
Application of Trehalose and Salicylic Acid Mitigates Drought Stress in Sweet Basil and Improves Plant Growth

 ,
,  ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Conditions
2.2. Leaf Characteristics
2.3. Leaf Total Phenolic Concentration
2.4. Electrolyte Leakage
2.5. Osmolyte Quantification
2.6. Determination of Hydrogen Peroxide
2.7. Determination of Malondialdehyde
2.8. Antioxidant Enzyme Extraction and Assay
2.9. Plant Vegetative Characteristics
2.10. Statistical Analyses
3. Results
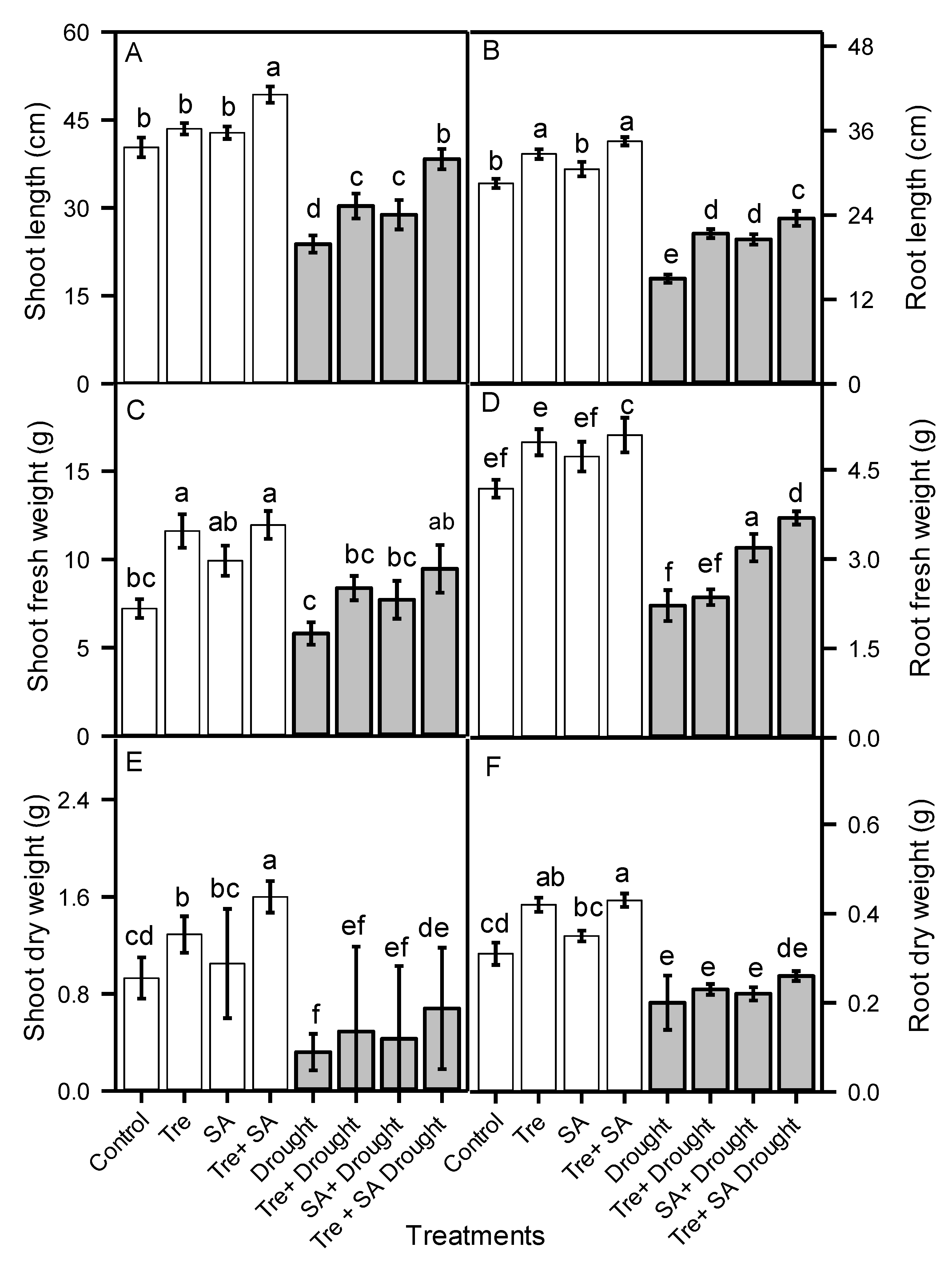
3.1. Plant Growth Responses
3.2. Leaf Chlorophyll Concentration and Leaf Gas Exchange
3.3. Chlorophyll Fluorescence
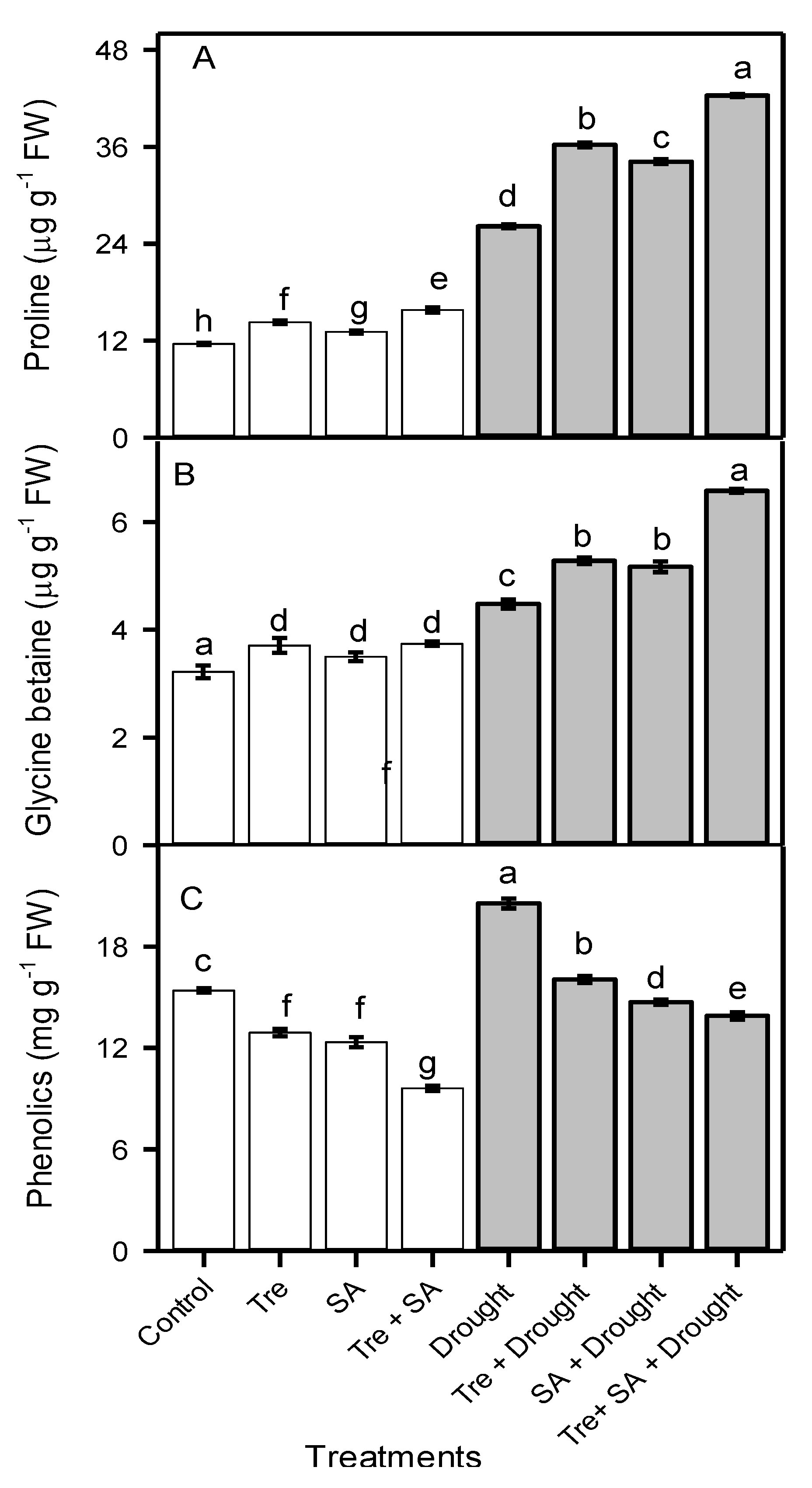
3.4. Leaf Free Proline and Glycine Betaine Concentrations
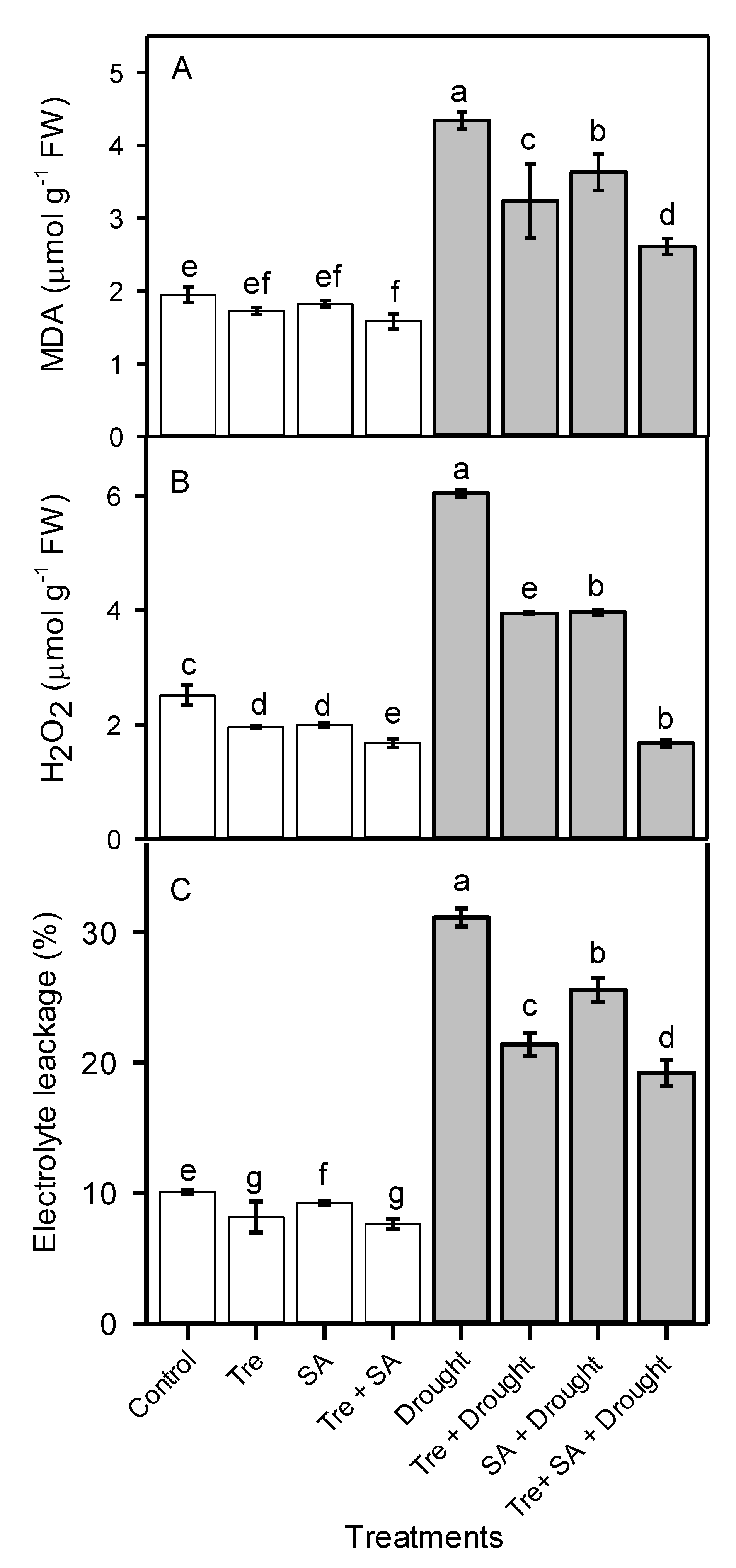
3.5. Electrolyte Leakage, and Hydrogen Peroxide and Malondialdehyde Concentrations
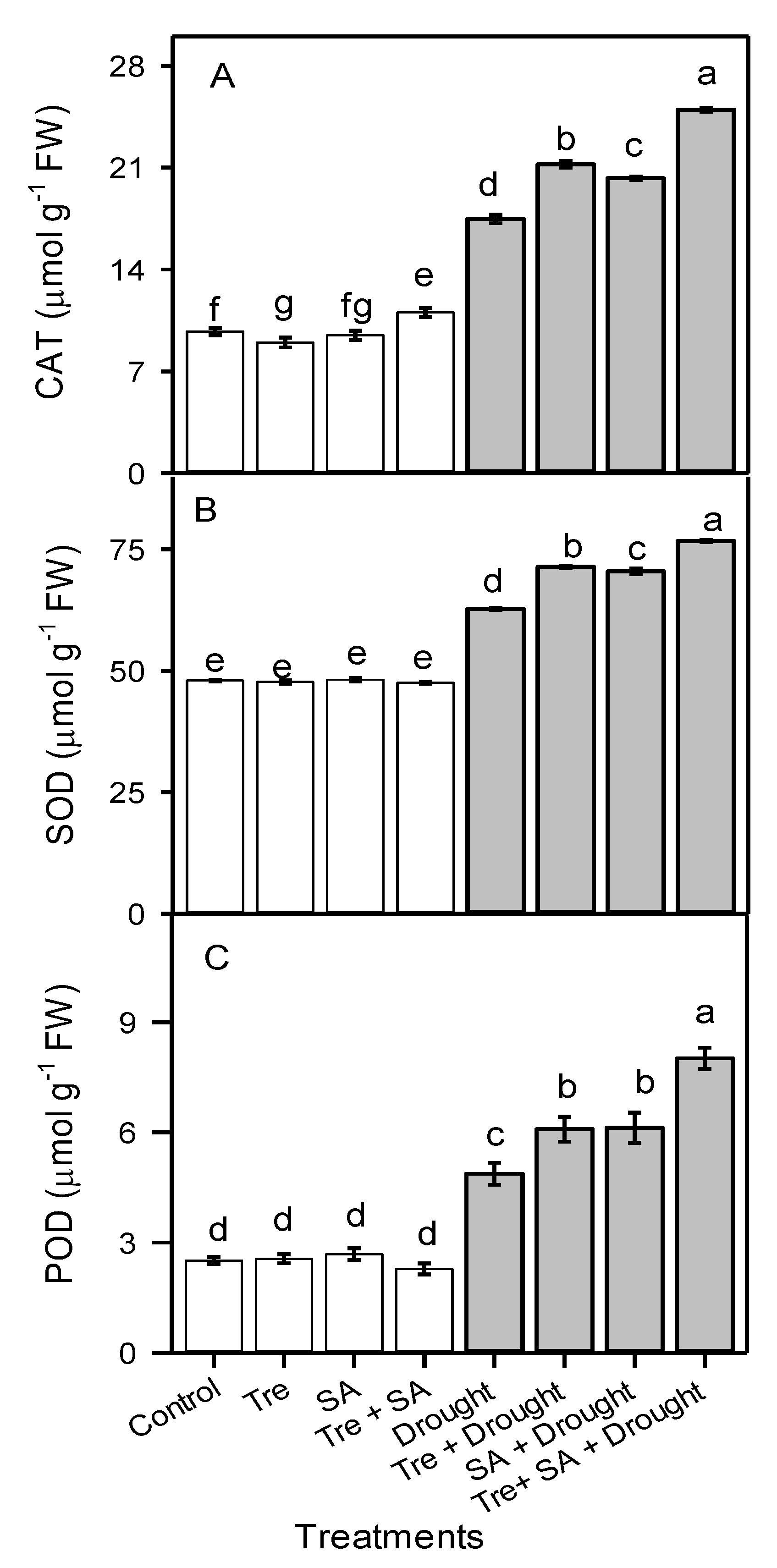
3.6. Activity of Antioxidant Enzymes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Anderson, R.; E Bayer, P.; Edwards, D. Climate change and the need for agricultural adaptation. Curr. Opin. Plant Biol. 2020, 56, 197–202. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Hancock, J.T. Hydrogen sulfide in horticulture: Emerging roles in the era of climate change. Plant Physiol. Biochem. 2020, 155, 667–675. [Google Scholar] [CrossRef]
- Pokhrel, Y.; Felfelani, F.; Satoh, Y.; Boulange, J.; Burek, P.; Gädeke, A.; Gerten, D.; Gosling, S.N.; Grillakis, M.; Gudmundsson, L.; et al. Global terrestrial water storage and drought severity under climate change. Nat. Clim. Chang. 2021, 11, 226–233. [Google Scholar] [CrossRef]
- Nam, S.; Kang, S.; Kim, J. Maintaining a constant soil moisture level can enhance the growth and phenolic content of sweet basil better than fluctuating irrigation. Agric. Water Manag. 2020, 238, 106203. [Google Scholar] [CrossRef]
- Taha, R.; Alharby, H.; Bamagoos, A.; Medani, R.; Rady, M. Elevating tolerance of drought stress in Ocimum basilicum using pollen grains extract; a natural biostimulant by regulation of plant performance and antioxidant defense system. S.Afr. J. Bot. 2020, 128, 42–53. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Younis, A.; Riaz, A.; Mansoor, F.; Hameed, M.; Akram, N.A.; Abideen, Z. Morpho-anatomical adaptations of two Tagetes erecta L. cultivars with contrasting response to drought stress. Pak. J. Bot. 2019, 52, 801–810. [Google Scholar] [CrossRef]
