
Influence of Extremely Low Temperatures of the Pole of Cold on the Lipid and Fatty-Acid Composition of Aerial Parts of the Horsetail Family (Equisetaceae)

 ,
,  , ,
, ,
Abstract
:1. Introduction
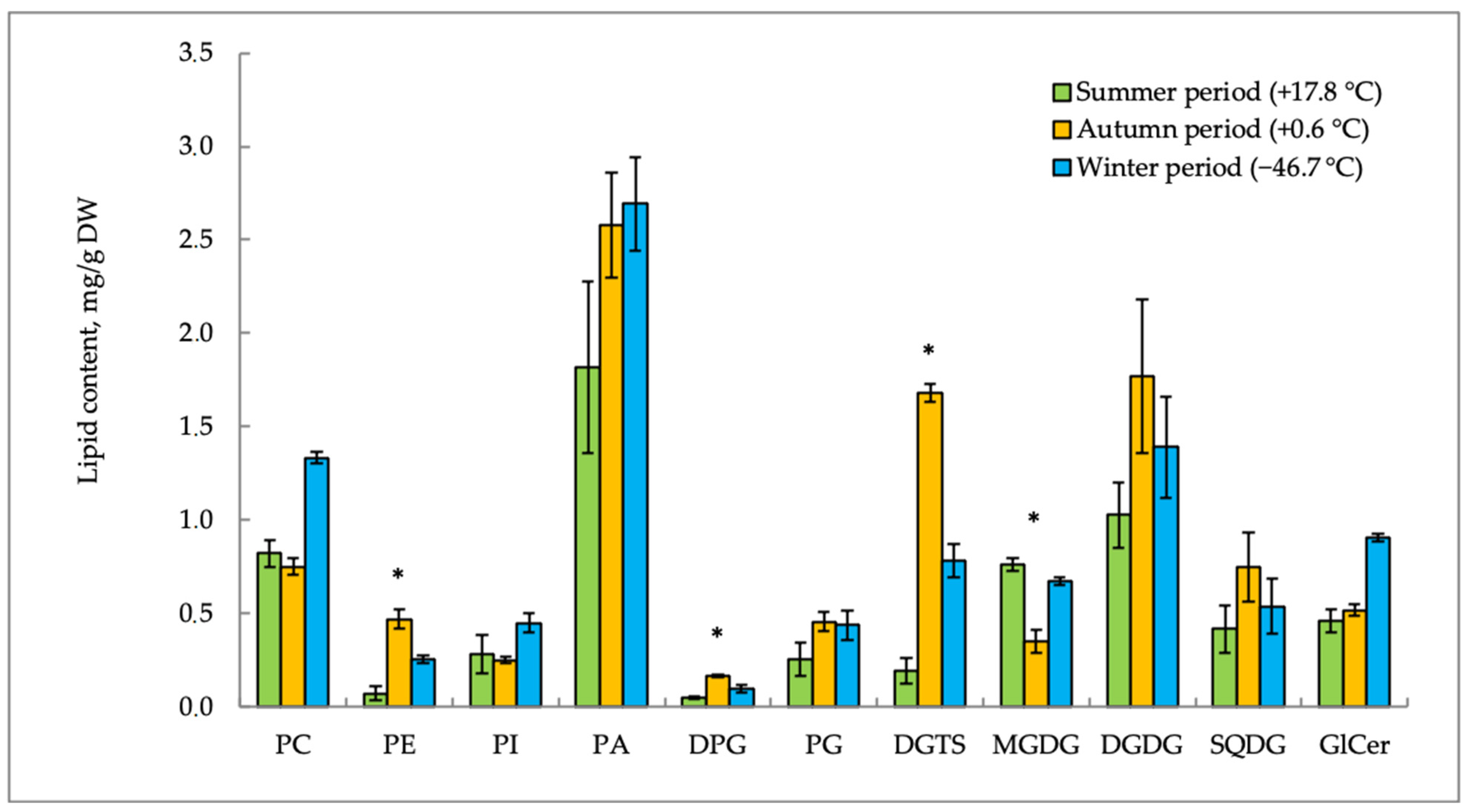
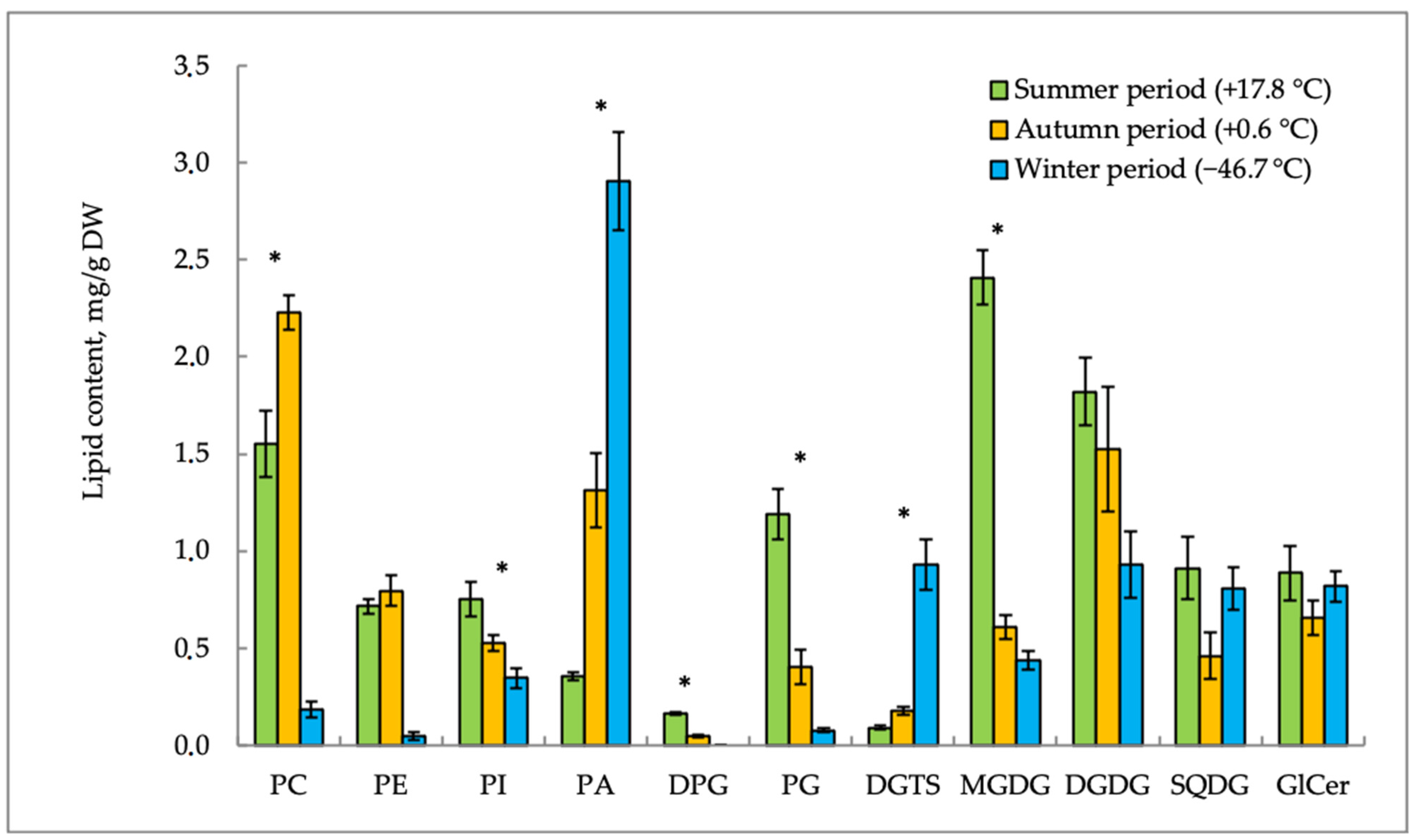
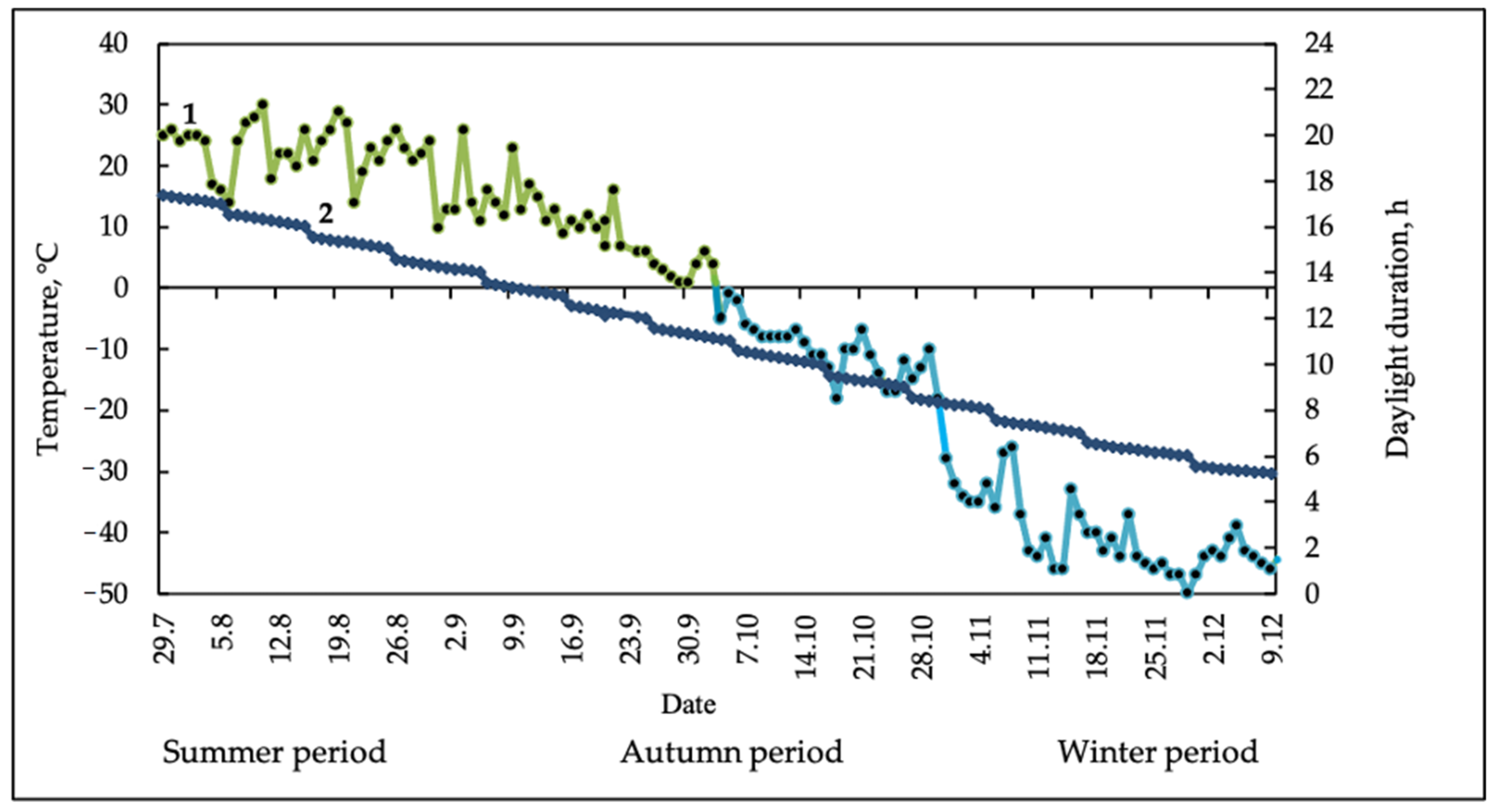
2. Results
2.1. Lipid Content
2.2. Fatty-Acid Composition
3. Discussion
4. Materials and Methods
DBI = ∑Pj nj/100,
ODR = (%C18:2 + %C18:3)/(%C18:1 + %C18:2 + %C18:3),
LDR = (%C18:3)/(%C18:2 + %C18:3).
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BLs | Betaine lipids |
| DBI | Double-bond index |
| DGDG | Digalactosyldiglyceride |
| DGTS | O-(1,2-diacylglycero)-N,N,N-trimethylhomoserine |
| DPG | Diphosphatidylglycerol |
| FA | Fatty acid |
| Gl | Glycolipids |
| GlCer | Glycoceramide |
| LDR | Linoleyl desaturase ratio |
| MGDG | Monogalactosyldiglyceride |
| ODR | Oleyl desaturase ratio |
| PA | Phosphatidic acid |
| PC | Phosphatidylcholine |
| PE | Phosphatidylethanolamine |
| PG | Phosphatidylglycerol |
| PI | Phosphatidylinositol |
| PLs | Phospholipids |
| SDR | Stearoyl desaturase ratio |
| SQDG | Sulfoquinovosyldiacylglycerol |
References
- Gabyshev, M.F. The Yakut Horse; Yakutsk Book Publishers: Yakutsk, USSR, 1957; p. 238. [Google Scholar]
- Egorov, A.D.; Potapov, V.Y.; Romanov, P.A. Zonal’no-Biokhimicheskie Osobennosti Kormovykh Rastenii Iakutii i Nekotorye Problemy Razvitiia Zhivotnovodstva [Zonal-Biochemical Characteristics of Fodder Plants in Yakutia and Some Problems in the Development of Animal Husbandry]; Yakutsk Book Publishing House: Yakutsk, Russia, 1962; p. 51. [Google Scholar]
- Petrov, K.A. Kriorezistentnost’ Rastenii: Ekologo-Fiziologicheskie i Biokhemicheskie Aspekty [Cryoresistance of Plants: Ecological, Physiological and Biochemical Aspects]; SB RAS Publishing House: Novosibirsk, Russia, 2016; p. 276. [Google Scholar]
