
Kentucky Bluegrass Invasion in the Northern Great Plains and Prospective Management Approaches to Mitigate Its Spread


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Distribution and Ecophysiology of Kentucky Bluegrass
3. Multifactorial Impacts on Ecosystem Stability and Functioning
4. Drivers of the Successful Invasion of Kentucky Bluegrass
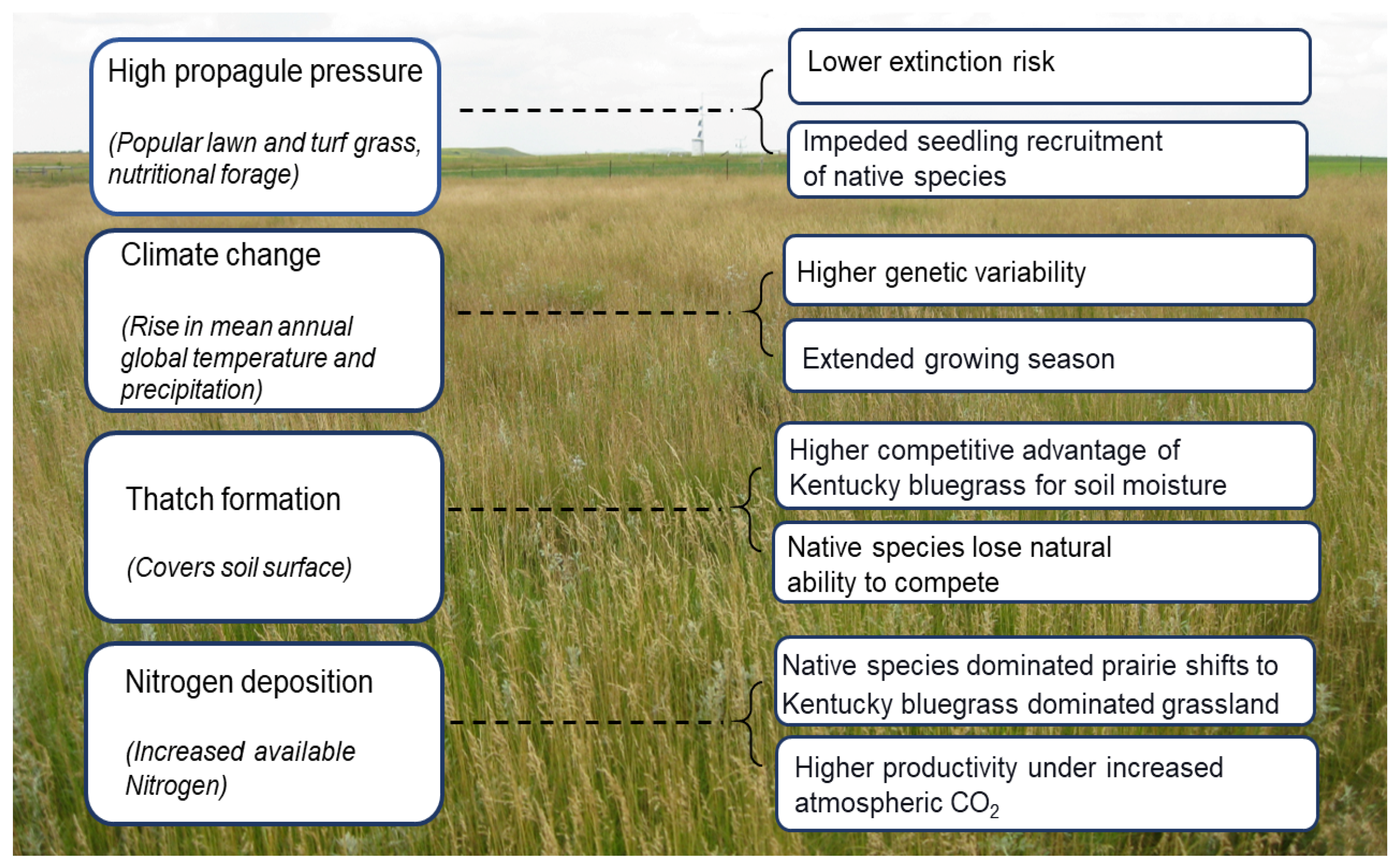
4.1. Propagule Pressure
4.2. Thatch Formation
4.3. Climate Change
4.4. Nitrogen Deposition
5. Requirement of Efficient Management Strategies
5.1. Modification of Existing Management Practices
5.2. Competition with Native Species as a Prospective Management Tool
6. Outlook
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Grant, T.A.; Shaffer, T.L.; Flanders, B. Resiliency of native prairies to invasion by Kentucky bluegrass, smooth brome, and woody vegetation. Rangel. Ecol. Manag. 2020, 73, 321–328. [Google Scholar] [CrossRef]
- Kobiela, B.; Quast, J.; Dixon, C.; DeKeyser, E.S. Targeting introduced species to improve plant community composition on USFWS-managed prairie remnants. Nat. Areas J. 2017, 37, 150–160. [Google Scholar] [CrossRef] [Green Version]
- DeKeyser, E.S.; Dennhardt, L.A.; Hendrickson, J. Kentucky bluegrass (Poa pratensis) Invasion in the Northern Great Plains: A story of rapid dominance in an endangered ecosystem. Invasive Plant Sci. Manag. 2015, 8, 255–261. [Google Scholar] [CrossRef] [Green Version]
- DeKeyser, S.; Clambey, G.; Krabbenhoft, K.; Ostendorf, J. Are changes in species composition on Central North Dakota Rangelands due to non-use management? Rangelands 2009, 31, 16–19. [Google Scholar] [CrossRef] [Green Version]
- Samson, F.; Knopf, F. Prairie Conservation in North America. Bioscience 1994, 44, 418–421. [Google Scholar] [CrossRef] [Green Version]
- Toledo, D.; Sanderson, M.; Spaeth, K.; Hendrickson, J.; Printz, J. Extent of Kentucky bluegrass and its effect on native plant species diversity and ecosystem Services in the Northern Great Plains of the United States. Invasive Plant Sci. Manag. 2014, 7, 543–552. [Google Scholar] [CrossRef]
- DiTomaso, J.M. Invasive weeds in rangelands: Species, impacts, and management. Weed Sci. 2000, 48, 255–265. [Google Scholar] [CrossRef] [Green Version]
- Mack, R.N. Invasion of Bromus tectorum L. into Western North America: An ecological chronicle. Agro-Ecosystems 1981, 7, 145–165. [Google Scholar] [CrossRef]
- Levine, J.M.; Vilà, M.; Antonio, C.M.D.; Dukes, J.S.; Grigulis, K.; Lavorel, S. Mechanisms underlying the impacts of exotic plant invasions. Proc. R. Soc. B Biol. Sci. 2003, 270, 775–781. [Google Scholar] [CrossRef]
- Wilcove, D.S.; Rothstein, D.; Dubow, J.; Phillips, A.; Losos, E. Quantifying threats to imperiled species in the United States. Bioscience 1998, 48, 607–615. [Google Scholar] [CrossRef] [Green Version]