- Babaei, K.; Moghaddam, M.; Farhadi, N.; Pirbalouti, A.G. Morphological, physiological and phytochemical responses of Mexican marigold (Tagetes minuta L.) to drought stress. Sci. Hortic. 2021, 284, 110116. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Al Mahmud, J.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef] [PubMed]
- Zulfiqar, F.; Ashraf, M. Bioregulators: Unlocking their potential role in regulation of the plant oxidative defense system. Plant Mol. Biol. 2021, 105, 11–41. [Google Scholar] [CrossRef] [PubMed]
- Shemi, R.; Wang, R.; Gheith, E.-S.M.S.; Hussain, H.A.; Hussain, S.; Irfan, M.; Cholidah, L.; Zhang, K.; Zhang, S.; Wang, L. Effects of salicylic acid, zinc and glycine betaine on morpho-physiological growth and yield of maize under drought stress. Sci. Rep. 2021, 11, 1–14. [Google Scholar] [CrossRef]
- Seleiman, M.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.; Battaglia, M. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef] [PubMed]
- Ozturk, M.; Turkyilmaz Unal, B.; García-Caparrós, P.; Khursheed, A.; Gul, A.; Hasanuzzaman, M. Osmoregulation and its actions during the drought stress in plants. Physiol. Plant. 2020. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Akram, N.A.; Ashraf, M. Osmoprotection in plants under abiotic stresses: New insights into a classical phenom-enon. Planta 2020, 251, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosar, F.; Akram, N.A.; Ashraf, M.; Ahmad, A.; Alyemeni, M.N.; Ahmad, P. Impact of exogenously applied trehalose on leaf biochemistry, achene yield and oil composition of sunflower under drought stress. Physiol. Plant. 2020. [Google Scholar] [CrossRef]
- Hameed, A.; Akram, N.; Saleem, M.; Ashraf, M.; Ahmed, S.; Ali, S.; Alsahli, A.A.; Alyemeni, M. Seed treatment with α-tocopherol regulates growth and key physio-biochemical attributes in carrot (Daucus carota l.) plants under water limited regimes. Agron. 2021, 11, 469. [Google Scholar] [CrossRef]