- Petrov, K.A.; Dudareva, L.V.; Nokhsorov, V.V.; Stoyanov, K.N.; Makhutova, O.N. Fatty Acid Content and Composition of the Yakutian Horses and Their Main Food Source: Living in Extreme Winter Conditions. Biomolecules 2020, 10, 315. [Google Scholar] [CrossRef] [Green Version]
- Dudareva, L.V.; Nokhsorov, V.V.; Rudikovskaya, E.G.; Petrov, K.A. Fatty-acid profiles of aerial parts of three horsetail species growing in Central and Northern Yakutia. Chem. Nat. Compd. 2015, 51, 220–223. [Google Scholar] [CrossRef]
- Petrov, K.A.; Sofronova, V.E.; Chepalov, V.A.; Perk, A.A.; Maksimov, T.K. Seasonal Changes in the Content of Photosynthetic Pigments in Perennial Grasses of Cryolithic Zone. Russ. J. Plant Physiol. 2010, 57, 181–188. [Google Scholar] [CrossRef]
- Thomashow, M.F. Plant cold acclimation: Freezing tolerance genes and regulatory mechanisms. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 571–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouellet, F.; Charron, J.-B. Cold acclimation and freezing tolerance in plants. In Els; Wiley: Hoboken, NJ, USA, 2013. [Google Scholar]
- Hazel, J.R. Thermal adaptation in biological membranes: Is homeoviscous adaptation the explanation? Annu. Rev. Physiol. 1995, 57, 19–42. [Google Scholar] [CrossRef] [PubMed]
- Lyons, J.M. Chilling injury in plants. Ann. Rev. Plant Physiol. 1973, 24, 445–466. [Google Scholar] [CrossRef]
- Trunova, T.I. Rastenie i Nizkotemperaturnyi Stress [The Plant and the Stress Caused by Low Temperatures]; Nauka: Moscow, Russia, 2007; pp. 1–54. [Google Scholar]
- Los, D.A. Desaturazy Zhirnykh Kislot [Fatty Acid Desaturases]; Nauchnyj Mir: Moscow, Russia, 2014; 486p. [Google Scholar]
- Joyard, J.; Marechal, E.; Miege Ch Block, M.A.; Dorne, A.-J.; Douce, R. Structure, distribution and biosynthesis of glycerolipids from higher plant chloroplasts. In Lipids in Photosynthesis: Structure, Function and Genetics; Siegenthaler, P.-A., Murata, N., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1998; Volume 6, pp. 22–46. [Google Scholar]
