
Safflower Extract Inhibits ADP-Induced Human Platelet Aggregation

 ,
,  , ,
, ,  and
and 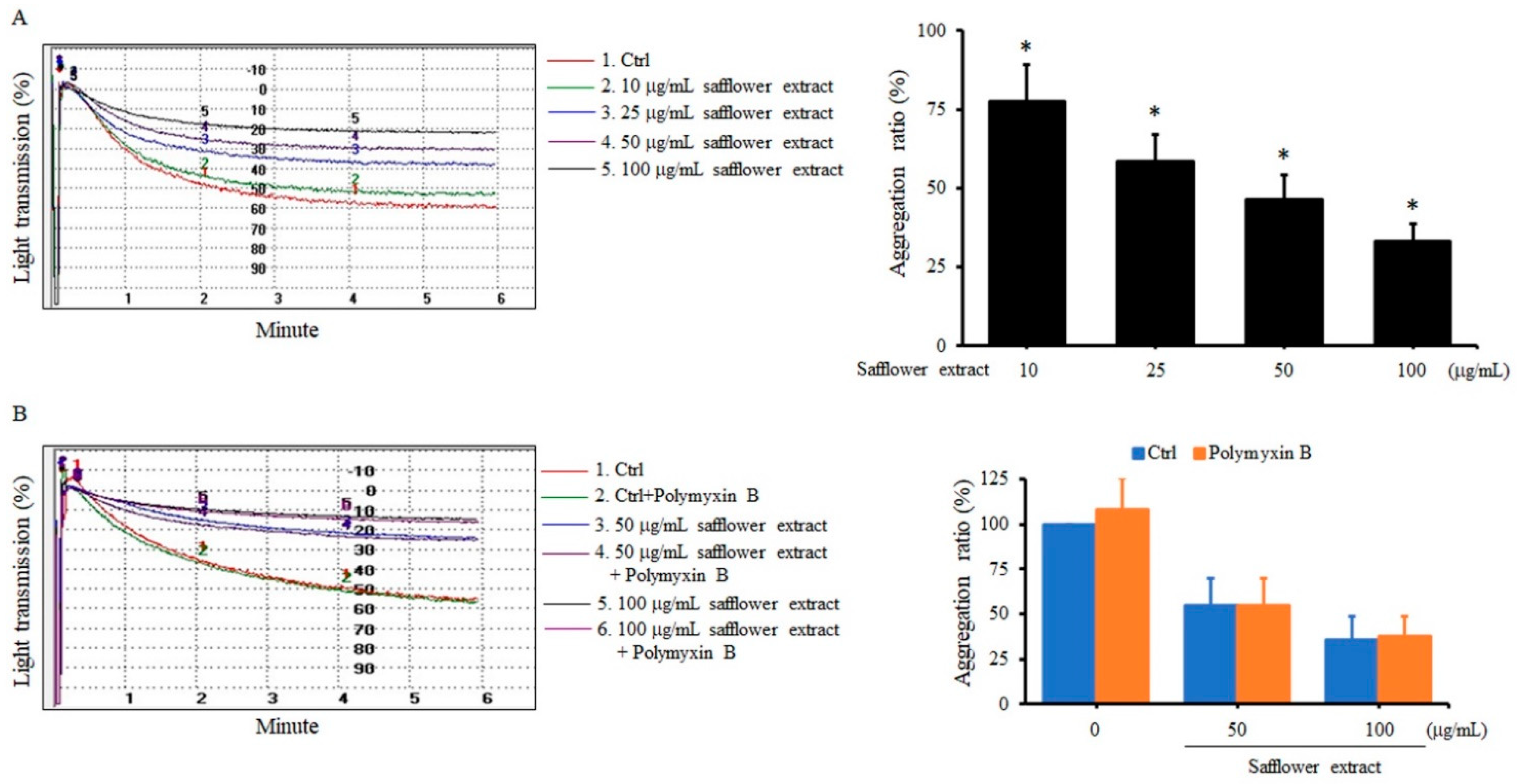{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Safflower Extract Inhibited ADP-Induced Human Platelet Aggregation
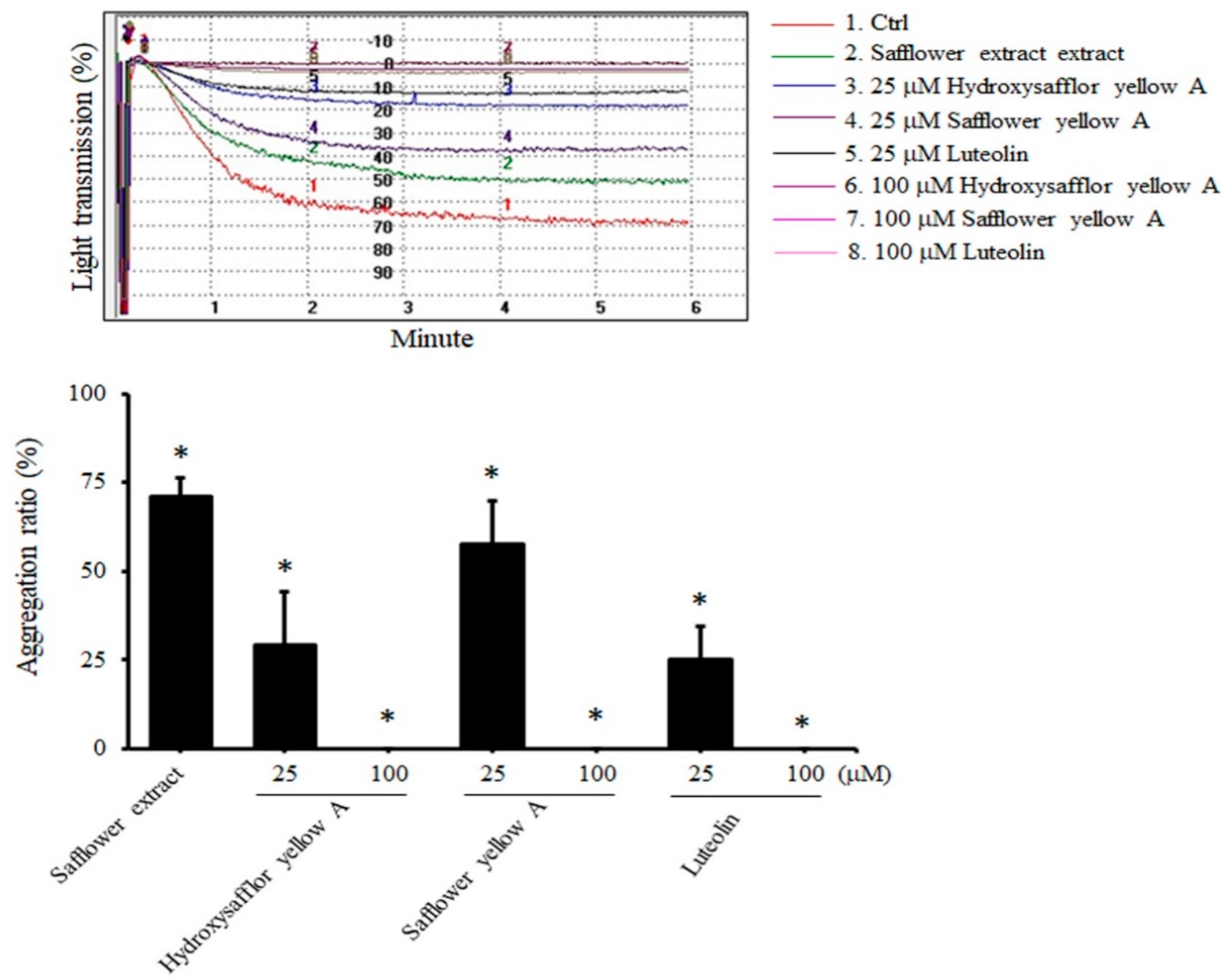
2.2. All Compounds Found in Safflower Inhibited ADP-Induced Human Platelet Aggregation
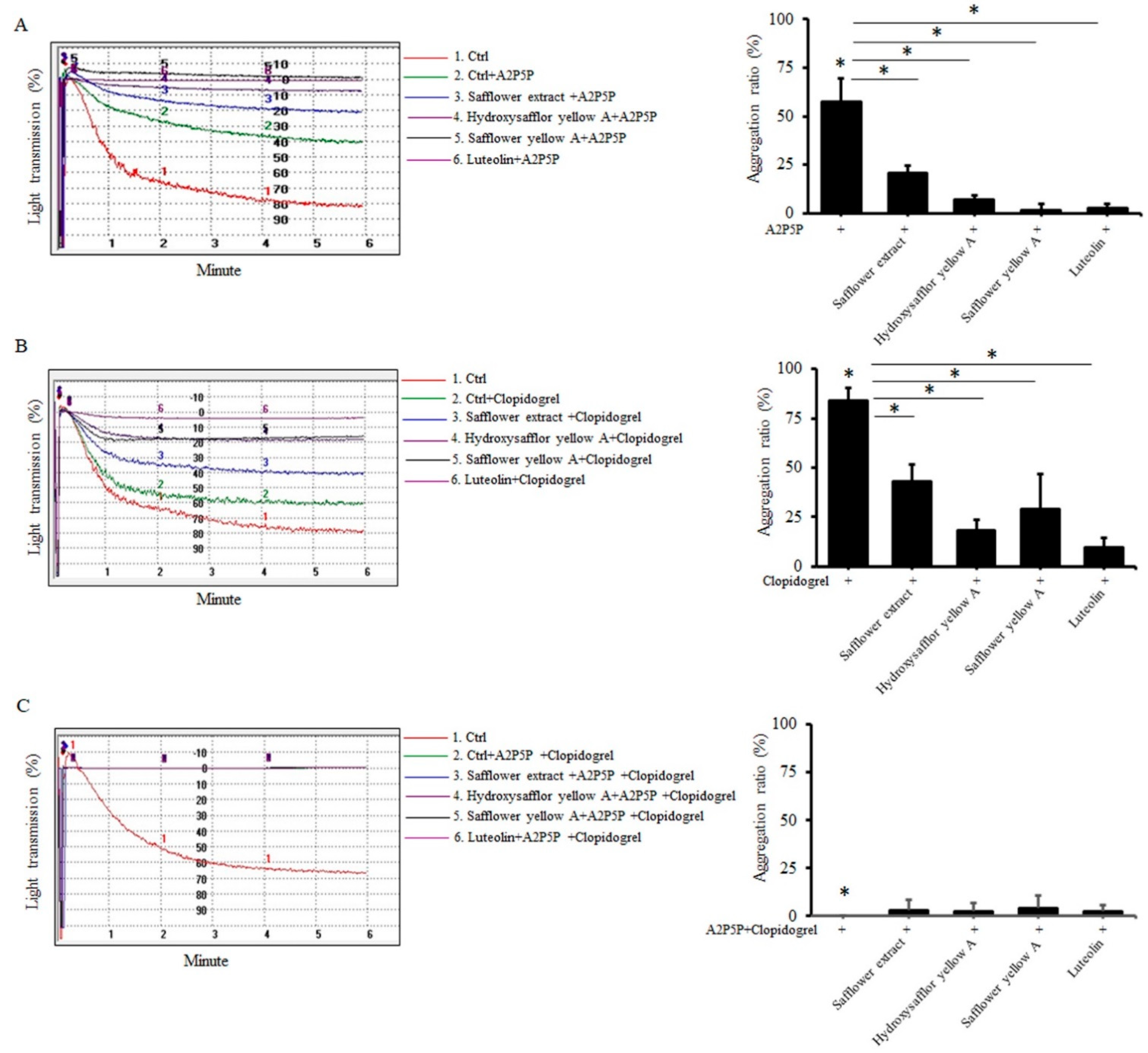
2.3. Safflower Extract and Its Compounds Inhibit Platelet Aggregation through P2Y1 and P2Y12 Receptors
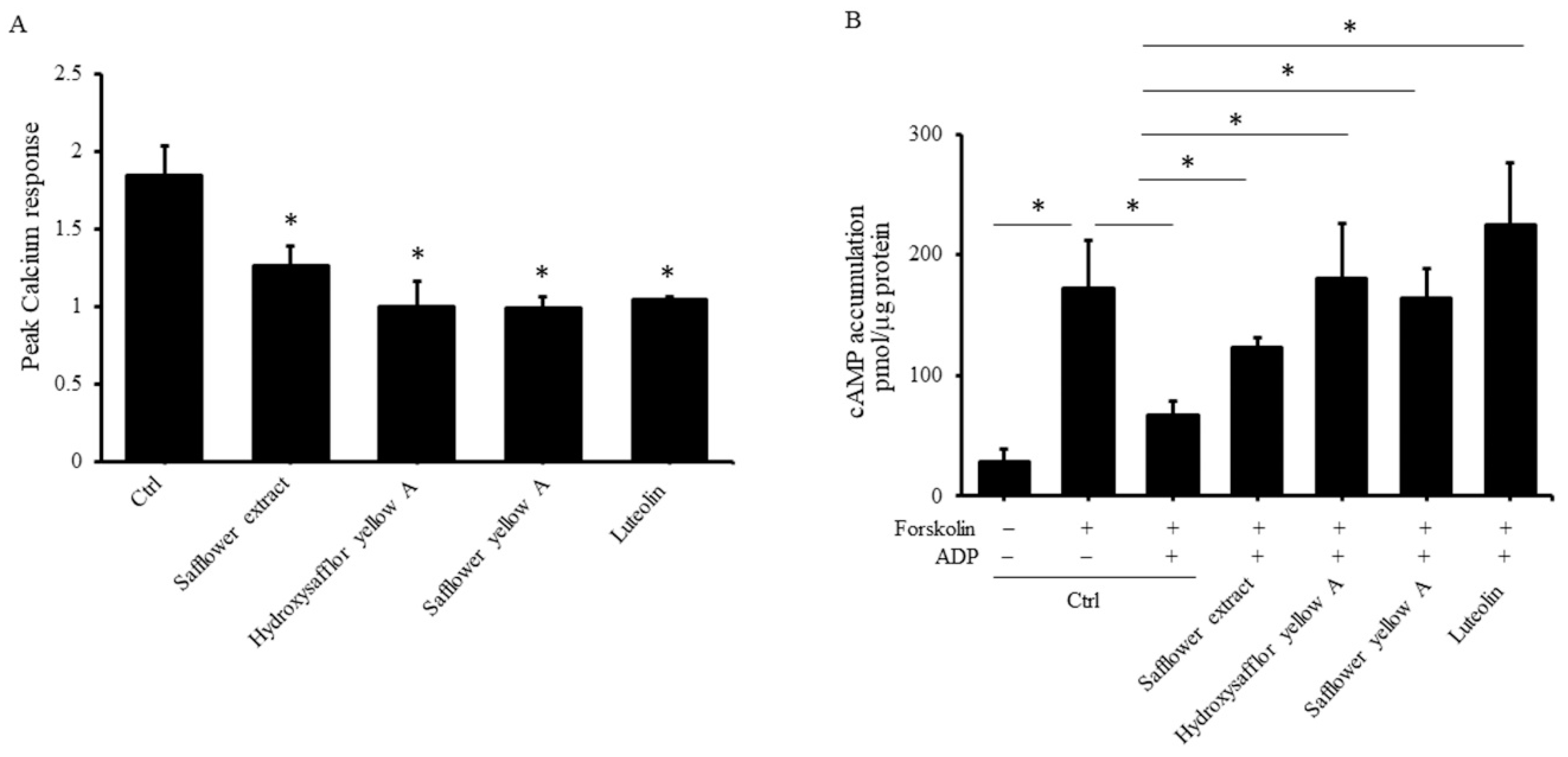
2.4. Safflower Extract and Its Compounds Inhibit the Activation of Calcium Ion Influx and the Production of cAMP Regulated by ADP
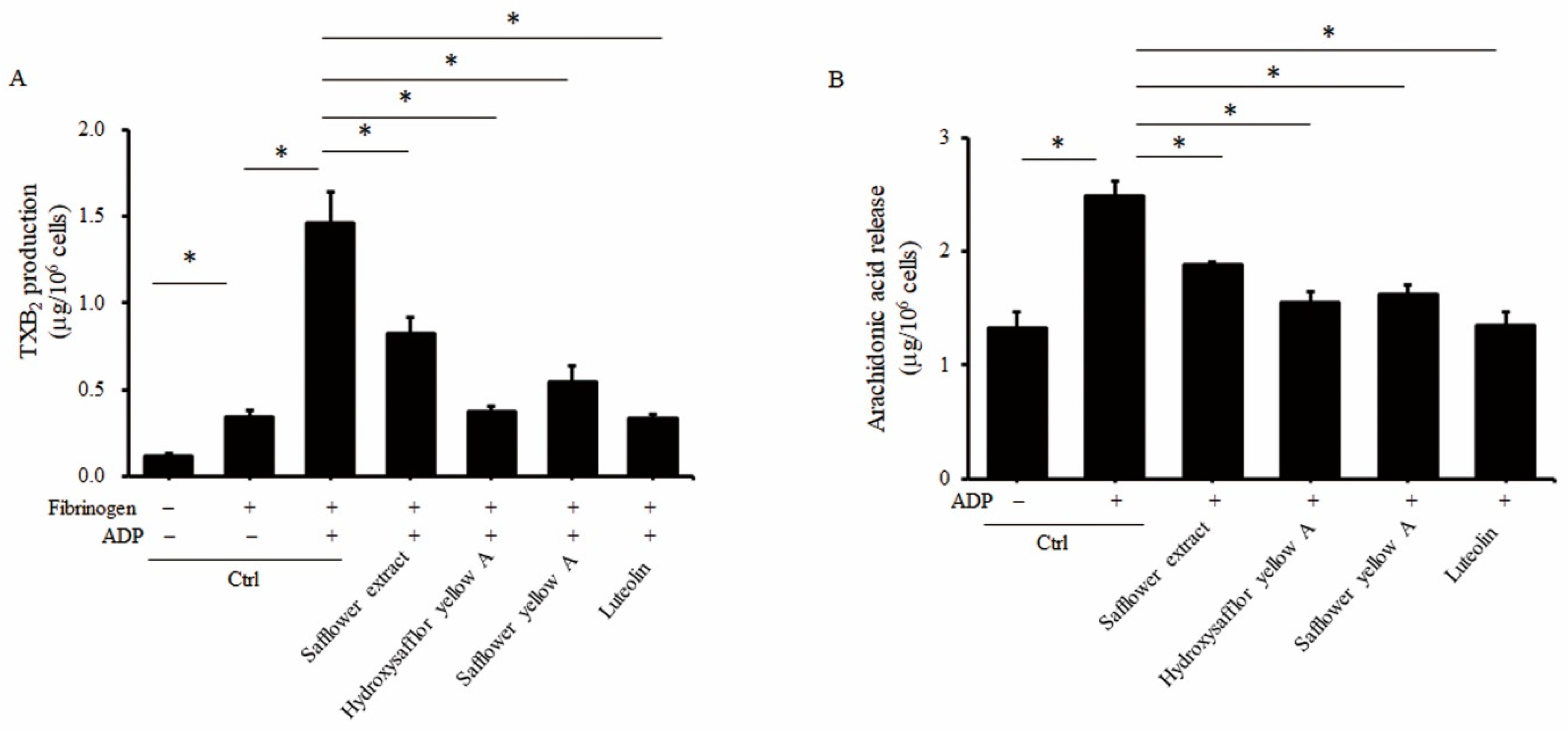
2.5. Safflower Extract and Its Compounds Inhibit the Production of Thromboxane A2 (TXA2) and Arachidonic Acid (AA)
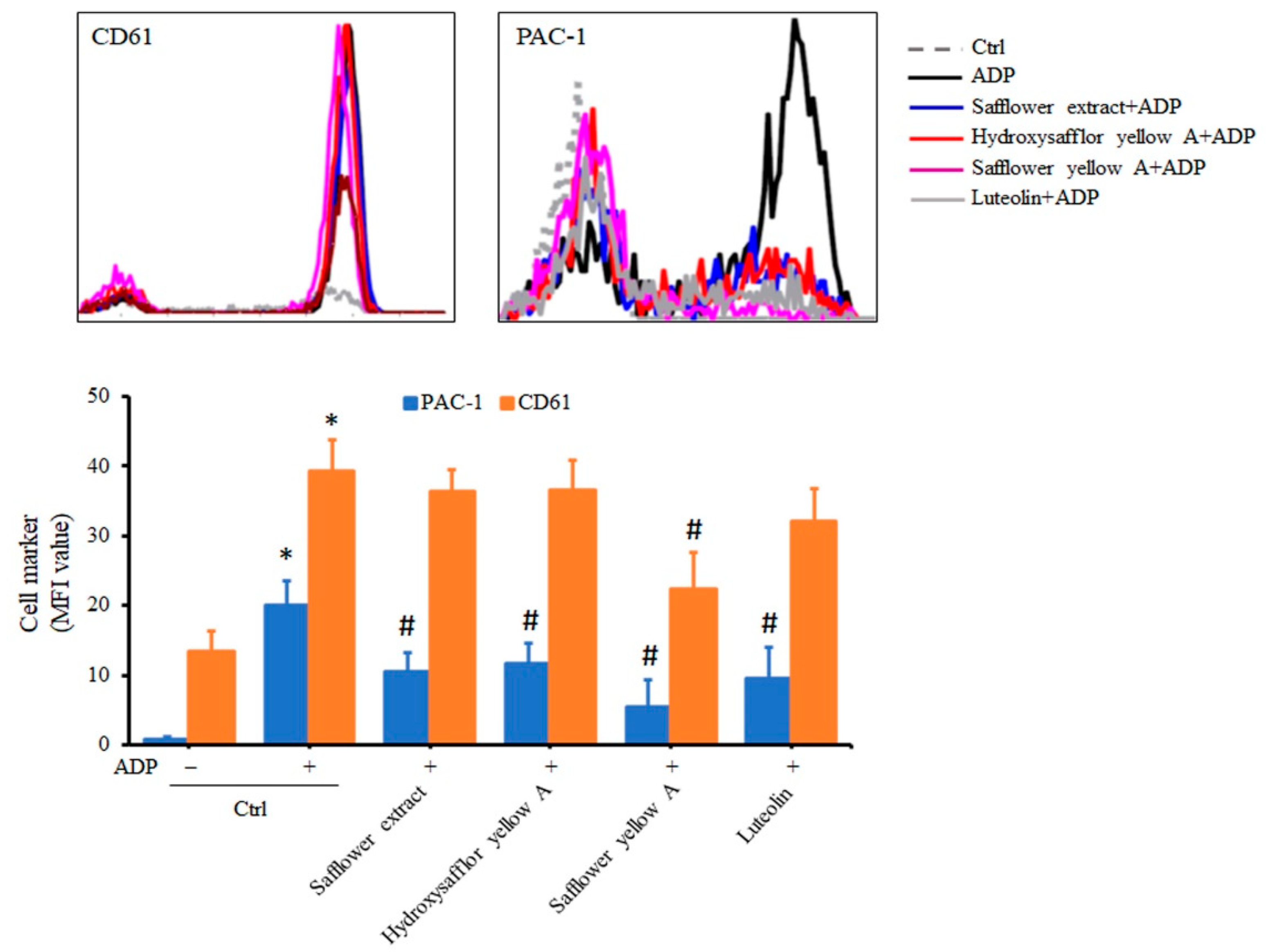
2.6. Safflower Extract and Its Compounds Inhibit the Formation of PAC-1 in Platelets
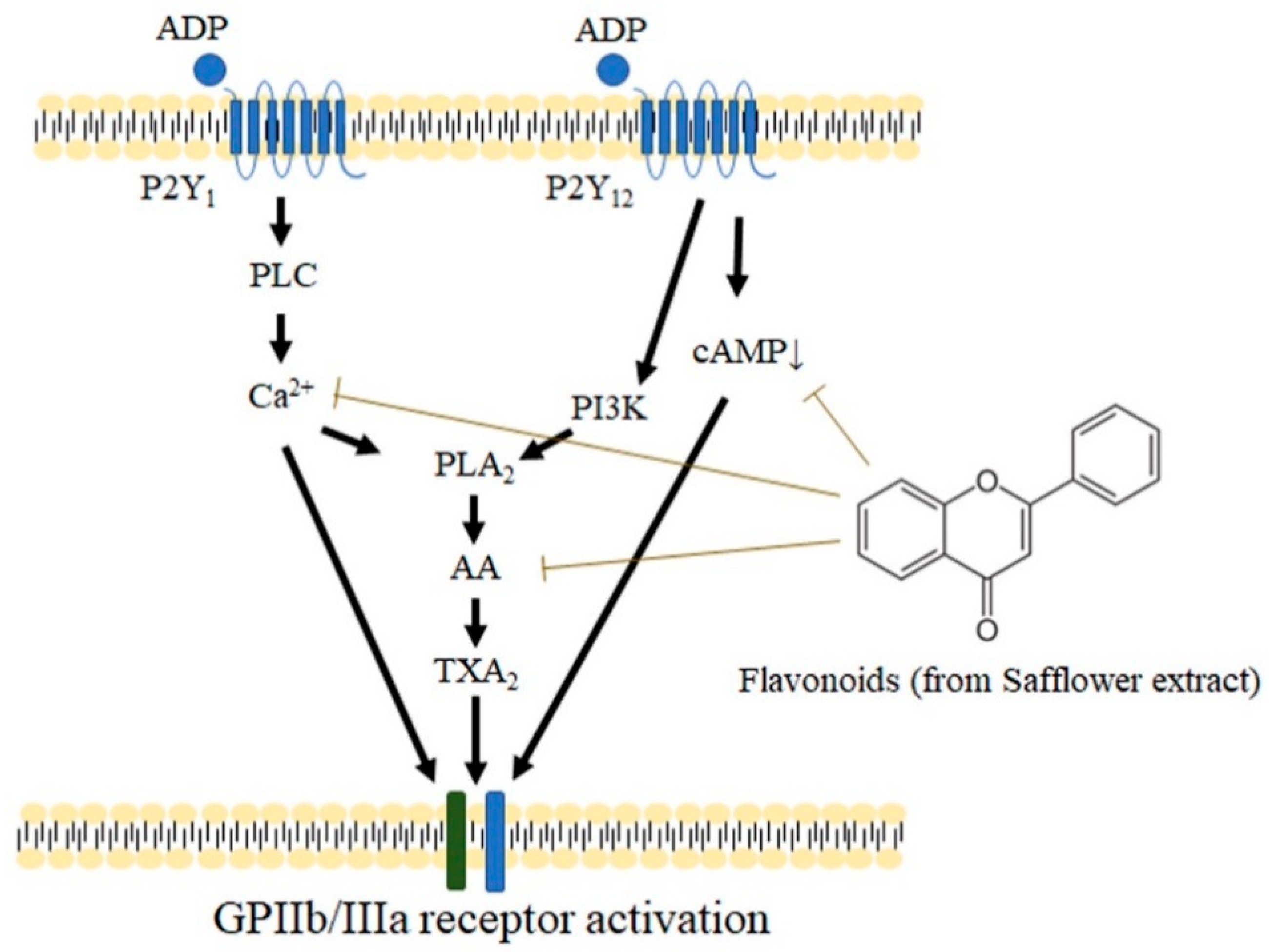
3. Discussion
4. Materials and Methods
4.1. Preparation of the Aqueous Extract of Safflower
4.2. Preparation of Platelet-Rich Plasma (PRP), Platelet-Poor Plasma (PPP), and Purified Platelets
4.3. Measurement of Platelet Aggregation
4.4. Detection of Calcium Ions and cAMP in Platelets
4.5. Determination of Thromboxane B2 (TXB2) and Arachidonic acid (AA) in Platelets
4.6. Measurement of Platelet Surface Activation Markers
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| TXA2 | thromboxane A2 |
| cAMP | cyclic AMP |
| PRP | platelet-rich plasma |
| PPP | platelet-poor plasma |
| TXB2 | thromboxane B2 |
| AA | arachidonic acid |