- Fichtner, F.; Lunn, J.E. The role of Trehalose 6-Phosphate (Tre6P) in plant metabolism and development. Annu. Rev. Plant Biol. 2021, 72, 72. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Fu, L.; Zhang, D.; He, X.; Chen, Q.; Peng, M.; Zhang, J. Interspecies and intraspecies analysis of trehalose contents and the biosynthesis pathway gene family reveals crucial roles of trehalose in osmotic-stress tolerance in Cassava. Int. J. Mol. Sci. 2016, 17, 1077. [Google Scholar] [CrossRef]
- Acosta-Pérez, P.; Camacho-Zamora, B.D.; Espinoza-Sánchez, E.A.; Gutiérrez-Soto, G.; Zavala-García, F.; Abraham-Juárez, M.J.; Sinagawa-García, S.R. Characterization of Trehalose-6-phosphate Synthase and Trehalose-6-phosphate Phosphatase genes and analysis of its differential expression in maize (Zea mays) seedlings under drought stress. Plants 2020, 9, 315. [Google Scholar] [CrossRef] [Green Version]
- Lin, Q.; Wang, S.; Dao, Y.; Wang, J.; Wang, K.; Wang, S. Arabidopsis thaliana trehalose-6-phosphate phosphatase gene TPPI enhances drought tolerance by regulating stomatal apertures. J. Exp. Bot. 2020, 71, 4285–4297. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Li, F.; Gao, M.; Huang, Y.; Song, Z. Mechanisms of trehalose-mediated mitigation of Cd toxicity in rice seedlings. J. Clean. Prod. 2020, 267, 121982. [Google Scholar] [CrossRef]
- Lin, Y.; Zhang, J.; Gao, W.; Chen, Y.; Li, H.; Lawlor, D.W.; Paul, M.J.; Pan, W. Exogenous trehalose improves growth under limiting nitrogen through upregulation of nitrogen metabolism. BMC Plant Biol. 2017, 17, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La, V.H.; Lee, B.R.; Islam, M.T.; Park, S.H.; Jung, H.I.; Bae, D.W.; Kim, T.H. Characterization of salicylic acid-mediated mod-ulation of the drought stress responses: Reactive oxygen species, proline, and redox state in Brassica napus. Environ. Exp. Bot. 2019, 157, 1–10. [Google Scholar] [CrossRef]
- Arif, Y.; Sami, F.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salicylic acid in relation to other phytohormones in plant: A study towards physiology and signal transduction under challenging environment. Environ. Exp. Bot. 2020, 175, 104040. [Google Scholar] [CrossRef]
