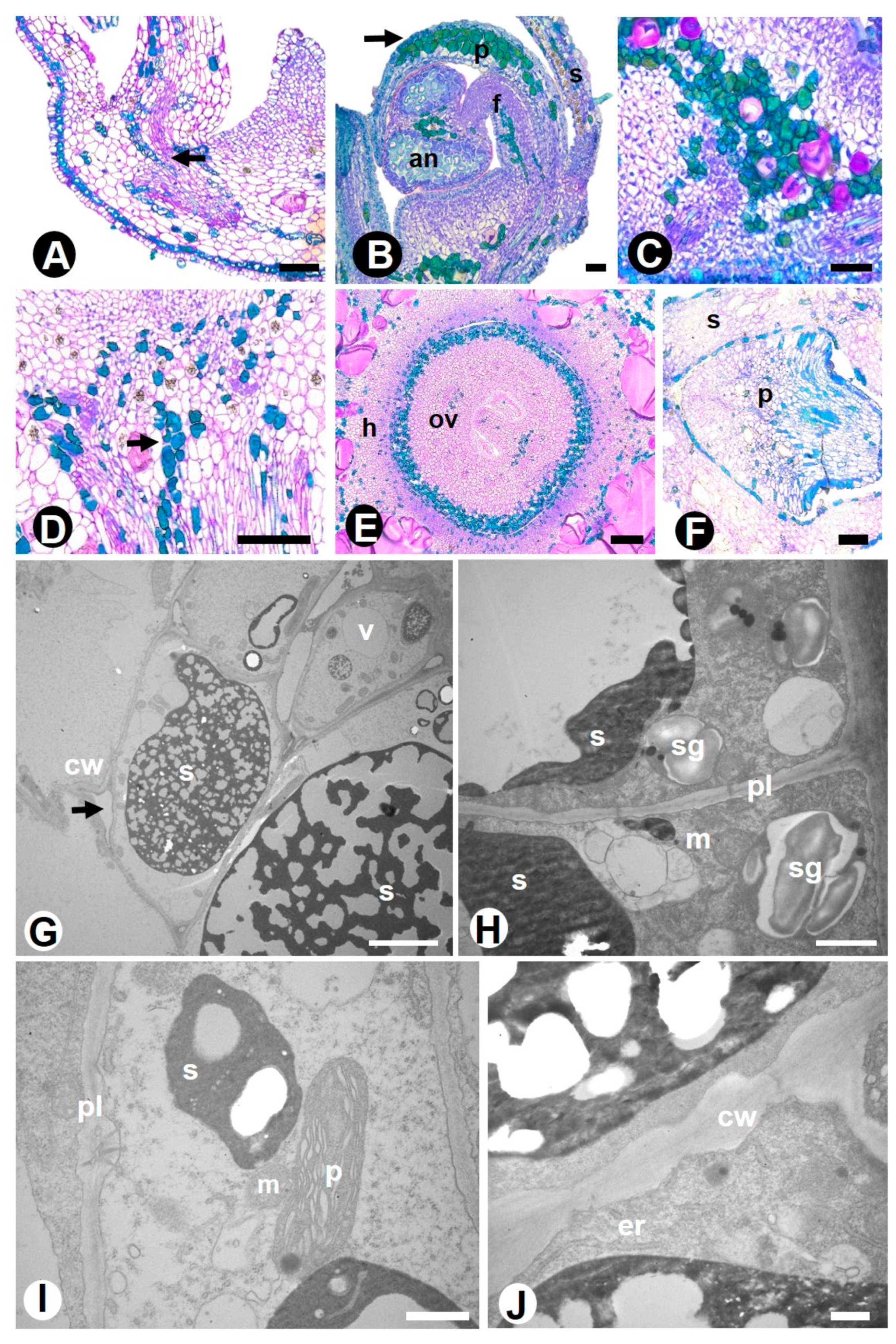
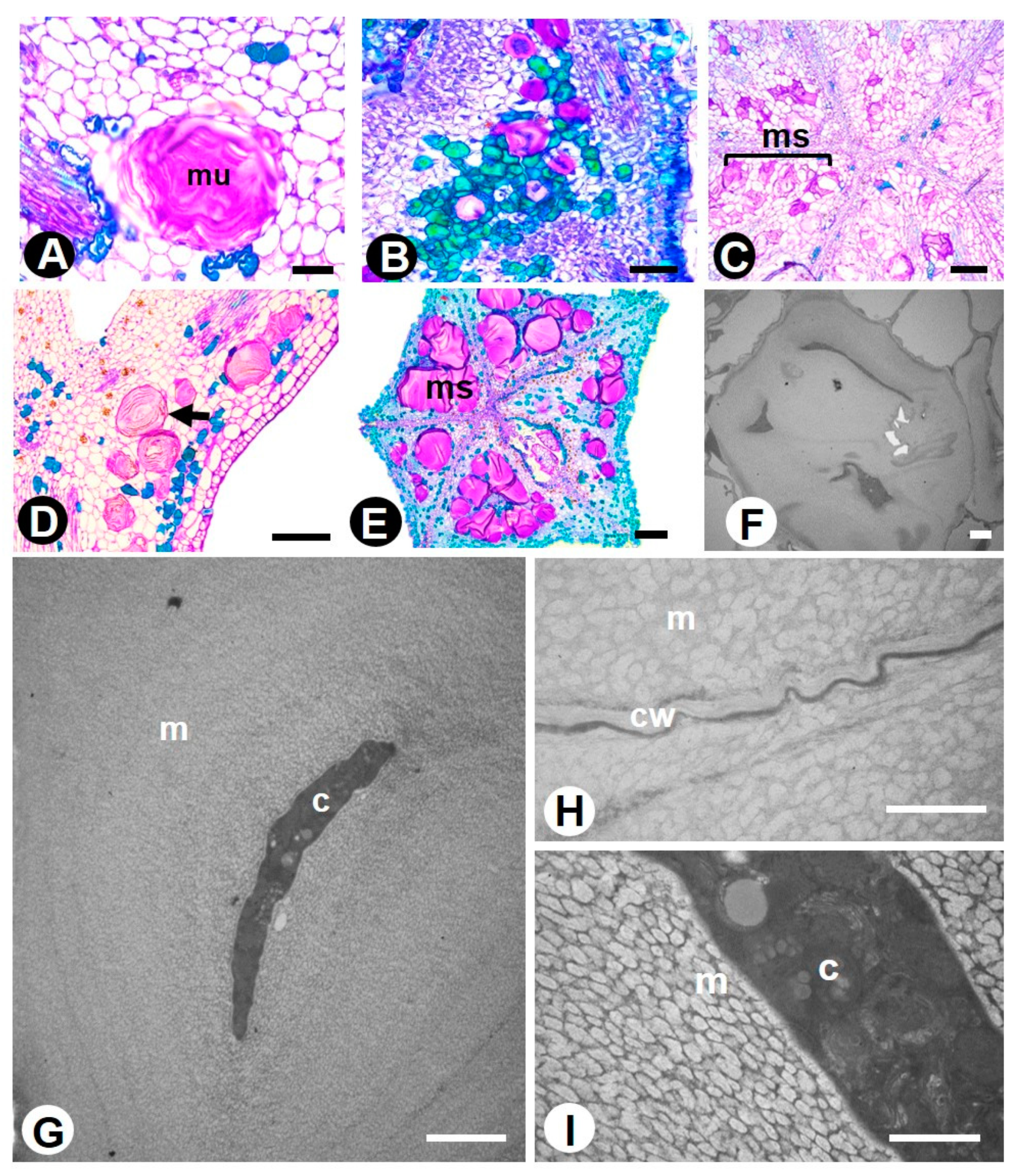
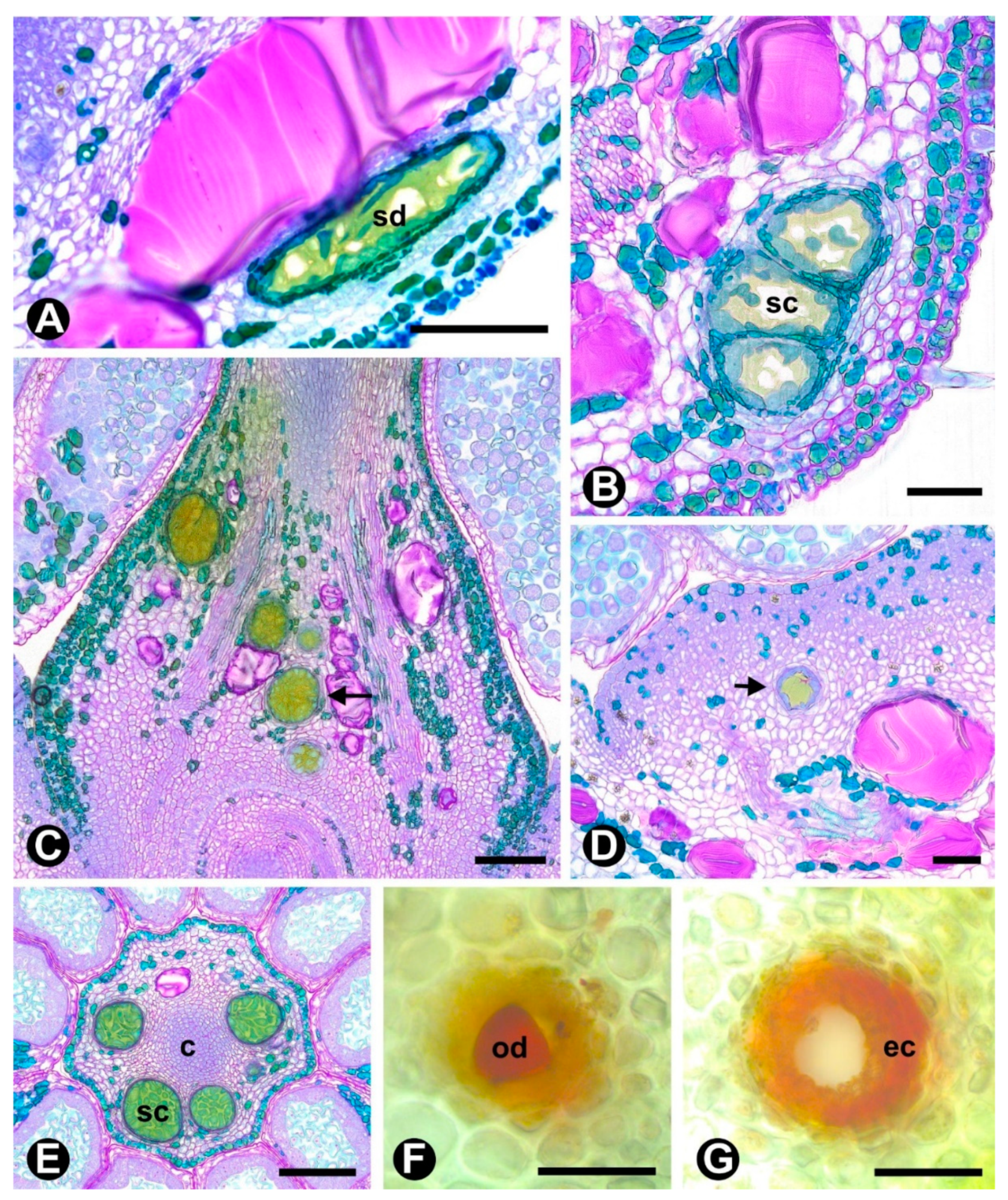
2.1. Novel Data
2.2. Data Obtained from the Literature
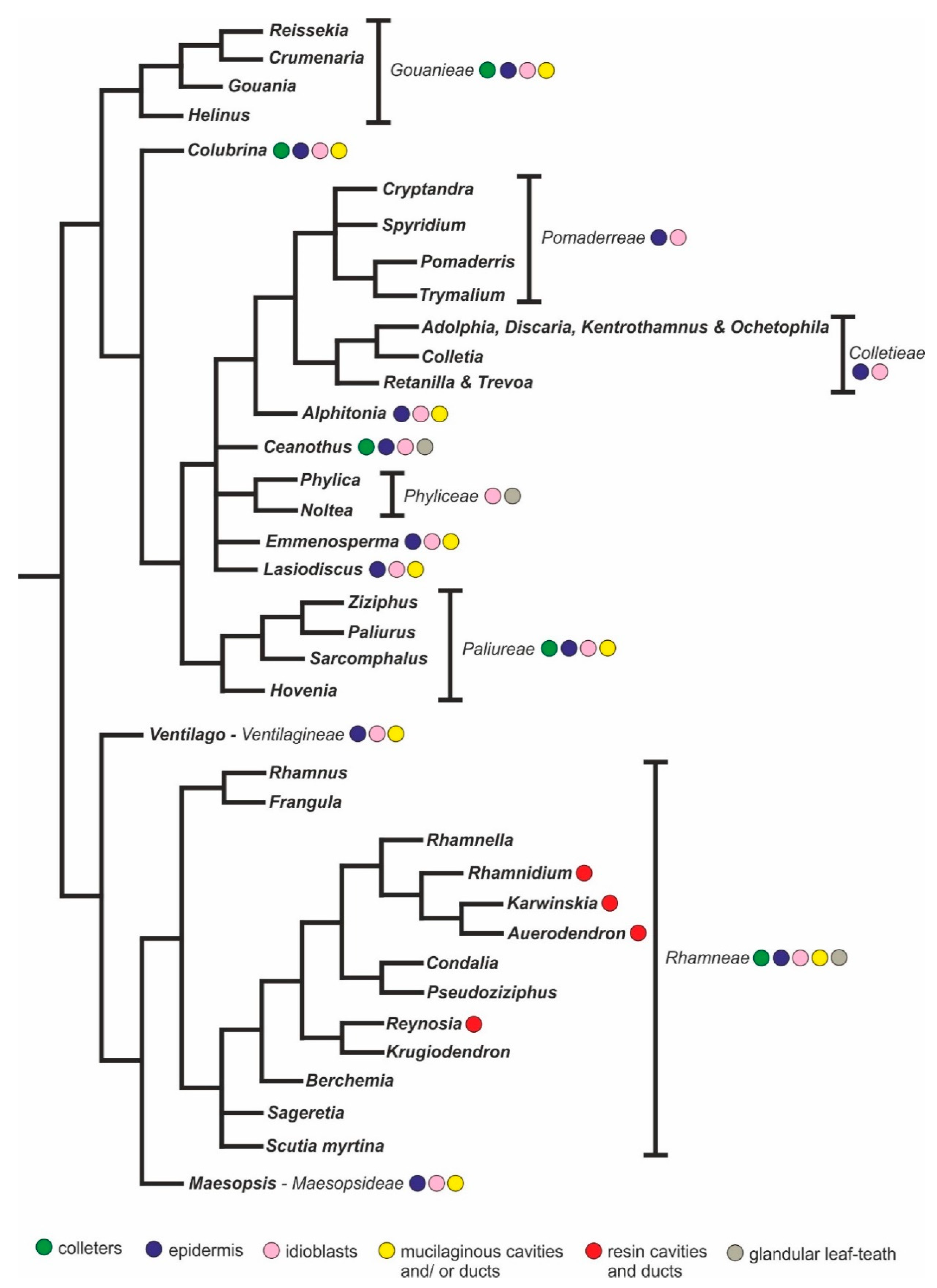
3. Discussion
3.1. An Overview of the Secretory Structures of Rhamnaceae
3.2. Outlook
4. Materials and Methods
4.1. Plant Material
4.2. Preparation and Fixation of Samples
4.3. Surface Analysis (SEM)
4.4. Anatomical Analysis (LM)
4.5. Ultrastructural Analysis (TEM)
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- The Angiosperm Phylogeny Group; Chase, M.W.; Christenhusz, M.J.M.; Fay, M.F.; Byng, J.W.; Judd, W.S.; Soltis, D.E.; Mabberley, D.J.; Sennikov, A.N.; Soltis, P.S.; et al. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar]
- Rhamnaceae in Flora do Brasil 2020 em Construção. Jardim Botânico do Rio de Janeiro. Available online: http://floradobrasil.jbrj.gov.br/reflora/floradobrasil/FB207 (accessed on 9 October 2020).
- Medan, D. Reproductive phenology, pollination biology, and gynoecium development in Discaria americana (Rhamnaceae). N. Z. J. Bot. 1991, 29, 31–42. [Google Scholar] [CrossRef]
- Medan, D.; D’Ambrogio, A.C. Reproductive biology of the andromonoecious shrub Trevoa quinquenervia (Rhamnaceae). Bot. J. Linn. Soc. 1998, 126, 191–206. [Google Scholar] [CrossRef]
- Medan, D.; Arce, M.E. Reproductive biology of the Andean-disjunct genus Retanilla (Rhamnaceae). Plant Syst. Evol. 1999, 218, 281–298. [Google Scholar] [CrossRef]
- Medan, D.; Toretta, J.P. The reproduction of Colletia hystrix and late flowering in Colletia (Rhamnaceae: Colletieae). Plant Syst. Evol. 2014, 301, 1181–1189. [Google Scholar] [CrossRef]
- Cerino, M.C.; Richard, G.A.; Torretta, J.P.; Gutierrez, H.F.; Pensiero, J.F. Reproductive biology of Ziziphus mistol Griseb. (Rhamnaceae), a wild fruit tree of saline environments. Flora Morphol. Distrib. Funct. Ecol. Plants 2015, 211, 18–25. [Google Scholar] [CrossRef]