References
- Hussain, M.I.; Al-Dakheel, A.J. Effect of salinity stress on phenotypic plasticity, yield stability, and signature of stable isotopes of carbon and nitrogen in safflower. Environ. Sci. Pollut. Res. Int. 2018, 25, 23685–23694. [Google Scholar] [CrossRef]
- Delshad, E.; Yousefi, M.; Sasannezhad, P.; Rakhshandeh, H.; Ayati, Z. Medical uses of Carthamus tinctorius L. (Safflower): A comprehensive review from Traditional Medicine to Modern Medicine. Electron. Physician 2018, 10, 6672–6681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mani, V.; Lee, S.K.; Yeo, Y.; Hahn, B.S. A Metabolic Perspective and Opportunities in Pharmacologically Important Safflower. Metabolites 2020, 10, 253. [Google Scholar] [CrossRef]
- Asgarpanah, J.; Kazemivash, N. Phytochemistry, pharmacology and medicinal properties of Carthamus tinctorius L. Chin. J. Integr. Med. 2013, 19, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.Y.; Ma, J.Q.; Duan, Y.Y.; Sun, Y.; Yu, S.; Li, B.; Zhang, G.M. Carthamin yellow protects the heart against Ischemia/Reperfusion Injury with reduced reactive oxygen Species Release and Inflammatory Response. J. Cardiovasc. Pharmacol. 2019, 74, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Zheng, M.; Pan, R.; Zang, B.; Gao, J.; Ma, H.; Jin, M. Hydroxysafflor yellow A (HSYA) targets the platelet-activating factor (PAF) receptor and inhibits human bronchial smooth muscle activation induced by PAF. Food Funct. 2019, 10, 4661–4673. [Google Scholar] [CrossRef]