- Park, S.-H.; Lee, B.-R.; La, V.H.; Mamun, A.; Bae, D.-W.; Kim, T.-H. Characterization of salicylic acid- and abscisic acid-mediated photosynthesis, Ca2+ and H2O2 accumulation in two distinct phases of drought stress intensity in Brassica napus. Environ. Exp. Bot. 2021, 186, 104434. [Google Scholar] [CrossRef]
- Abbaszadeh, B.; Layeghhaghighi, M.; Azimi, R.; Hadi, N. Improving water use efficiency through drought stress and using salicylic acid for proper production of Rosmarinus officinalis L. Ind. Crop. Prod. 2020, 144, 111893. [Google Scholar] [CrossRef]
- Chavoushi, M.; Najafi, F.; Salimi, A.; Angaji, S. Improvement in drought stress tolerance of safflower during vegetative growth by exogenous application of salicylic acid and sodium nitroprusside. Ind. Crop. Prod. 2019, 134, 168–176. [Google Scholar] [CrossRef]
- Damalas, C.A. Improving drought tolerance in sweet basil (Ocimum basilicum) with salicylic acid. Sci. Hortic. 2019, 246, 360–365. [Google Scholar] [CrossRef]
- Ioannidis, D.; Bonner, L.; Johnson, C.B. UV-B is required for normal development of oil glands in Ocimum basilicum L. (Sweet Basil). Ann. Bot. 2002, 90, 453–460. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.; Vasiliev, M.; Alameh, K. LED Illumination spectrum manipulation for increasing the yield of sweet basil (Ocimum basilicum L.). Plants 2021, 10, 344. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Carillo, P.; Cristofano, F.; Cardarelli, M.; Colla, G. Effects of vegetal- versus animal-derived protein hydrolysate on sweet basil morpho-physiological and metabolic traits. Sci. Hortic. 2021, 284, 110123. [Google Scholar] [CrossRef]
- Rihan, H.Z.; Aldarkazali, M.; Mohamed, S.J.; McMulkin, N.B.; Jbara, M.H.; Fuller, M.P. A novel new light recipe significantly increases the growth and yield of sweet basil (Ocimum basilicum) grown in plant factory system. Agronomy. 2020, 10, 934. [Google Scholar] [CrossRef]
- Pirbalouti, A.G.; Malekpoor, F.; Salimi, A.; Golparvar, A. Exogenous application of chitosan on biochemical and physiological characteristics, phenolic content and antioxidant activity of two species of basil (Ocimum ciliatum and Ocimum basilicum) under reduced irrigation. Sci. Hortic. 2017, 217, 114–122. [Google Scholar] [CrossRef]