- Medan, D.; Aagesen, L. Comparative flower and fruit structure in the Colletieae (Rhamnaceae). Bot. Jahrb. Syst. 1995, 117, 531–564. [Google Scholar]
- Gotelli, M.M.; Galati, B.G.; Medan, D. Morphological and ultrastructural studies of floral nectaries in Rhamnaceae. J. Torrey Bot. Soc. 2017, 144, 63–73. [Google Scholar] [CrossRef]
- Gemoll, K. Anatomisch-systematische untersuchung des blattes der Rhamneen aus den triben: Rhamneen, Colletieen und Gouanieen. Beihefte Botanischen Centralblatt 1902, 12, 351–424. [Google Scholar]
- Herzog, T.H. Anatomisch-systematische untersuchung des blattes der Rhamneen aus den triben: Ventilagineen, Zizypheen und Rhamneen. Beihefte Botanischen Centralblatt 1903, 15, 95–207. [Google Scholar]
- Solereder, H. Systematic Anatomy of the Dicotyledons: A Handbook for Laboratories of Pure and Applied Botany; Clarendon Press: Oxford, UK, 1908; Volumes 1 and 2, pp. 218–221, 885–889. [Google Scholar]
- Metcalfe, C.R.; Chalk, L. Anatomy of the Dicotyledons: Leaves, Stem and Wood in Relation to Taxonomy with Notes on Economic Uses; Clarendon Press: Oxford, UK, 1957; Volume 1, pp. 404–413. [Google Scholar]
- Medan, D. Anatomia foliar de Kentrothamnus (Rhamnaceae). Bol. Soc. Argent. Bot. 1974, 16, 83–88. [Google Scholar]
- Gregory, M.; Baas, P. A survey of mucilage cells in vegetative organs of the dicotyledons. Isr. J. Plant Sci. 1989, 38, 125–174. [Google Scholar]
- Mantese, A.; Medan, D. Anatomia y arquitectura foliares de Retanilla (Rhamnaceae). Darwiniana 1992, 31, 253–259. [Google Scholar]