- Du, S.; Deng, Y.; Yuan, H.; Sun, Y. Safflower yellow B protects brain against cerebral ischemia reperfusion injury through AMPK/NF-kB pathway. Evid. Based Complement Alternat. Med. 2019, 2019, 7219740. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Li, J.; Song, H.; Xiong, Y.; Liu, D.; Bai, X. Hydroxysafflor yellow A suppresses angiogenesis of hepatocellular carcinoma through inhibition of p38 MAPK phosphorylation. Biomed. Pharmacother. 2019, 109, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Feng, C.; Wang, J.; Chen, Z.; Wei, P.; Fan, A.; Wang, X.; Yu, X.; Ge, D.; Xie, H.; et al. Hydroxyl safflower yellow A regulates the tumor immune microenvironment to produce an anticancer effect in a mouse model of hepatocellular carcinoma. Oncol. Lett. 2019, 17, 3503–3510. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.Y.; Lee, Y.J.; Kim, J.D.; Kang, S.N.; Lee, S.K.; Jang, J.Y.; Lee, H.K.; Lim, J.H.; Lee, O.H. Phenolic composition, antioxidant activity and anti-adipogenic effect of hot water extract from safflower (Carthamus tinctorius L.) seed. Nutrients 2013, 5, 4894–4907. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.H.; Jang, J.H.; Yoon, J.Y.; Han, C.D.; Choi, Y.; Choi, S.W. Effects of a safflower tea supplement on antioxidative status and bone markers in postmenopausal women. Nutr. Res. Pract. 2011, 5, 20–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, H.O.; Park, Y.S.; Lee, J.H.; Seo, J.B.; Koo, K.I.; Jeong, S.C.; Jin, S.D.; Lee, Y.H.; Eom, H.S.; Yun, I. Effect of extracts from safflower seeds on osteoblast differentiation and intracellular calcium ion concentration in MC3T3-E1 cells. Nat. Prod. Res. 2007, 21, 787–797. [Google Scholar] [CrossRef]