- Samson, F.B.; Knopf, F.L.; Ostlie, W.R. Great Plains ecosystems: Past, present, and future. Wildl. Soc. Bull. 2004, 32, 6–15. [Google Scholar] [CrossRef]
- Hoekstra, J.M.; Boucher, T.M.; Ricketts, T.H.; Roberts, C. Confronting a biome crisis: Global disparities of habitat loss and protection. Ecol. Lett. 2004, 8, 23–29. [Google Scholar] [CrossRef]
- Hendrickson, J.R.; Sedivec, K.K.; Toledo, D.; Printz, J. Challenges facing grasslands in the Northern Great Plains and North Central Region. Rangelands 2019, 41, 23–29. [Google Scholar] [CrossRef]
- Grant, T.A.; Shaffer, T.L.; Flanders, B. Patterns of Smooth Brome, Kentucky bluegrass, and shrub invasion in the Northern Great Plains vary with temperature and precipitation. Nat. Areas J. 2020, 40, 11. [Google Scholar] [CrossRef]
- Pimentel, D.; McNair, S.; Janecka, J.; Wightman, J.; Simmonds, C.; O’Connell, C.; Wong, E.; Russel, L.; Zern, J.; Aquino, T.; et al. Economic and environmental threats of alien plant, animal, and microbe invasions. Agric. Ecosyst. Environ. 2001, 84, 1–20. [Google Scholar] [CrossRef]
- Fournier, A.; Penone, C.; Pennino, M.G.; Courchamp, F. Predicting future invaders and future invasions. Proc. Natl. Acad. Sci. USA 2019, 116, 7905–7910. [Google Scholar] [CrossRef] [Green Version]
- Stohlgren, T.J.; Schnase, J.L. Risk analysis for biological hazards: What we need to know about invasive species. Risk Anal. 2006, 26, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, D.; Zuniga, R.; Morrison, D. Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecol. Econ. 2005, 52, 273–288. [Google Scholar] [CrossRef]
- Sohrabi, S.; Downey, P.O.; Gherekhloo, J.; Hassanpour-Bourkheili, S. Testing the Australian post-border Weed Risk Management (WRM) system for invasive plants in Iran. J. Nat. Conserv. 2020, 53, 125780. [Google Scholar] [CrossRef]
- Ellis-Felege, S.N.; Dixon, C.S.; Wilson, S.D. Impacts and management of invasive cool-season grasses in the Northern Great Plains: Challenges and opportunities for wildlife. Wildl. Soc. Bull. 2013, 37, 510–516. [Google Scholar] [CrossRef]
- Great Plains Flora Association. Flora of the Great Plains; McGregor, R.L., Barkley, T.M., Eds.; University Press of Kansas: Lawrence, KS, USA, 1986. [Google Scholar]
- Schulte, J.R. The Fire Ecology of Kentucky Bluegrass (Poa pratensis). Master’s Thesis, North Dakota State University, Fargo, ND, USA, 2011. [Google Scholar]
- Huff, D.R.; Bara, J.M. Determining genetic origins of aberrant progeny from facultative apomictic Kentucky bluegrass using a combination of flow cytometry and silver-stained RAPD markers. Theor. Appl. Genet. 1993, 87, 201–208. [Google Scholar] [CrossRef]