- Clifford, S.C.; Arndt, S.K.; Popp, M.; Jones, H.G. Mucilages and polysaccharides in Ziziphus species (Rhamnaceae): Localization, composition and physiological roles during drought-stress. J. Exp. Bot. 2002, 53, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Medan, D.; Schirarend, C. Rhamnaceae. In Flowering Plants: Dicotyledons; Kubitzki, K., Ed.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 320–338. [Google Scholar]
- Serdar, B.; Çoskunçelebi, K.; Terzioglu, S.; Hampe, A. Anatomical notes on Turkish Frangula alnus Mill. (Rhamnaceae). Plant Biosyst. 2007, 141, 69–74. [Google Scholar] [CrossRef] [Green Version]
- Shisode, S.B.; Patil, D.A. Petiolar anatomy in some Rhamnaceae. Curr. Bot. 2011, 2, 22–25. [Google Scholar]
- Sivasankari, M.P.; Sankaravadivoo, A. Leaf anatomy of Ziziphus mauritiana Lam. Int. J. Curr. Res. Biosci. Plant Biol. 2017, 4, 73–79. [Google Scholar] [CrossRef]
- Arsenijević, J.; Drobac, M.; Slavkovska, V.; Kovačević, N.; Lakušić, B. Anatomical analysis and phytochemical screening of Frangula rupestris (Scop.) Schur (Rhamnaceae). Bot. Serb. 2018, 42, 231–239. [Google Scholar]
- Schirarend, C.; Hoffmann, P. Untersuchungen zur blütenmorphologie der gattung Reynosia Griseb. (Rhamnaceae). Flora 1993, 188, 275–286. [Google Scholar] [CrossRef]
- Gotelli, M.M.; Galati, B.G.; Zarlavsky, G.; Nicolau, M.; Reposi, S. Localization, morphology, anatomy and ultrastructure of osmophores in species of Rhamnaceae. Protoplasma 2020, 257, 1109–1121. [Google Scholar] [CrossRef] [PubMed]
- Asgarpanah, J.; Haghighat, E. Phytochemistry and pharmacologic properties of Ziziphus spina christi (L.) Willd. Afr. J. Pharm. Pharmacol. 2012, 6, 2332–2339. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Zaher, A.O.; Salim, S.Y.; Assaf, M.H.; Abdel-Hady, R.H. Antidiabetic activity and toxicity of Zizyphus spina-christi leaves. J. Ethnopharmacol. 2005, 101, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Pawlowska, A.M.; Camangi, F.; Bader, A.; Braca, A. Flavonoids of Zizyphus jujuba L. and Zizyphus spina-christi (L.) Willd (Rhamnaceae) fruits. Food Chem. 2009, 112, 858–862. [Google Scholar] [CrossRef]