- Wolff, R.L.; Christie, W.W.; Pedrono, F.; Marpeau, A.M.; Tsevegsüren, N.; Aitzetmüller, K.; Gunstone, F.D. Δ5-olefinic acids in the seed lipids from four Ephedra species and their distribution between the α and β positions of triacylglycerols. Characteristics common to coniferophytes and cycadophytes. Lipids 1999, 58, 101–115. [Google Scholar] [CrossRef]
- Testerink, C.; Munnik, T. Molecular, cellular and physiological responses to phosphatidic acid formation in plants. J. Exp. Bot. 2011, 62, 2349–2361. [Google Scholar] [CrossRef] [Green Version]
- Simidjiev, I.; Stoylova, S.; Amenitsch, H.; Javorfi, T.; Mustardy, L.; Laggner, P.; Holzenburg, A.; Garab, G. Self-assembly of large, ordered lamellae from non-bilayer lipids and integral membrane proteins in vitro. Proc. Natl. Acad. Sci. USA 2000, 97, 1473–1476. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.G. Membrane lipids: It’s only a phase. Curr. Biol. 2000, 10, 377–380. [Google Scholar] [CrossRef] [Green Version]
- Seiwert, D.; Witt, H.; Ritz, S.; Janshoff, A.; Paulsen, H. The nonbilayer lipid MGDG and the major light-harvesting complex (LHCII) promote membrane stacking in supported lipid bilayers. Biochemistry 2018, 57, 2278–2288. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.W.; Lin, Y.T.; Li, H.M. Increased ratio of galactolipid MGDG: DGDG induces jasmonic acid overproduction and changes chloroplast shape. New Phytol. 2020, 228, 1327–1335. [Google Scholar] [CrossRef] [PubMed]
- Rozentsvet, O.A.; Nesterov, V.N.; Bogdanova, E.S.; Tabalenkova, G.N.; Zakhozhiy, I.G. Biochemical conditionality of differentiation of halophytes by the type of regulation of salt metabolism in Prieltonye. Contemp. Prob. Ecol. 2016, 9, 98–106. [Google Scholar] [CrossRef]
- Li, H.M.; Yu, C.W. Chloroplast Galactolipids: The Link Between Photosynthesis, Chloroplast Shape, Jasmonates, Phosphate Starvation and Freezing Tolerance. Plant Cell Physiol. 2018, 59, 1128–1134. [Google Scholar] [CrossRef] [Green Version]