- Mack, R.N.; Simberloff, D.; Lonsdale, W.M.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic invasions: Causes, epidemiology, global consequences, and control. Ecol. Appl. 2000, 10, 689. [Google Scholar] [CrossRef]
- DeKeyser, E.S.; Meehan, M.; Clambey, G.; Krabbenhoft, K. Cool Season invasive grasses in Northern Great Plains natural Areas. Nat. Areas J. 2013, 33, 81–90. [Google Scholar] [CrossRef]
- Cully, A.C.; Cully, J.F.; Hiebert, R.D. Invasion of exotic plant species in tallgrass prairie fragments. Conserv. Biol. 2003, 17, 990–998. [Google Scholar] [CrossRef]
- Estes, J.A.; Terborgh, J.; Brashares, J.S.; Power, M.E.; Berger, J.; Bond, W.J.; Carpenter, S.R.; Essington, T.E.; Holt, R.D.; Jackson, J.B.C.; et al. Trophic downgrading of planet earth. Science 2011, 333, 301–306. [Google Scholar] [CrossRef] [Green Version]
- Hobbs, R.J.; Higgs, E.; Harris, J.A. Novel ecosystems: Implications for conservation and restoration. Trends Ecol. Evol. 2009, 24, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Mckinney, M.L.; Lockwood, J.L. Taxonomic and ecological enhancement of homogenization. Tree 1999, 5347, 450–453. [Google Scholar]
- Gasch, C.K.; Toledo, D.; Kral-O’Brien, K.; Baldwin, C.; Bendel, C.; Fick, W.; Gerhard, L.; Harmon, J.; Hendrickson, J.; Hovick, T.; et al. Kentucky bluegrass invaded rangeland: Ecosystem implications and adaptive management approaches. Rangelands 2020, 42, 106–116. [Google Scholar] [CrossRef]
- Link, A.; Kobiela, B.; DeKeyser, S.; Huffington, M. Effectiveness of burning, herbicide, and seeding toward restoring rangelands in Southeastern North Dakota. Rangel. Ecol. Manag. 2017, 70, 599–603. [Google Scholar] [CrossRef]
- Ereth, C.B.; Hendrickson, J.R.; Kirby, D.; DeKeyser, E.S.; Sedivec, K.K.; West, M.S. Controlling Kentucky Bluegrass with herbicide and burning is influenced by invasion level. Invasive Plant Sci. Manag. 2017, 10, 80–89. [Google Scholar] [CrossRef]
- United States Department of Agriculture, Natural Resources Conservation Service. Plants Profile for Poa pratensis (Kentucky bluegrass). Available online: https://plants.usda.gov/core/profile?symbol=POPR (accessed on 5 February 2021).
- Huff, D.R. Kentucky Bluegrass. In Turfgrass Biology, Genetics, and Breeding; Casler, M., Duncan, R., Eds.; Wiley: Bokoben, NJ, USA, 2003; pp. 345–379. [Google Scholar]
- Carrier, L.; Bort, S.K. The history of Kentucky bluegrass and white clover in the United States. Agron. J. 1916, 8, 256–266. [Google Scholar] [CrossRef]
- Uchytil, R.J. Poa pratensis. In Fire Effects Information System; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer): Fort Collins, CO, USA, 1993; Available online: https://www.fs.fed.us/database/feis/plants/graminoid/poapra/all.html. (accessed on 5 February 2021).