- Hyun, T.K.; Eom, S.H.; Yu, C.Y.; Roitsch, T. Hovenia dulcis—An Asian traditional herb. Planta Med. 2010, 76, 943–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abalaka, M.E.; Mann, A.; Adeyemo, S.O. Studies on in-vitro antioxidant and free radical scavenging potential and phytochemical screening of leaves of Ziziphus mauritiana L. and Ziziphus spina-christi L. compared with ascorbic acid. J. Med. Genet. Genom. 2011, 3, 28–34. [Google Scholar]
- Wang, M.; Jiang, C.; Ma, L.; Zhang, Z.; Cao, L.; Liu, J.; Zeng, X. Preparation, preliminary characterization and immunostimulatory activity of polysaccharide fractions from the peduncles of Hovenia dulcis. Food Chem. 2013, 138, 41–47. [Google Scholar] [CrossRef]
- Ashraf, A.; Sarfrazi, R.A.; Anwar, F.; Shahid, S.A.; Alkharfy, K.M. Chemical composition and biological activities of leaves of Ziziphus mauritiana L. native to Pakistan. Pak. J. Bot. 2015, 47, 367–376. [Google Scholar]
- Brito, S.M.O.; Coutinho, H.D.M.; Talvani, A.; Coronel, C.; Barbosa, A.G.R.; Vega, C.; Figueredo, F.; Tintino, S.; Lima, L.F.; Boligon, A.A.; et al. Analysis of bioactivities and chemical composition of Ziziphus joazeiro Mart. using HPLC–DAD. Food Chem. 2015, 186, 185–191. [Google Scholar] [CrossRef] [Green Version]
- Ji, X.; Peng, Q.; Yuan, Y.; Shen, J.; Xie, X.; Wang, M. Isolation, structures and bioactivities of the polysaccharides from jujube fruit (Ziziphus jujuba Mill.): A review. Food Chem. 2017, 227, 349–357. [Google Scholar] [CrossRef]
- Hussein, A.S. Ziziphus spina-christi: Analysis of bioactivities and chemical composition. In Wild Fruits: Composition, Nutritional Value and Products; Mariod, A.A., Ed.; Springer: Cham, Switzerland, 2019; pp. 175–197. [Google Scholar]