- Arisz, S.A.; Wijk, R.; Roels, W.; Zhu, J.K.; Haring, M.A.; Munnik, T. Rapid phosphatidic acid accumulation in response to low temperature stress in Arabidopsis is generated through diacylglycerol kinase. Front. Plant Sci. 2013, 4, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruelland, E.; Cantrel, C.; Gawer, M.; Kader, J.C.; Zachowski, A. Activation of phospholipases C and D is an early response to a cold exposure in Arabidopsis suspension cells. Plant Physiol. 2002, 130, 999–1007. [Google Scholar] [CrossRef] [Green Version]
- Senik, S.V.; Maloshenok, L.G.; Kotlova, E.R.; Shavarda, A.L.; Moiseenko, K.V.; Bruskin, S.A.; Koroleva, O.V.; Psurtseva, N.V. Diacylglyceryltrimethylhomoserine Content and Gene Expression Changes Triggered by Phosphate Deprivation in the Mycelium of the Basidiomycete Flammulina velutipes. Phytochemistry 2015, 117, 34–42. [Google Scholar] [CrossRef]
- Rozentsvet, O.A. Distribution of a Betaine Lipid O-(1,2-Diacylglycero)-4′-(N,N,N-Trimethyl) homoserine in Tissues of Some Lycopodiophyta Species. Russ. J. Bioorganic Chem. 2004, 30, 582–585. [Google Scholar] [CrossRef]
- Abida, H.; Dolch, L.-J.; Mei, C.; Villanova, V.; Conte, M.; Block, M.A.; Finazzi, G.; Bastien, O.; Tirichine, L.; Bowler, C.; et al. Membrane Glycerolipid Remodeling Triggered by Nitrogen and Phosphorus Starvation in Phaeodactylum tricornutum. Plant Physiol. 2015, 167, 118–136. [Google Scholar] [CrossRef] [Green Version]
- Benning, C.; Huang, Z.H.; Gage, D.A. Accumulation of a novel glycolipid and a betaine lipid in cell of Rhodobacter sphaeroides grown under phosphate limitation. Arch. Biochem. Biophys 1995, 317, 103–111. [Google Scholar] [CrossRef]
- Rozentsvet, O.A.; Bogdanova, E.S.; Tabalenkova, G.N.; Golovko, T.K.; Zakhozhiy, I.K. Ecological and biochemical properties of representatives of higher spore plants of South Timan. Povolzhskiy Ecol. J. 2011, 4, 489–498. [Google Scholar]