- Sedivec, K.; Barker, W. Selected North. Dakota and Minnesota Range Plants EB-69; NDSU Extension Service: Fargo, ND, USA, 1997. [Google Scholar]
- Hurto, K.A.; Turgeon, A.J.; Spomer, L.A. Physical characteristics of thatch as a turfgrass growing medium 1. Agron. J. 1980, 72, 165–167. [Google Scholar] [CrossRef]
- Garrison, M.A.; Stier, J.C. Cool-Season turfgrass colony and seed survival in a restored prairie. Crop Sci. 2010, 50, 345–356. [Google Scholar] [CrossRef]
- Setter, C.M.; Lym, R.G. Change in leafy Spurge (Euphorbia esula) density and soil Seedbank Composition 10 Years following Release of Aphthona spp. Biological Control Agents. Invasive Plant Sci. Manag. 2013, 6, 147–160. [Google Scholar] [CrossRef] [Green Version]
- VanEss, B.M.; Wilson, S.D. Impact and management of crested wheatgrass (Agropyron cristatum) in the northern Great Plains. Can. J. Plant Sci. 2007, 87, 1023–1028. [Google Scholar] [CrossRef]
- Pritekel, C.; Whittemore-Olson, A.; Snow, N.; Moore, J.C. Impacts from invasive plant species and their control on the plant community and belowground ecosystem at Rocky Mountain National Park, USA. Appl. Soil Ecol. 2006, 32, 132–141. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Harold, M.A.; Jane, L.; Jerry, M.M. Human domination of earth’s ecosystems. Science 1997, 277, 494–499. [Google Scholar] [CrossRef] [Green Version]
- Theoharides, K.A.; Dukes, J.S. Plant invasion across space and time: Factors affecting nonindigenous species success during four stages of invasion. New Phytol. 2007, 176, 256–273. [Google Scholar] [CrossRef]
- Wedin, D.A. Species, Nitrogen, and grassland dynamics: The constraints of stuff. In Linking Species & Ecosystems; Springer: New York, NY, USA, 1995; pp. 253–262. [Google Scholar]
- Tilman, D. Secondary succession and the pattern of plant dominance along experimental nitrogen gradients. Ecol. Monogr. 1987, 57, 189–214. [Google Scholar] [CrossRef]
- Tatina, R. Ordination and description of prairie plant communities from the southern Missouri Coteau in South Dakota. Gt. Plains Res. 1994, 4, 147–162. [Google Scholar]
- Emery, S.M.; Gross, K.L. Dominant species identity, not community evenness, regulates invasion in experimental grassland plant communities. Ecology 2007, 88, 954–964. [Google Scholar] [CrossRef]
- Wedin, D.A.; Pastor, J. Nitrogen mineralization dynamics in grass monocultures. Oecologia 1993, 96, 186–192. [Google Scholar] [CrossRef] [PubMed]
- DeAngelis, D.L. Dynamics of Nutrient Cycling and Food Webs; Chapman & Hall: London, UK, 1992. [Google Scholar]
- Sanderson, M.A.; Johnson, H.; Liebig, M.A.; Hendrickson, J.R.; Duke, S.E. Kentucky bluegrass invasion alters soil carbon and vegetation structure on Northern mixed-Grass prairie of the United States. Invasive Plant Sci. Manag. 2017, 10, 9–16. [Google Scholar] [CrossRef]
- Mahaney, W.M.; Smemo, K.A.; Gross, K.L. Impacts of C4 grass introductions on soil carbon and nitrogen cycling in C3-dominated successional systems. Oecologia 2008, 157, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Piper, C.L.; Lamb, E.G.; Siciliano, S.D. Smooth brome changes gross soil nitrogen cycling processes during invasion of a rough fescue grassland. Plant. Ecol. 2015, 216, 235–246. [Google Scholar] [CrossRef]
- Spaeth, K.E.; Pierson, F.B.; Weltz, M.A.; Awang, J.B. Gradient analysis of infiltration and environmental variables as related to rangeland vegetation. Biomass 1996, 39, 67–77. [Google Scholar] [CrossRef]
- Lockwood, J.L.; Cassey, P.; Blackburn, T. The role of propagule pressure in explaining species invasions. Trends Ecol. Evol. 2005, 20, 223–228. [Google Scholar] [CrossRef]
- Duncan, R.P.; Blackburn, T.M.; Rossinelli, S.; Bacher, S. Quantifying invasion risk: The relationship between establishment probability and founding population size. Methods Ecol. Evol. 2014, 5, 1255–1263. [Google Scholar] [CrossRef] [Green Version]