- Yang, B.; Wu, Q.; Luo, Y.; Yang, Q.; Chen, G.; Wei, X.; Kan, J. Japanese grape (Hovenia dulcis) polysaccharides: New insight into extraction, characterization, rheological properties, and bioactivities. Int. J. Biol. Macromol. 2019, 134, 631–644. [Google Scholar] [CrossRef]
- Mahajan, R.T.; Chopda, M.Z. Phyto-pharmacology of Ziziphus jujuba Mill—A plant review. Pharmacogn. Rev. 2009, 3, 320–329. [Google Scholar]
- Silva, L.A.; Sales, J.F.; Guimarães, R.M.; Oliveira, J.A.; Vasconcelos Filho, S.C. Morphological aspects of fruits, seeds and seedlings of Rhamnidium elaeocarpum Reissek. Semin. Cienc. Agrar. 2015, 36, 1179–1190. [Google Scholar] [CrossRef] [Green Version]
- Fahn, A. Secretory Tissues in Plants; Academic Press: London, UK, 1979. [Google Scholar]
- Evert, R.F. Esau’s Plant Anatomy—Meristems, Cells, and Tissues of the Plant Body: Their Structure, Function, and Development, 3rd ed.; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2006. [Google Scholar]
- Rudall, P.J. Laticifers in Euphorbiaceae—A conspectus. Bot. J. Linn. Soc. 1987, 94, 143–163. [Google Scholar] [CrossRef]
- Marinho, C.R.; Teixeira, S.P. Novel reports of laticifers in Moraceae and Urticaceae: Revisiting synapomorphies. Plant Syst. Evol. 2019, 305, 13–31. [Google Scholar] [CrossRef]
- Demarco, D. Floral glands in asclepiads: Structure, diversity and evolution. Acta Bot. Bras. 2017, 31, 477–502. [Google Scholar] [CrossRef] [Green Version]
- Marin, M.; Koko, V.; Duletić-Laušević, S.; Marin, P.D.; Rančić, D.; Dajic-Stevanovic, Z. Glandular trichomes on the leaves of Rosmarinus officinalis: Morphology, stereology and histochemistry. S. Afr. J. Bot. 2006, 72, 378–382. [Google Scholar] [CrossRef] [Green Version]