- Shemi, A.; Schatz, D.; Fredricks, H.F.; Van Mooy, B.A.-S.; Porat, Z.; Vardi, A. Phosphorus starvation induces membrane remodeling and recycling in Emiliania huxleyi. New Phytol. 2016, 211, 886–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozentsvet, O.A.; Filin, V.R.; Saksonov, S.V.; Meshcheryakov, V.V. Seasonal Changes in Polar Lipids in Fronds of the Ferns Dryopteris filixmas and Matteuccia struthiopteris. Biochemistry (Moscow) 2002, 9, 1006–1011. [Google Scholar] [CrossRef]
- Loll, B.; Kern, J.; Saenger, W.; Zouni, A.; Biesiadka, J. Lipids in photosystem II: Interactions with protein and cofactors. Biochim. Biophys. Acta 2007, 1767, 509–519. [Google Scholar] [CrossRef] [Green Version]
- Zhukov, A.V.; Kuznetsova, E.I.; Sidorov, R.A.; Pchelkin, V.P.; Tsydendambaev, V.D. Fatty acid composition of lipids from leaves and strobila of Cycas revoluta. Russ. J. Plant Physiol. 2018, 65, 23–29. [Google Scholar] [CrossRef]
- Nokhsorov, V.V.; Dudareva, L.V.; Petrov, K.A. Content and Composition of Lipids and Their Fatty Acids in Needles of Pinus sylvestris L. and Picea obovata Ledeb. upon Cold Hardening in the Cryolithozone of Yakutia. Russ. J. Plant Physiol. 2019, 66, 548–555. [Google Scholar] [CrossRef]
- Shanab, S.; Hafez, R.M.; Fouad, A.S. A review on algae and plants as potential source of arachidonic acid. J. Adv. Res. 2018, 11, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Kalberer, S.R.; Wisniewski, M.; Arora, R. Deacclimation and reacclimation of cold-hardy plants: Current understanding and emerging concepts. Plant Sci. 2006, 171, 3–16. [Google Scholar] [CrossRef]
- Christie, W.W. Preparation of ester derivatives of fatty acids for chromatographic analysis. Adv. Lipid Methodol. 1993, 2, 69–111. [Google Scholar]
- Christie, W.W. The AOCS Lipid Library: Methyl Esters of Fatty Acids—Archive of Mass Spectra. Available online: http://lipidlibrary.aocs.org/ms/arch_me/index.htm (accessed on 1 December 2010).