- Dawson, W.; Burslem, D.F.R.P.; Hulme, P.E. Factors explaining alien plant invasion success in a tropical ecosystem differ at each stage of invasion. J. Ecol. 2009, 97, 657–665. [Google Scholar] [CrossRef]
- Kolar, C.S.; Lodge, D.M. Progress in invasion biology: Predicting invaders. Trends Ecol. Evol. 2001, 16, 199–204. [Google Scholar] [CrossRef]
- Lockwood, J.L.; Cassey, P.; Blackburn, T.M. The more you introduce the more you get: The role of colonization pressure and propagule pressure in invasion ecology. Divers. Distrib. 2009, 15, 904–910. [Google Scholar] [CrossRef]
- Barney, J.N. North American history of two invasive plant species: Phytogeographic distribution, dispersal vectors, and multiple introductions. Biol. Invasions 2006, 8, 703–717. [Google Scholar] [CrossRef]
- Hall, C.R.; Hodges, A.W.; Haydu, J.J. The economic impact of the green industry in the United States. HortTechnology 2006, 16, 345–353. [Google Scholar] [CrossRef]
- Honig, J.A.; Bonos, S.A.; Meyer, W.A. Isolation and characterization of 88 polymorphic microsatellite markers in Kentucky bluegrass (Poa pratensis L.). HortScience 2010, 45, 1759–1763. [Google Scholar] [CrossRef] [Green Version]
- Dennhardt, L.A.; DeKeyser, E.S.; Tennefos, S.A.; Travers, S.E. There is no evidence of geographical patterning among invasive Kentucky bluegrass (Poa pratensis) populations in the Northern Great Plains. Weed Sci. 2016, 64, 409–420. [Google Scholar] [CrossRef]
- Printz, J.L.; Hendrickson, J.R. Impacts of Kentucky bluegrass invasion (Poa pratensis L.) on ecological processes in the Northern Great Plains. Rangelands 2015, 37, 226–232. [Google Scholar] [CrossRef] [Green Version]
- Badh, A.; Akyuz, A.; Vocke, G.; Mullins, B. Impact of climate change on the growing seasons in select cities of North Dakota, United States of America. Int. J. Clim. Chang. Impacts Responses 2009, 1, 105–118. [Google Scholar] [CrossRef]
- Thirkell, T.J.; Pastok, D.; Field, K.J. Carbon for nutrient exchange between arbuscular mycorrhizal fungi and wheat varies according to cultivar and changes in atmospheric carbon dioxide concentration. Glob. Chang. Biol. 2019, 26, 1725–1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dusenge, M.E.; Duarte, A.G.; Way, D.A. Plant carbon metabolism and climate change: Elevated CO2 and temperature impacts on photosynthesis, photorespiration and respiration. New Phytol. 2018, 221, 32–49. [Google Scholar] [CrossRef] [Green Version]
- Meinshausen, M.; Smith, S.J.; Calvin, K.; Daniel, J.S.; Kainuma, M.L.T.; Lamarque, J.F.; Matsumoto, K.; Montzka, S.A.; Raper, S.C.B.; Riahi, K.; et al. The RCP greenhouse gas concentrations and their extensions from 1765 to 2300. Clim. Chang. 2011, 109, 213–241. [Google Scholar] [CrossRef] [Green Version]
- Ciais, P.; Sabine, C.; Bala, G. Carbon and other biogeochemical cycles. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.K., Eds.; Peter: Bern, Switzerland, 2013. [Google Scholar]
- He, H.; Kirkham, M.B.; Lawlor, D.J.; Kanemasu, E.T. Photosynthesis and water relations of big bluestem (C4) and Kentucky bluegrass (C3 ) under high concentration carbon dioxide. Trans. Kans. Acad. Sci. 1992, 95, 139–152. [Google Scholar] [CrossRef]
- Wand, S.J.E.; Midgley, G.F.; Jones, M.H.; Curtis, P.S. Responses of wild C4 and C3 grass (Poaceae) species to elevated atmospheric CO2 concentration: A meta-analytic test of current theories and perceptions. Glob. Chang. Biol. 1999, 5, 723–741. [Google Scholar] [CrossRef] [Green Version]
- Dennhardt, L.A. Evidence of Climate Niche Creation in the Northern Great Plains: The History of Invasion, Population Genetics, Competitive Effect, and Long-Term Trends of Poa Pratensis L.; North Dakota State University: Fargo, ND, USA, 2016. [Google Scholar]
- Regional Climate Centers–National Oceanic and Atmospheric Administration. High Plains Regional Climate Center. 2014. Available online: http://www.hprcc.unl.edu/data/historical/index.php?state5nd&action5se-lect_state&submit5Select+State (accessed on 5 February 2021).