- Alam, M.R.; Kim, S.M.; Lee, J.I.; Chon, S.K.; Choi, S.J.; Choi, I.H.; Kim, N.S. Effects of Safflower seed oil in osteoporosis induced-ovariectomized rats. Am. J. Chin. Med. 2006, 34, 601–612. [Google Scholar] [CrossRef]
- Kim, J.H.; He, M.T.; Kim, M.J.; Yang, C.Y.; Shin, Y.S.; Yokozawa, T.; Park, C.H.; Cho, E.J. Safflower (Carthamus tinctorius L.) seed attenuates memory impairment induced by scopolamine in mice via regulation of cholinergic dysfunction and oxidative stress. Food Funct. 2019, 10, 3650–3659. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, Z.; Zhai, W.; Pang, J.; Mo, Y.; Yang, G.; Qu, Z.; Hu, Y. Safflower yellow attenuates learning and memory deficits in amyloid beta-induced Alzheimer’s disease rats by inhibiting neuroglia cell activation and inflammatory signaling pathways. Metab. Brain Dis. 2019, 34, 927–939. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Guo, J.; Dong, H.; Zhao, X.; Zhou, L.; Li, X.; Liu, J.; Niu, Y. Hydroxysafflor yellow A protects against chronic carbon tetrachloride-induced liver fibrosis. Eur. J. Pharmacol. 2011, 660, 438–444. [Google Scholar] [CrossRef]
- van der Meijden, P.E.J.; Heemskerk, J.W.M. Platelet biology and functions: New concepts and clinical perspectives. Nat. Rev. Cardiol. 2019, 16, 166–179. [Google Scholar] [CrossRef]
- Ghoshal, K.; Bhattacharyya, M. Overview of platelet physiology: Its hemostatic and nonhemostatic role in disease pathogenesis. Sci. World J. 2014, 2014, 781857. [Google Scholar] [CrossRef] [Green Version]