- Lazarević, B.; Šatović, Z.; Nimac, A.; Vidak, M.; Gunjača, J.; Politeo, O.; Carović-Stanko, K. Application of phenotyping methods in detection of drought and salinity stress in Basil (Ocimum basilicum L.). Front. Plant. Sci. 2021, 12, 174. [Google Scholar] [CrossRef]
- Kulak, M.; Jorrín-Novo, J.V.; Romero-Rodriguez, M.C.; Yildirim, E.D.; Gul, F.; Karaman, S. Seed priming with salicylic acid on plant growth and essential oil composition in basil (Ocimum basilicum L.) plants grown under water stress conditions. Ind. Crop. Prod. 2021, 161, 113235. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzyme in isolated chloroplast: Polyphenol oxidase in Beta vulgaris. Plant. Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Julkenen-Titto, R. Phenolic constituents in the leaves of northern willows: Methods for the analysis of certain phenolics. J. Agric. Food Chem. 1985, 33, 213–217. [Google Scholar] [CrossRef]
- Dionisio-Sese, M.L.; Tobita, S. Antioxidant responses of rice seedlings to salinity stress. Plant Sci. 1998, 135, 1–9. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Grieve, C.M.; Grattan, S.R. Rapid assay for determination of water soluble quaternary ammonium compounds. Plant Soil 1983, 70, 303–307. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Pro-tective role of exogenous polyamines. Plant. Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in Isolated Chloroplasts: I. Kinetics and Stoichiometry of Fatty Acid Peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A. Assay of catalases and peroxidases. Methods Enzym. 1955, 2, 764–775. [Google Scholar] [CrossRef]
- Van Rossum, M.W.P.C.; Alberda, M.; Van Der Plas, L.H.W. Role of oxidative damage in tulip bulb scale micropropagation. Plant Sci. 1997, 130, 207–216. [Google Scholar] [CrossRef]
- Kosar, F.; Akram, N.A.; Ashraf, M.; Sadiq, M.; Al-Qurainy, F. Trehalose-induced improvement in growth, photosynthetic characteristics and levels of some key osmoprotectants in sunflower (Helianthus annuus L.) under drought stress. Pak. J. Bot. 2018, 50, 955–961. [Google Scholar]