- Ciccarelli, D.; Garbari, F.; Pagni, A.M. The flower of Myrtus communis (Myrtaceae): Secretory structures, unicellular papillae, and their ecological role. Flora 2008, 203, 85–93. [Google Scholar] [CrossRef]
- Hauenschild, F.; Matuszak, S.; Muellner-Riehl, A.N.; Favre, A. Phylogenetic relationships within the cosmopolitan buckthorn family (Rhamnaceae) support the resurrection of Sarcomphalus and the description of Pseudoziziphus gen. nov. TAXON 2016, 65, 47–64. [Google Scholar] [CrossRef]
- Thomas, V. Structural, functional and phylogenetic aspects of the colleter. Ann. Bot. 1991, 68, 287–305. [Google Scholar] [CrossRef]
- Machado, A.F.P.; Souza, A.M.; Leitão, C.A.E. Secretory structures at syconia and flowers of Ficus enormis (Moraceae): A specialization at ostiolar bracts and the first report of inflorescence colleters. Flora 2013, 208, 45–51. [Google Scholar] [CrossRef]
- Foster, A.S. Plant idioblasts: Remarkable examples of cell specialization. Protoplasma 1956, 46, 184–193. [Google Scholar] [CrossRef]
- Schultz, J.C. Tannin-insect interactions. In Chemistry and Significance of Condensed Tannins; Hemingway, R.W., Karehesy, J.J., Eds.; Plenum Press: New York, NY, USA, 1989; pp. 417–433. [Google Scholar]
- Peters, N.K.; Verma, D.P.S. Phenolic compounds as regulators of gene expression in plant-microbe interactions. Mol. Plant Microbe Interact. 1990, 3, 4–8. [Google Scholar] [CrossRef]
- Schlesinger, W.H. Biogeochemistry: An Analysis of Global Change; Academic Press: San Diego, CA, USA, 1991. [Google Scholar]
- Appel, H.M. Phenolics in ecological interactions: The importance of oxidation. J. Chem. Ecol. 1993, 19, 1521–1552. [Google Scholar] [CrossRef] [PubMed]
- Castro, M.M.; Demarco, D. Phenolic compounds produced by secretory structures in plants: A brief review. Nat. Prod. Commun. 2008, 3, 1273–1284. [Google Scholar]