- Lyons, J.M.; Wheaton, T.A.; Pratt, H.R. Relationship between the physical nature of mitochondrial membranes and chilling sensitivity in plants. Plant Physiol. 1964, 39, 262–268. [Google Scholar] [CrossRef] [Green Version]
- Jaworski, J.G.; Stumpf, P.K. Fat metabolism in higher plants. Properties of a soluble stearyl-acyl carrier protein desaturase from maturing Carthamus tinctorius. Arch. Biochem. Biophys. 1974, 162, 158–165. [Google Scholar] [CrossRef]
- Kates, M. Techniques of Lipidology: Isolation, Analysis and Identification of Lipids; North-Holland: Amsterdam, The Netherlands, 1972; pp. 269–610. [Google Scholar]
- Potapov, V.Y. Uglevody i Lignin v Kormovyh Travah Iakutii [Carbohydrates and Lignin in Forage Grasses of Yakuia]; Nauka: Moscow, Russia, 1967; p. 174. [Google Scholar]
- Petrov, K.A.; Makhutova, O.N.; Gladyshev, M.I. Fatty Acid Composition of Yakut Horse Tissues. Dokl. Biochem. Biophys. 2020, 492, 105–107. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acids | Summer Period (+17.8 °C *) | Autumn Period (+0.6 °C *) | Winter Period (−46.7 °C *) | |||
|---|---|---|---|---|---|---|
| mg/g DW | % | mg/g DW | % | mg/g DW | % | |
| C12:0 | traces | 0.2 ± 0.0 a | – | – | – | – |
| C14:0 | 0.1 ± 0.0 a | 1.2 ± 0.4 a | 0.1 ± 0.0 b | 0.9 ± 0.1 a | 0.2 ± 0.0 b | 0.9 ± 0.2 a |
| C15:0 | traces | 0.4 ± 0.0 a | 0.1 ± 0.0 a | 0.5 ± 0.2 a | traces | 0.2 ± 0.0 b |
| C16:0 | 2.9 ± 0.7 a | 24.7 ± 1.1 a | 4.2 ± 0.7 a | 26.7 ± 2.1 c | 6.5 ± 1.2 b | 30.6 ± 1.3 c |
| C16:1 | 0.3 ± 0.1 a | 2.4 ± 0.8 b | 0.2 ± 0.0 a | 1 ± 0.2 a | 0.1 ± 0.0 a | 0.6 ± 0.1 a |
| C16:2(n-6) | 0.1 ± 0.0 b | 1 ± 0.3 a | 0.1 ± 0.0 b | 0.7 ± 0.1 b | 0.1 ± 0.0 a | 0.7 ± 0.1 a |
| C16:3(Δ7,11,14) | 0.7 ± 0.2 a | 6.2 ± 1.2 b | 0.2 ± 0.0 a | 1.3 ± 0.3 a | 0.8 ± 0.1 a | 4 ± 0.9 b |
| C17:0 | traces | 0.3 ± 0.0 | 0.1 ± 0.0 b | 0.6 ± 0.1 a | 0.1 ± 0.0 b | 0.5 ± 0.1 a |
| C18:0 | 0.2 ± 0.0 b | 2.1 ± 0.9 a | 0.5 ± 0.1 a | 3.3 ± 0.8 b | 0.5 ± 0.1 a | 2.3 ± 0.8 b |
| C18:1(n-9) | 0.6 ± 0.1 a | 5.2 ± 1.2 b | 0.9 ± 0.3 a | 5.5 ± 1.4 c | 1 ± 0.2 a | 4.6 ± 0.9 c |
| C18:1(n-7) | 0.1 ± 0.0 b | 0.5 ± 0.1 a | – | – | 0.1 ± 0.0 a | 0.5 ± 0.0 a |
| C18:2(n-6) | 1.3 ± 0.3 b | 11.3 ± 1.2 b | 3.7 ± 1.2 b | 23.3 ± 1.5 c | 3.2 ± 0.7 b | 15.2 ± 1.3 c |
| C18:3(n-3) | 4.6 ± 0.9 a | 39.5 ± 2.2 b | 4.1 ± 1.1 b | 25.7 ± 2.2 c | 6.6 ± 1.3 b | 31.5 ± 2.3 c |