- Najafabadi, M.Y.; Ehsanzadeh, P. Photosynthetic and antioxidative upregulation in drought-stressed sesame (Sesamum indicum L.) subjected to foliar-applied salicylic acid. Photosynthetica 2017, 55, 611–622. [Google Scholar] [CrossRef]
- El-Mageed, T.A.; Semida, W.M.; Mohamed, G.F.; Rady, M.M. Combined effect of foliar-applied salicylic acid and deficit irri-gation on physiological–anatomical responses, and yield of squash plants under saline soil. S. Afr. J. Bot. 2016, 106, 8–16. [Google Scholar] [CrossRef]
- Jesus, C.; Meijón, M.; Monteiro, P.; Correia, B.; Amaral, J.; Escandón, M.; Cañal, M.J.; Pinto, G. Salicylic acid application modulates physiological and hormonal changes in Eucalyptus globulus under water deficit. Environ. Exp. Bot. 2015, 118, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Bijanzadeh, E.; Naderi, R.; Egan, T.P. Exogenous application of humic acid and salicylic acid to alleviate seedling drought stress in two corn (Zea mays L.) hybrids. J. Plant Nutr. 2019, 42, 1483–1495. [Google Scholar] [CrossRef]
- La, V.H.; Lee, B.-R.; Zhang, Q.; Park, S.-H.; Islam, T.; Kim, T.-H. Salicylic acid improves drought-stress tolerance by regulating the redox status and proline metabolism in Brassica rapa. Hortic. Environ. Biotechnol. 2018, 60, 31–40. [Google Scholar] [CrossRef]
- Maghsoudi, K.; Emam, Y.; Ashraf, M.; Arvin, M.J. Alleviation of field water stress in wheat cultivars by using silicon and salicylic acid applied separately or in combination. Crop. Pasture Sci. 2019, 70, 36. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of Phenylpropanoid Pathway and the Role of Polyphenols in Plants under Abiotic Stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [Green Version]
- Gharibi, S.; Sayed Tabatabaei, B.E.; Saeidi, G.; Talebi, M.; Matkowski, A. The effect of drought stress on polyphenolic com-pounds and expression of flavonoid biosynthesis related genes in Achillea pachycephala Rech.f. Phytochemistry 2019, 162, 90–98. [Google Scholar] [CrossRef]
- Per, T.S.; Khan, N.A.; Reddy, P.S.; Masood, A.; Hasanuzzaman, M.; Khan, M.I.R.; Anjum, N.A. Approaches in modulating proline metabolism in plants for salt and drought stress tolerance: Phytohormones, mineral nutrients and transgenics. Plant Physiol. Biochem. 2017, 115, 126–140. [Google Scholar] [CrossRef]