- Golebiewska, E.M.; Poole, A.W. Platelet secretion: From haemostasis to wound healing and beyond. Blood Rev. 2015, 29, 153–162. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.R.; Carrim, N.; Neves, M.A.; McKeown, T.; Stratton, T.W.; Coelho, R.M.; Lei, X.; Chen, P.; Xu, J.; Dai, X.; et al. Platelets and platelet adhesion molecules: Novel mechanisms of thrombosis and anti-thrombotic therapies. Thromb. J. 2016, 14, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruggeri, Z.M.; Mendolicchio, G.L. Adhesion mechanisms in platelet function. Circ. Res. 2007, 100, 1673–1685. [Google Scholar] [CrossRef] [PubMed]
- Reininger, A.J.; Heijnen, H.F.; Schumann, H.; Specht, H.M.; Schramm, W.; Ruggeri, Z.M. Mechanism of platelet adhesion to von Willebrand factor and microparticle formation under high shear stress. Blood 2006, 107, 3537–3545. [Google Scholar] [CrossRef]
- Tomokiyo, K.; Kamikubo, Y.; Hanada, T.; Araki, T.; Nakatomi, Y.; Ogata, Y.; Jung, S.M.; Nakagaki, T.; Moroi, M. Von Willebrand factor accelerates platelet adhesion and thrombus formation on a collagen surface in platelet-reduced blood under flow conditions. Blood 2005, 105, 1078–1084. [Google Scholar] [CrossRef] [PubMed]
- Gerhards, C.; Uhlig, S.; Etemad, M.; Christodoulou, F.; Bieback, K.; Kluter, H.; Bugert, P. Expression of ADP receptor P2Y12, thromboxane A2 receptor and C-type lectin-like receptor 2 in cord blood-derived megakaryopoiesis. Platelets 2021, 32, 618–625. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Schmaier, A.H. Platelet aggregation testing in platelet-rich plasma: Description of procedures with the aim to develop standards in the field. Am. J. Clin. Pathol. 2005, 123, 172–183. [Google Scholar] [CrossRef]
- Woulfe, D.; Yang, J.; Brass, L. ADP and platelets: The end of the beginning. J. Clin. Investig. 2001, 107, 1503–1505. [Google Scholar] [CrossRef]