- Leme, F.M.; Borella, P.H.; Marinho, C.R.; Teixeira, S.P. Expanding the laticifer knowledge in Cannabaceae: Distribution, morphology, origin, and latex composition. Protoplasma 2020, 257, 1183–1199. [Google Scholar] [CrossRef]
- Guignard, L.; Colin, H. Sur la présence de réservoirs a gomme chez les Rhamnées. Bull. Soc. Bot. France 1888, 35, 325–327. [Google Scholar] [CrossRef]
- Bouchet, P. Étude ultrastructurale des cellules constituant les poches “lysigènes” à mucilage de la bourdaine: Rhamnus frangula L. C. R. Hebd. Séances Acad. Sci. 1974, 279, 1073–1076. [Google Scholar]
- Pool, A. New species, combinations, and lectotypifications in Neotropical and northern Mexican Frangula (Rhamnaceae). Novon 2013, 22, 447–467. [Google Scholar] [CrossRef]
- Wollenweber, E.; Dörr, M.; Bohm, B.A.; Roitman, J.N. Exudate flavonoids of eight species of Ceanothus (Rhamnaceae). Z. Naturforsch. 2004, 59, 459–462. [Google Scholar] [CrossRef]
- Sun, M.; Naeem, R.; Su, J.-X.; Cao, Z.-Y.; Burleigh, J.G.; Soltis, P.S.; Soltis, D.E.; Chen, Z.-D. Phylogeny of the Rosidae: A dense taxon sampling analysis. J. Syst. Evol. 2016, 54, 363–391. [Google Scholar] [CrossRef] [Green Version]
- Duffey, S.S.; Stout, M.J. Antinutritive and toxic components of plant defense against insects. Arch. Insect Biochem. Physiol. 1996, 32, 3–37. [Google Scholar] [CrossRef]
- Romeo, J.T.; Saunders, J.A.; Barbosa, P. Phytochemical Diversity and Redundancy in Ecological Interactions; Plenum Press: New York, NY, USA, 1996. [Google Scholar]
- Agrawal, A.A.; Fishbein, M. Plant defense syndromes. Ecology 2006, 87, S132–S149. [Google Scholar] [CrossRef]
- POWO. Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. 2019. Available online: http://www.plantsoftheworldonline.org/ (accessed on 10 November 2020).