- Annunziata, M.G.; Ciarmiello, L.F.; Woodrow, P.; Dell’Aversana, E.; Carillo, P. Spatial and temporal profile of glycine betaine accumulation in plants under abiotic stresses. Front. Plant Sci. 2019, 10, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, H.A.; Abdellatif, Y.M. Effect of maltose and trehalose on growth, yield and some biochemical components of wheat plant under water stress. Ann. Agric. Sci. 2016, 61, 267–274. [Google Scholar] [CrossRef] [Green Version]
- Nazar, R.; Umar, S.; Khan, N.; Sareer, O. Salicylic acid supplementation improves photosynthesis and growth in mustard through changes in proline accumulation and ethylene formation under drought stress. S. Afr. J. Bot. 2015, 98, 84–94. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Chl-a (mg·g−1 FW) | Chl-b (mg·g−1 FW) | Total Chl (mg g−1 FW) |
|---|---|---|---|
| Control (CK) | 1.75 ± 0.006 d | 0.56 ± 0.003 d | 2.31 ± 0.004 d |
| CK + Tre | 1.87 ± 0.003 b | 0.63 ± 0.006 b | 2.50 ± 0.003 b |
| CK + SA | 1.82 ± 0.003 c | 0.60 ± 0.006 c | 2.43 ± 0.007 c |
| CK + Tre + SA | 1.99 ± 0.005 a | 0.71 ± 0.009 a | 2.70 ± 0.007 a |
| Drought stress | 0.98 ± 0.007 h | 0.34 ± 0.005 h | 1.33 ± 0.007 h |
| Drought stress + Tre | 1.43 ± 0.004 f | 0.45 ± 0.003 f | 1.87 ± 0.002 f |
| Drought stress + SA | 1.28 ± 0.005 g | 0.40 ± 0.006 g | 1.67 ± 0.004 g |
| Drought stress + Tre + SA | 1.60 ± 0.007 e | 0.50 ± 0.009 e | 2.10 ± 0.012 e |
| Treatments | CO2 Assimilation Rate (A) (µmol CO2·m−2·s−1) | Stomatal Conductance (gs) (mmol H2O·m−2·s−1) | Transpiration (E) (mmol H2O·m−2·s−1) | Intercellular CO2 conc. (Ci) (cm3·m−3) |
|---|---|---|---|---|
| Control (CK) | 6.49 ± 0.13 d | 53.00 ± 1.44 d | 3.21 ± 0.017 c | 222.67 ± 1.38 c |
| CK + Tre | 7.71 ± 0.08 b | 74.67 ± 1.38 b | 3.89 ± 0.035 a | 241.00 ± 2.35 b |
| CK + SA | 7.26 ± 0.05 c | 68.17 ± 3.79 c | 3.72 ± 0.024 b | 235.50 ± 2.09 b |
| CK + Tre + SA | 8.74 ± 0.07 a | 88.67 ± 2.79 a | 3.98 ± 0.016 a | 273.50 ± 2.96 a |
| Drought stress | 3.29 ± 0.08 h | 10.67 ± 0.61 g | 0.61 ± 0.061 f | 141.17 ± 2.15 g |
| Drought stress + Tre | 4.85 ± 0.03 f | 25.50 ± 0.81 f | 1.13 ± 0.017 e | 178.50 ± 3.42 e |