- Jarvis, G.E.; Humphries, R.G.; Robertson, M.J.; Leff, P. ADP can induce aggregation of human platelets via both P2Y(1) and P(2T) receptors. Br. J. Pharmacol. 2000, 129, 275–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandyopadhyay, S.K.; Azharuddin, M.; Dasgupta, A.K.; Ganguli, B.; SenRoy, S.; Patra, H.K.; Deb, S. Probing ADP Induced Aggregation Kinetics During Platelet-Nanoparticle Interactions: Functional Dynamics Analysis to Rationalize Safety and Benefits. Front. Bioeng. Biotechnol. 2019, 7, 163. [Google Scholar] [CrossRef] [Green Version]
- Shah, B.H.; Rasheed, H.; Rahman, I.H.; Shariff, A.H.; Khan, F.L.; Rahman, H.B.; Hanif, S.; Saeed, S.A. Molecular mechanisms involved in human platelet aggregation by synergistic interaction of platelet-activating factor and 5-hydroxytryptamine. Exp. Mol. Med. 2001, 33, 226–233. [Google Scholar] [CrossRef] [Green Version]
- Li, H.X.; Han, S.Y.; Wang, X.W.; Ma, X.; Zhang, K.; Wang, L.; Ma, Z.Z.; Tu, P.F. Effect of the carthamins yellow from Carthamus tinctorius L. on hemorheological disorders of blood stasis in rats. Food Chem. Toxicol. 2009, 47, 1797–1802. [Google Scholar] [CrossRef]
- Zhu, H.B.; Zhang, L.; Wang, Z.H.; Tian, J.W.; Fu, F.H.; Liu, K.; Li, C.L. Therapeutic effects of hydroxysafflor yellow A on focal cerebral ischemic injury in rats and its primary mechanisms. J. Asian Nat. Prod. Res. 2005, 7, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Yan, K.; Wang, X.; Pan, H.; Wang, L.; Yang, H.; Liu, M.; Zhu, H.; Gong, F. Safflower Yellow and Its Main Component HSYA Alleviate Diet-Induced Obesity in Mice: Possible Involvement of the Increased Antioxidant Enzymes in Liver and Adipose Tissue. Front. Pharmacol. 2020, 11, 482. [Google Scholar] [CrossRef] [PubMed]