- Stevens, O.A. Handbook of North Dakota Plants; North Dakota State University: Fargo, ND, USA, 1953. [Google Scholar]
- Lowe, E. A Natural History of British Grasses; Bell and Daldy: York, UK, 1858. [Google Scholar]
- Clark, J.S.; Grimm, E.C.; Donovan, J.J.; Fritz, S.C.; Engstrom, D.R.; Almendinger, J.E. Drought cycles and landscape responses to past aridity on prairies of the Northern Great Plains, USA. Ecology 2002, 83, 595–601. [Google Scholar] [CrossRef]
- Nie, D.; Kirkham, M.B.; Ballou, L.K.; Lawlor, D.J.; Kanemasu, E.T. Changes in prairie vegetation under elevated carbon dioxide levels and two soil moisture regimes. J. Veg. Sci. 1992, 3, 673–678. [Google Scholar] [CrossRef]
- Bobbink, R.; Hicks, K.; Galloway, J.; Spranger, T.; Alkemade, R.; Ashmore, M.; Bustamante, M.; Cinderby, S.; Davidson, E.; Dentener, F.; et al. Global assessment of nitrogen deposition effects on terrestrial plant diversity: A synthesis. Ecol. Appl. 2010, 20, 30–59. [Google Scholar] [CrossRef] [Green Version]
- Bobbink, R.; Hornung, M.; Roelofs, J.G.M. The effects of air-borne nitrogen pollutants on species diversity in natural and semi-natural European vegetation. J. Ecol. 1998, 86, 717–738. [Google Scholar] [CrossRef]
- Dentener, F.; Drevet, J.; Lamarque, J.F.; Bey, I.; Eickhout, B.; Fiore, A.M.; Hauglustaine, D.; Horowitz, L.W.; Krol, M.; Kulshrestha, U.C.; et al. Nitrogen and sulfur deposition on regional and global scales: A multimodel evaluation. Glob. Biogeochem. Cycles 2006, 20. [Google Scholar] [CrossRef]
- Phoenix, G.K.; Hicks, W.K.; Cinderby, S.; Kuylenstierna, J.C.I.; Stock, W.D.; Dentener, F.J.; Giller, K.E.; Austin, A.T.; Lefroy, R.D.B.; Gimeno, B.S.; et al. Atmospheric nitrogen deposition in world biodiversity hotspots: The need for a greater global perspective in assessing N deposition impacts. Glob. Chang. Biol. 2006, 12, 470–476. [Google Scholar] [CrossRef]
- Bowman, D.M.J.S.; Balch, J.K.; Artaxo, P.; Bond, W.J.; Carlson, J.M.; Cochrane, M.A.; D’Antonio, C.M.; DeFries, R.S.; Doyle, J.C.; Harrison, S.P.; et al. Fire in the Earth System. Science 2009, 324, 481–484. [Google Scholar] [CrossRef]
- Goergen, E.M.; Chambers, J.C. Influence of a native legume on soil N and plant response following prescribed fire in sagebrush steppe. Int. J. Wildland Fire 2009, 18, 665–675. [Google Scholar] [CrossRef] [Green Version]
- Certini, G. Effects of fire on properties of forest soils: A review. Oecologia 2005, 143, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hendrickson, J.R.; Wienhold, B.J.; Berdahl, J.D. Decomposition Rates of Native and Improved Cultivars of Grasses in the Northern Great Plains. Arid. Land Res. Manag. 2001, 15, 347–357. [Google Scholar] [CrossRef]
- Wedin, D.A.; Tilman, D. Species effects on nitrogen cycling: A test with perennial grasses. Oecologia 1990, 84, 433–441. [Google Scholar] [CrossRef]
- Brooks, M.L.; D’Antonio, C.M.; Richardson, D.M.; Grace, J.B.; Keeley, J.E.; DiTomaso, J.M.; Hobbs, R.J.; Pellant, M.; Pyke, D.A. Effects of Invasive Alien Plants on Fire Regimes. Bioscience 2004, 54, 677–688. [Google Scholar] [CrossRef] [Green Version]
- Bradley, B.A.; Htonw, R.A.H.G.; Mustard, J.F.; Hamburg, S.P. Invasive grass reduces aboveground carbon stocks in shrublands of the Western US. Glob. Chang. Biol. 2006, 12, 1815–1822. [Google Scholar] [CrossRef] [Green Version]
- Toledo, D.; Hendrickson, J.R.; Agricultural Research Services, United States, Department of Agriculture, Mandan, ND, USA. Relation of Soil Nitrogen Based on the Extent Kentucky Bluegrass (Poa pratensis L.) Invasion in the Northern Great Plains. Personal Communication, 2020. [Google Scholar]
- Grygiel, C.E.; Norland, J.E.; Biondini, M.E. Can carbon and phosphorous amendments increase native forbs in a restoration process? A case study in the Northern tall-grass rairie (U.S.A.). Restor. Ecol. 2010, 20, 122–130. [Google Scholar] [CrossRef]