| Drought stress + SA | 4.14 ± 0.02 g | 23.83 ± 0.91 f | 1.04 ± 0.039 e | 169.17 ± 0.95 f |
| Drought stress + Tre + SA | 5.49 ± 0.06 e | 44.83 ± 1.19 e | 1.57 ± 0.060 d | 194.50 ± 2.36 d |
| Treatments | PSII Efficiency (Fv/Fm) | PSII Quantum Yield (FPSII) | Photochemical Quenching (qp) | Non-Photochemical Quenching (NPQ) |
|---|---|---|---|---|
| Control (CK) | 0.80 ± 0.008 b | 0.61 ± 0.02 b | 0.87 ± 0.01 a | 0.63 ± 0.01 d |
| CK + Tre | 0.86 ± 0.003 a | 0.68 ± 0.01 b | 0.91 ± 0.009 a | 0.44 ± 0.01 e |
| CK + SA | 0.83 ± 0.007 a | 0.63 ± 0.007 b | 0.90 ± 0.013 a | 0.39 ± 0.01 e |
| CK + Tre + SA | 0.91 ± 0.011 a | 0.73 ± 0.02 a | 0.94 ± 0.016 a | 0.35 ± 0.009 f |
| Drought stress | 0.55 ± 0.004 d | 0.46 ± 0.007 d | 0.60 ± 0.006 d | 0.95 ± 0.004 a |
| Drought stress + Tre | 0.67 ± 0.020 c | 0.55 ± 0.007 c | 0.71 ± 0.010 c | 0.73 ± 0.012 c |
| Drought stress + SA | 0.62 ± 0.007 c | 0.51 ± 0.008 c | 0.67 ± 0.010 c | 0.83 ± 0.025 b |
| Drought stress + Tre + SA | 0.90 ± 0.012 a | 0.76 ± 0.013 a | 0.81 ± 0.005 b | 0.68 ± 0.009 d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zulfiqar, F.; Chen, J.; Finnegan, P.M.; Younis, A.; Nafees, M.; Zorrig, W.; Hamed, K.B. Application of Trehalose and Salicylic Acid Mitigates Drought Stress in Sweet Basil and Improves Plant Growth. Plants 2021, 10, 1078. https://doi.org/10.3390/plants10061078
Zulfiqar F, Chen J, Finnegan PM, Younis A, Nafees M, Zorrig W, Hamed KB. Application of Trehalose and Salicylic Acid Mitigates Drought Stress in Sweet Basil and Improves Plant Growth. Plants. 2021; 10(6):1078. https://doi.org/10.3390/plants10061078
Chicago/Turabian StyleZulfiqar, Faisal, Jianjun Chen, Patrick M. Finnegan, Adnan Younis, Muhammad Nafees, Walid Zorrig, and Karim Ben Hamed. 2021. "Application of Trehalose and Salicylic Acid Mitigates Drought Stress in Sweet Basil and Improves Plant Growth" Plants 10, no. 6: 1078. https://doi.org/10.3390/plants10061078
APA StyleZulfiqar, F., Chen, J., Finnegan, P. M., Younis, A., Nafees, M., Zorrig, W., & Hamed, K. B. (2021). Application of Trehalose and Salicylic Acid Mitigates Drought Stress in Sweet Basil and Improves Plant Growth. Plants, 10(6), 1078. https://doi.org/10.3390/plants10061078