- Maleki, S.J.; Crespo, J.F.; Cabanillas, B. Anti-inflammatory effects of flavonoids. Food Chem. 2019, 299, 125124. [Google Scholar] [CrossRef]
- Choy, K.W.; Murugan, D.; Leong, X.F.; Abas, R.; Alias, A.; Mustafa, M.R. Flavonoids as Natural Anti-Inflammatory Agents Targeting Nuclear Factor-Kappa B (NFkappaB) Signaling in Cardiovascular Diseases: A Mini Review. Front. Pharmacol. 2019, 10, 1295. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.F.; Liu, J.; Yang, Y.A.; Zhu, H.L. A Review: The Anti-inflammatory, Anticancer and antibacterial properties of four kinds of Licorice flavonoids isolated from Licorice. Curr. Med. Chem. 2020, 27, 1997–2011. [Google Scholar] [CrossRef]
- Singh, R.P.; Agarwal, R. Natural flavonoids targeting deregulated cell cycle progression in cancer cells. Curr. Drug Targets 2006, 7, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Romagnolo, D.F.; Selmin, O.I. Flavonoids and cancer prevention: A review of the evidence. J. Nutr. Gerontol. Geriatr. 2012, 31, 206–238. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Garcia, C.; Sanchez-Quesada, C.; Gaforio, J.J. Dietary flavonoids as cancer chemopreventive agents: An updated review of human studies. Antioxidants 2019, 8, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Ouyang, Y.Y.; Liu, J.; Zhao, G. Flavonoid intake and risk of CVD: A systematic review and meta-analysis of prospective cohort studies. Br. J. Nutr. 2014, 111, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Guerrero, J.A.; Lozano, M.L.; Castillo, J.; Benavente-Garcia, O.; Vicente, V.; Rivera, J. Flavonoids inhibit platelet function through binding to the thromboxane A2 receptor. J. Thromb. Haemost. 