- Clark, G. Staining Procedures, 4th ed.; Williams & Wilkins: Baltimore, MD, USA, 1981. [Google Scholar]
- Gerrits, P.O.; Horobin, R.W. The Application of Glycol Methacrylate in Histotechnology; Some Fundamental Principles; Faculteit der Geneeskunde, Rijksuniversiteit Groningen: Groningen, The Netherlands, 1991. [Google Scholar]
- O’Brien, T.P.; Feder, N.; McCully, M.E. Polychromatic staining of plant cell walls by toluidine blue O. Protoplasma 1964, 59, 368–373. [Google Scholar] [CrossRef]
- Jensen, W.E. Botanical Histochemistry: Principles and Practice; W.H. Freeman & Co.: San Francisco, CA, USA, 1962. [Google Scholar]
- Pearse, A.G.E. Histochemistry: Theoretical and Applied; C. Livingstone: Edinburgh, UK, 1972. [Google Scholar]
- David, R.; Carde, J. Coloration différentielle des inclusions lipidiques et terpeniques des pseudophylles du pin maritime au moyen du reactif Nadi. C. R. Hebd. Seanc. Acad. Sci. Paris 1964, 258, 1338–1340. [Google Scholar]
- Karnovsky, M.J. A formaldehyde-glutaraldehyde fixative of high osmolarity for use in eletron microscopy. J. Cell Biol. 1965, 27, 137–138. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Colleter | Cavity | Duct | Idioblast |
|---|---|---|---|---|
| C. glandulosa | + (bract) | - | - | + (bract, pedicel, hypanthium, sepal, petal, filament, connective, anther, carpel) |
| G. virgata | + (bract) | - | - | + (bract, pedicel, hypanthium, sepal, petal, filament, connective, anther, carpel) |
| G. latifolia | + (bract) | - | - | + (bract, pedicel, hypanthium, sepal, petal, filament, connective, anther, carpel) |
| H. dulcis | + (bract) | - | - | + (bract, pedicel, hypanthium, sepal, petal, filament, connective, anther, carpel) |
| R. elaeocarpum | + (bract) | + (hypanthium, carpel) | + (pedicel, hypanthium) | + (bract, pedicel, hypanthium, sepal, petal, filament, connective, anther, carpel) |
| S. joazeiro | + (bract) | - | - | + (bract, pedicel, hypanthium, sepal, petal, filament, connective, anther, carpel) |
| Species | Clade | Tribe | Vouchers | Collection Sites |
|---|---|---|---|---|
| Colubrina glandulosa | Ziziphoid | Incertae sedis | SPFR 17155 | Ribeirão Preto, SP, Brazil (FORP 1-USP 2). |
| Gouania virgata | Ziziphoid | Gouanieae | SPFR 17156 | Ribeirão Preto, SP, Brazil (Santa Tereza Forest). |
| Gouania latifolia | Ziziphoid | Gouanieae | Cortez, 70 | Brasília-DF, Brazil (National Park of Brasília). |
| Hovenia dulcis | Ziziphoid | Paliureae | SPFR 17157 | Ribeirão Preto, SP, Brazil (CREU 3-USP). |
| Rhamnidium elaeocarpum | Rhamnoid | Rhamneae | SPFR 17158 | Ribeirão Preto, SP, Brazil (CEFER 4-USP). |
| Sarcomphalus joazeiro | Ziziphoid | Paliureae | RB 6544 | Rio de Janeiro, RJ, Brazil (JBRJ 5). |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ribeiro, C.; Marinho, C.; Teixeira, S. Uncovering the Neglected Floral Secretory Structures of Rhamnaceae and Their Functional and Systematic Significance. Plants 2021, 10, 736. https://doi.org/10.3390/plants10040736
Ribeiro C, Marinho C, Teixeira S. Uncovering the Neglected Floral Secretory Structures of Rhamnaceae and Their Functional and Systematic Significance. Plants. 2021; 10(4):736. https://doi.org/10.3390/plants10040736
Chicago/Turabian StyleRibeiro, Carimi, Cristina Marinho, and Simone Teixeira. 2021. "Uncovering the Neglected Floral Secretory Structures of Rhamnaceae and Their Functional and Systematic Significance" Plants 10, no. 4: 736. https://doi.org/10.3390/plants10040736
APA StyleRibeiro, C., Marinho, C., & Teixeira, S. (2021). Uncovering the Neglected Floral Secretory Structures of Rhamnaceae and Their Functional and Systematic Significance. Plants, 10(4), 736. https://doi.org/10.3390/plants10040736