- Grant, T.A.; Flanders-Wanner, B.; Shaffer, T.L.; Murphy, R.K.; Knutsen, G.A. An emerging crisis across Northern Prairie Refuges: Prevalence of invasive plants and a plan for adaptive management. Ecol. Restor. 2009, 27, 58–65. [Google Scholar] [CrossRef]
- Murphy, R.K.; Grant, T.A. Land management history and floristics in mixed-grass prairie, North Dakota, USA. Nat. Areas J. 2005, 25, 351–358. [Google Scholar]
- Kral, K.; Limb, R.; Ganguli, A.; Hovick, T.; Sedivec, K. Seasonal prescribed fire variation decreases inhibitory ability of Poa pratensis L. and promotes native plant diversity. J. Environ. Manag. 2018, 223, 908–916. [Google Scholar] [CrossRef]
- Mangla, S.; Sheley, R.L.; James, J.J.; Radosevich, S.R. Intra and interspecific competition among invasive and native species during early stages of plant growth. Plant. Ecol. 2011, 212, 531–542. [Google Scholar] [CrossRef]
- Brown, C.S.; Anderson, V.J.; Claassen, V.P.; Stannard, M.E.; Wilson, L.M.; Atkinson, S.Y.; Bromberg, J.E.; Grant, T.A.; Munis, M.D. Restoration ecology and invasive plants in the semiarid West. Invasive Plant Sci. Manag. 2008, 1, 399–413. [Google Scholar] [CrossRef]
- Dornbusch, M.J.; Limb, R.F.; Gasch, C.K. Facilitation of an exotic grass through nitrogen enrichment by an exotic legume. Rangel. Ecol. Manag. 2018, 71, 691–694. [Google Scholar] [CrossRef]
- Kirkman, L.K.; Barnett, A.; Williams, B.W.; Hiers, J.K.; Pokswinski, S.M.; Mitchell, R.J. A dynamic reference model: A framework for assessing biodiversity restoration goals in a fire-dependent ecosystem. Ecol. Appl. 2013, 23, 1574–1587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jauni, M.; Gripenberg, S.; Ramula, S. Non-native plant species benefit from disturbance: A meta-analysis. Oikos 2015, 124, 122–129. [Google Scholar] [CrossRef]
- Simmons, M.T.; Windhager, S.; Power, P.; Lott, J.; Lyons, R.K.; Schwope, C. Selective and non-selective control of invasive plants: The short-term effects of growing-season prescribed fire, herbicide, and mowing in two texas prairies. Restor. Ecol. 2007, 15, 662–669. [Google Scholar] [CrossRef]
- Lang, M.; Hanslin, H.M.; Kollmann, J.; Wagner, T. Suppression of an invasive legume by a native grass: High impact of priority effects. Basic Appl. Ecol. 2017, 22, 20–27. [Google Scholar] [CrossRef]
- Leskovšek, R.; Eler, K.; Batič, F.; Simončič, A. The influence of nitrogen, water and competition on the vegetative and reproductive growth of common ragweed (Ambrosia artemisiifolia L.). Plant. Ecol. 2012, 213, 769–781. [Google Scholar] [CrossRef]
- Hwang, B.C.; Lauenroth, W.K. Effect of nitrogen, water and neighbor density on the growth of Hesperis matronalis and two native perennials. Biol. Invasions 2007, 10, 771–779. [Google Scholar] [CrossRef]
- Huenneke, L.F.; Hamburg, S.P.; Koide, R.; Mooney, H.A.; Vitousek, P.M. Effects of soil resources on plant invasion and community structure in Californian Serpentine Grassland. Ecology 1990, 71, 478–491. [Google Scholar] [CrossRef]
- Limpens, J.; Berendse, F.; Klees, H. N deposition affects N availability in interstitial water, growth of Sphagnum and invasion of vascular plants in bog vegetation. New Phytol. 2003, 157, 339–347. [Google Scholar] [CrossRef]
- Lowe, P.N.; Lauenroth, W.K.; Burke, I.C. Effects of nitrogen availability on competition between Bromus tectorum and Bouteloua gracilis. Plant. Ecol. 2003, 167, 247–254. [Google Scholar] [CrossRef]
- Thomsen, M.A.; Corbin, J.D.; D’Antonio, C.M. The effect of soil nitrogen on competition between native and exotic perennial grasses from northern coastal California. Plant. Ecol. 2006, 186, 23–35. [Google Scholar] [CrossRef]
- Kolb, A.; Alpert, P. Effects of nitrogen and salinity on growth and competition between a native grass and an invasive congener. Biol. Invasions 2003, 5, 229–238. [Google Scholar] [CrossRef]
- Daehler, C.C. Performance comparisons of co-occurring native and alien invasive plants: Implications for conservation and restoration. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 183–211. [Google Scholar] [CrossRef]