| C20:0 | traces | 0.2 ± 0.0 a | 0.1 ± 0.0 a | 0.9 ± 0.1 a | 0.2 ± 0.0 a | 0.8 ± 0.1 a |
| C20:1(n-9) | traces | 0.3 ± 0.0 a | 0.2 ± 0.0 a | 1.2 ± 0.3 b | 0.1 ± 0.0 a | 0.6 ± 0.0 a |
| C20:3(Δ11,14,17) | 0.2 ± 0.0 a | 1.9 ± 0.9 b | 0.9 ± 0.1 a | 6 ± 1.3 c | 0.8 ± 0.2 a | 3.6 ± 1.1 b |
| C20:4(Δ5,11,14,17) | 0.3 ± 0.0 a | 2.4 ± 0.8 a | 0.4 ± 0.0 | 2.5 ± 0.9 b | 0.6 ± 0.1 a | 3 ± 0.9 b |
| C22:0 | traces | 0.3 ± 0.0 a | – | – | 0.1 ± 0.0 a | 0.4 ± 0.0 a |
| Total FAs | 11.6 ± 1.1 b | 100 | 15.8 ± 1.2 a | 100 | 21.1 ± 1.6 b | 100 |
| Σsaturated | 3.4 ± 0.8 a | 29.4 ± 2.1 c | 5.2 ± 0.7 a | 32.9 ± 2.2 c | 7.5 ± 0.8 a | 35.7 ± 1.9 c |
| Σunsaturated | 8.2 ± 0.9 b | 70.7 ± 1.9 c | 10.6 ± 1.1 b | 67.2 ± 2.4 b | 13.6 ± 1.1 c | 64.3 ± 2.1 c |
| k | 2.4 | 2 | 1.8 | |||
| DBI | 1.9 | 1.6 | 1.7 | |||
| SDR | 0.7 | 0.6 | 0.7 | |||
| ODR | 0.9 | 0.9 | 0.9 | |||
| LDR | 0.8 | 0.5 | 0.7 | |||
| Summer Period (+17.8 °C *) | Autumn Period (+0.6 °C *) | Winter Period (−46.7 °C *) | ||||
|---|---|---|---|---|---|---|
| Fatty Acids | ||||||
| mg/g DW | % | mg/g DW | % | mg/g DW | % | |
| C12:0 | traces | 0.3 ± 0.0 a | traces | 0.2 ± 0.0 a | – | – |
| C14:0 | 0.2 ± 0.0 a | 1.3 ± 0.2 a | 0.1 ± 0.0 a | 0.8 ± 0.1 a | 0.1 ± 0.0 a | 0.7 ± 0.1 a |
| C15:0 | traces | 0.3 ± 0.0 a | 0.1 ± 0.0 a | 0.3 ± 0.0 b | – | – |
| C16:0 | 2.7 ± 0.9 a | 21.1 ± 1.2 c | 3.9 ± 0.9 a | 22.6 ± 1.6 c | 5.9 ± 1.1 b | 31.3 ± 2.3 c |
| C16:1 | 0.1 ± 0.0 a | 1 ± 0.2 a | 0.2 ± 0.0 a | 1.4 ± 0.6 a | traces | 0.2 ± 0.0 a |
| C16:2(n-6) | 0.1 ± 0.0 a | 0.7 ± 0.1 a | 0.2 ± 0.0 c | 1.2 ± 0.5 a | – | – |
| C16:3(Δ7,11,14) | 0.3 ± 0.0 a | 2.6 ± 0.9 b | 1.0 ± 0.2 a | 5.7 ± 1.1 c | 1 ± 0.4 a | 5.2 ± 1.4 b |
| C17:0 | 0.1 ± 0.0 a | 0.7 ± 0.1 a | 0.1 ± 0.0 b | 0.3 ± 0.0 a | 0.1 ± 0.0 a | 0.6 ± 0.2 a |
| C18:0 | 0.7 ± 0.1 a | 5.2 ± 1.1 c | 0.3 ± 0.0 a | 1.8 ± 0.3 a | 0.3 ± 0.0 a | 1.8 ± 0.4 a |
| C18:1(n-9) | 0.8 ± 0.1 a | 6.1 ± 1.4 c | 0.6 ± 0.0 a | 3.7 ± 1.3 c | 0.9 ± 0.2 b | 4.8 ± 1.1 c |
| C18:1(n-7) | 0.1 ± 0.0 a | 0.6 ± 0.1 a | 0.1 ± 0.0 a | 0.3 ± 0.0 a | 0.1 ± 0.0 a | 0.3 ± 0.0 a |
| C18:2 (Δ5,9) | 0.7 ± 0.2 a | 5.8 ± 0.9 b | 0.5 ± 0.1 a | 3.1 ± 0.8 a | 0.5 ± 0.0 a | 2.4 ± 0.6 a |
| C18:2(n-6) | 2.0 ± 0.4 b | 16.1 ± 1.4 c | 2.3 ± 0.6 b | 13.1 ± 1.1 c | 3.6 ± 0.8 b | 18.8 ± 1.4 c |