2005, 3, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Wang, P.; Jiao, Y.; Zhang, X.; Chen, D.; Xu, H. Hydroxysafflor Yellow A: A Systematical Review on Botanical Resources, Physicochemical Properties, Drug Delivery System, Pharmacokinetics, and Pharmacological Effects. Front. Pharmacol. 2020, 11, 579332. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.C.; Tsai, F.M.; Chen, M.L.; Wu, S.; Lee, M.C.; Tsai, T.C.; Wang, L.K.; Wang, C.H. Antipsychotic Drugs Inhibit Platelet Aggregation via P2Y 1 and P2Y 12 Receptors. Biomed Res. Int. 2016, 2016, 2532371. [Google Scholar] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, P.-H.; Kuo, C.-Y.; Chan, C.-C.; Wang, L.-K.; Chen, M.-L.; Tzeng, I.-S.; Tsai, F.-M. Safflower Extract Inhibits ADP-Induced Human Platelet Aggregation. Plants 2021, 10, 1192. https://doi.org/10.3390/plants10061192
Lu P-H, Kuo C-Y, Chan C-C, Wang L-K, Chen M-L, Tzeng I-S, Tsai F-M. Safflower Extract Inhibits ADP-Induced Human Platelet Aggregation. Plants. 2021; 10(6):1192. https://doi.org/10.3390/plants10061192
Chicago/Turabian StyleLu, Ping-Hsun, Chan-Yen Kuo, Chuan-Chi Chan, Lu-Kai Wang, Mao-Liang Chen, I-Shiang Tzeng, and Fu-Ming Tsai. 2021. "Safflower Extract Inhibits ADP-Induced Human Platelet Aggregation" Plants 10, no. 6: 1192. https://doi.org/10.3390/plants10061192
APA StyleLu, P. -H., Kuo, C. -Y., Chan, C. -C., Wang, L. -K., Chen, M. -L., Tzeng, I. -S., & Tsai, F. -M. (2021). Safflower Extract Inhibits ADP-Induced Human Platelet Aggregation. Plants, 10(6), 1192. https://doi.org/10.3390/plants10061192