- Fagúndez, J.; Lema, M. A competition experiment of an invasive alien grass and two native species: Are functionally similar species better competitors? Biol. Invasions 2019, 21, 3619–3631. [Google Scholar] [CrossRef]
- Funk, J.L.; Cleland, E.E.; Suding, K.N.; Zavaleta, E.S. Restoration through reassembly: Plant traits and invasion resistance. Trends Ecol. Evol. 2008, 23, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Yannelli, F.A.; Koch, C.; Jeschke, J.M.; Kollmann, J. Limiting similarity and Darwin’s naturalization hypothesis: Understanding the drivers of biotic resistance against invasive plant species. Oecologia 2017, 183, 775–784. [Google Scholar] [CrossRef]
- Goldberg, D.E.; Landa, K. Competitive effect and response: Hierarchies and correlated traits in the early stages of competition. J. Ecol. 1991, 79, 1013. [Google Scholar] [CrossRef] [Green Version]
- Stephens, M. Phenotypic Plasticity of Native and Invasive Cool-Season Grasses in Response to Frequency of Moisture Availability; South Dakota State University: Broolings, SD, USA, 2017. [Google Scholar]
- Dickson, T.L.; Hopwood, J.L.; Wilsey, B.J. Do priority effects benefit invasive plants more than native plants? An experiment with six grassland species. Biol. Invasions 2012, 14, 2617–2624. [Google Scholar] [CrossRef]
- Abraham, J.K.; Corbin, J.D.; D’Antonio, C.M. California native and exotic perennial grasses differ in their response to soil nitrogen, exotic annual grass density, and order of emergence. Herbaceous Plant Ecol. 2008, 2009, 81–92. [Google Scholar] [CrossRef]
- Wurst, S.; Ohgushi, T. Do plant and soil mediated legacy effects impact future biotic interactions? Funct. Ecol. 2015, 29, 1373–1382. [Google Scholar] [CrossRef]
- Bever, J.D.; Dickie, I.A.; Facelli, E.; Facelli, J.M.; Klironomos, J.; Moora, M.; Rillig, M.C.; Stock, W.D.; Tibbett, M.; Zobel, M. Rooting theories of plant community ecology in microbial interactions. Trends Ecol. Evol. 2010, 25, 468–478. [Google Scholar] [CrossRef] [Green Version]
- Bever, J.D. Soil community feedback and the coexistence of competitors: Conceptual frameworks and empirical tests. New Phytol. 2003, 157, 465–473. [Google Scholar] [CrossRef] [Green Version]
- Driscoll, D.A.; Strong, C. Covariation of soil nutrients drives occurrence of exotic and native plant species. J. Appl. Ecol. 2017, 55, 777–785. [Google Scholar] [CrossRef]
- Borer, E.T.; Seabloom, E.W.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Lind, E.M.; Adler, P.B.; Alberti, J.; Anderson, T.M.; Bakker, J.D.; et al. Herbivores and nutrients control grassland plant diversity via light limitation. Nat. Cell Biol. 2014, 508, 517–520. [Google Scholar] [CrossRef]
- Lai, H.R.; Mayfield, M.M.; Gay-Des-Combes, J.M.; Spiegelberger, T.; Dwyer, J.M. Distinct invasion strategies operating within a natural annual plant system. Ecol. Lett. 2015, 18, 336–346. [Google Scholar] [CrossRef]
- Wassen, M.J.; Venterink, H.O.; Lapshina, E.D.; Tanneberger, F. Endangered plants persist under phosphorus limitation. Nat. Cell Biol. 2005, 437, 547–550. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palit, R.; Gramig, G.; DeKeyser, E.S. Kentucky Bluegrass Invasion in the Northern Great Plains and Prospective Management Approaches to Mitigate Its Spread. Plants 2021, 10, 817. https://doi.org/10.3390/plants10040817
Palit R, Gramig G, DeKeyser ES. Kentucky Bluegrass Invasion in the Northern Great Plains and Prospective Management Approaches to Mitigate Its Spread. Plants. 2021; 10(4):817. https://doi.org/10.3390/plants10040817
Chicago/Turabian StylePalit, Rakhi, Greta Gramig, and Edward S. DeKeyser. 2021. "Kentucky Bluegrass Invasion in the Northern Great Plains and Prospective Management Approaches to Mitigate Its Spread" Plants 10, no. 4: 817. https://doi.org/10.3390/plants10040817
APA StylePalit, R., Gramig, G., & DeKeyser, E. S. (2021). Kentucky Bluegrass Invasion in the Northern Great Plains and Prospective Management Approaches to Mitigate Its Spread. Plants, 10(4), 817. https://doi.org/10.3390/plants10040817