| C18:3(n-3) | 4.0 ± 0.9 c | 31.4 ± 1.6 c | 6.5 ± 1.0 c | 37.5 ± 2.7 c | 5.9 ± 1.1 c | 31 ± 1.9 c |
| C20:0 | 0.1 ± 0.0 b | 0.8 ± 0.1 a | traces | 0.2 ± 0.0 b | 0.1 ± 0.0 a | 0.5 ± 0.0 a |
| C20:1(n-9) | traces | 0.1 ± 0.0 a | traces | 0.1 ± 0.0 b | – | – |
| C20:3(Δ11,14,17) | 0.4 ± 0.1 a | 2.8 ± 0.7 b | 0.6 ± 0.1 a | 3.7 ± 1.1 c | traces | 0.2 ± 0.0 a |
| C20:4(Δ5,11,14,17) | 0.3 ± 0.0 b | 2.2 ± 0.6 b | 0.6 ± 0.1 a | 3.4 ± 1.3 b | 0.3 ± 0.0 a | 1.7 ± 0.3 a |
| C22:0 | 0.1 ± 0.0 a | 0.8 ± 0.2 a | 0.1 ± 0.0 a | 0.5 ± 0.1 a | 0.1 ± 0.0 a | 0.5 ± 0.1 a |
| Total FAs | 12.6 ± 1.2 a | 100 | 17.2 ±1.4 c | 100 | 18.9 ± 1.2 b | 100 |
| Σsaturated | 3.8 ± 0.8 b | 30.5 ± 1.3 c | 4.6 ± 0.9 b | 26.7 ± 1.9 a | 6.7 ± 1.2 b | 35.4 ± 3.2 c |
| Σunsaturated | 8.7 ± 0.9 b | 69.4 ± 1.9 c | 12.6 ± 1.3 c | 73.2 ± 2.2 c | 12.2 ± 2.3 c | 64.6 ± 3.4 c |
| k | 2.3 | 2.7 | 1.8 | |||
| DBI | 1.8 | 1.8 | 1.6 | |||
| SDR | 0.7 | 0.6 | 0.7 | |||
| ODR | 0.9 | 0.9 | 0.9 | |||
| LDR | 0.7 | 0.7 | 0.6 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nokhsorov, V.V.; Dudareva, L.V.; Senik, S.V.; Chirikova, N.K.; Petrov, K.A. Influence of Extremely Low Temperatures of the Pole of Cold on the Lipid and Fatty-Acid Composition of Aerial Parts of the Horsetail Family (Equisetaceae). Plants 2021, 10, 996. https://doi.org/10.3390/plants10050996
Nokhsorov VV, Dudareva LV, Senik SV, Chirikova NK, Petrov KA. Influence of Extremely Low Temperatures of the Pole of Cold on the Lipid and Fatty-Acid Composition of Aerial Parts of the Horsetail Family (Equisetaceae). Plants. 2021; 10(5):996. https://doi.org/10.3390/plants10050996
Chicago/Turabian StyleNokhsorov, Vasiliy V., Lyubov V. Dudareva, Svetlana V. Senik, Nadezhda K. Chirikova, and Klim A. Petrov. 2021. "Influence of Extremely Low Temperatures of the Pole of Cold on the Lipid and Fatty-Acid Composition of Aerial Parts of the Horsetail Family (Equisetaceae)" Plants 10, no. 5: 996. https://doi.org/10.3390/plants10050996
APA StyleNokhsorov, V. V., Dudareva, L. V., Senik, S. V., Chirikova, N. K., & Petrov, K. A. (2021). Influence of Extremely Low Temperatures of the Pole of Cold on the Lipid and Fatty-Acid Composition of Aerial Parts of the Horsetail Family (Equisetaceae). Plants, 10(5), 996. https://doi.org/10.3390/plants10050996