
The Anti-Inflammatory and the Antinociceptive Effects of Mixed Agrimonia pilosa Ledeb. and Salvia miltiorrhiza Bunge Extract

 ,
,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Chromatographic Analysis of ME
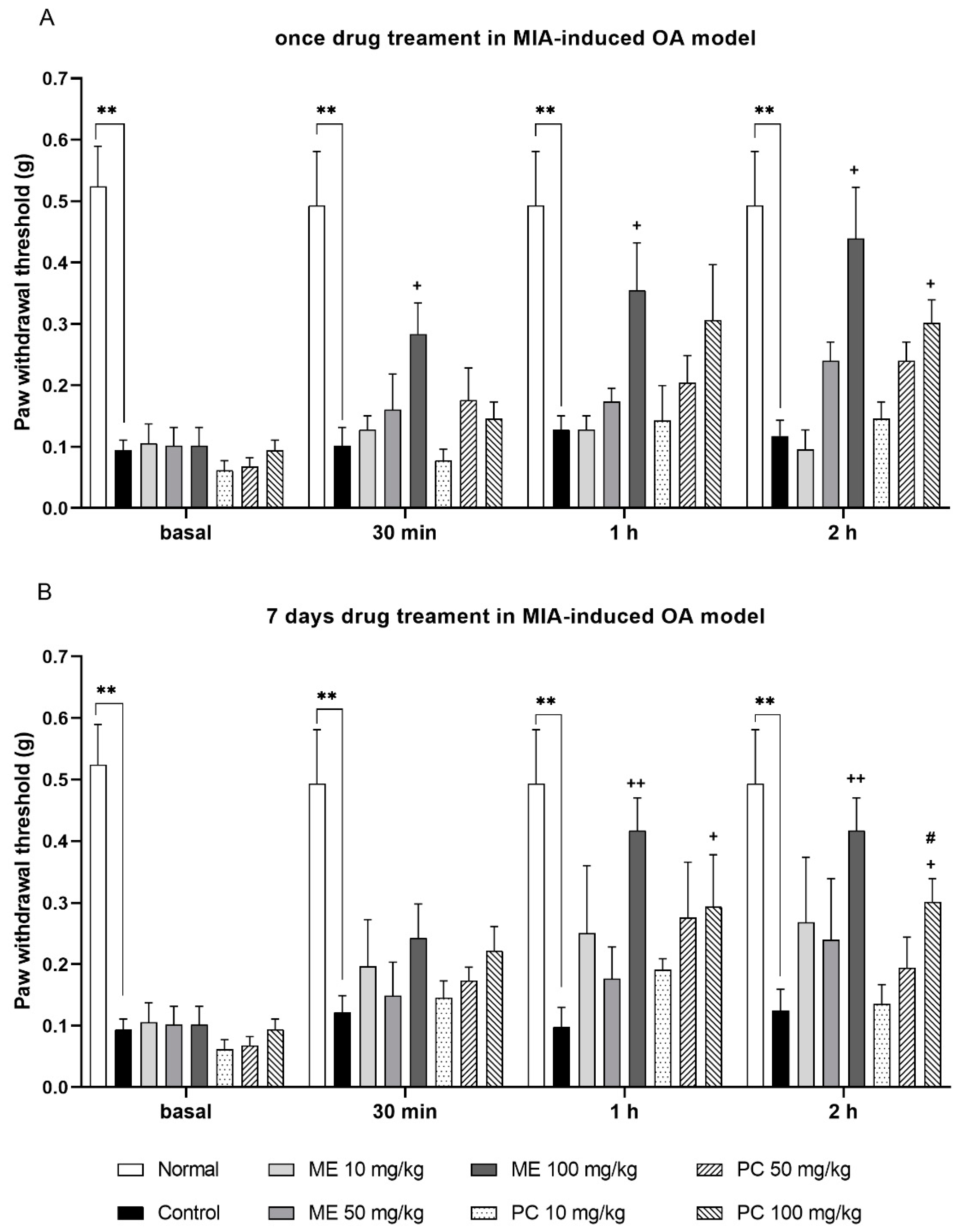
2.2. ME and PC Reduced Mechanical Pain Threshold in the MIA-Induced OA Model
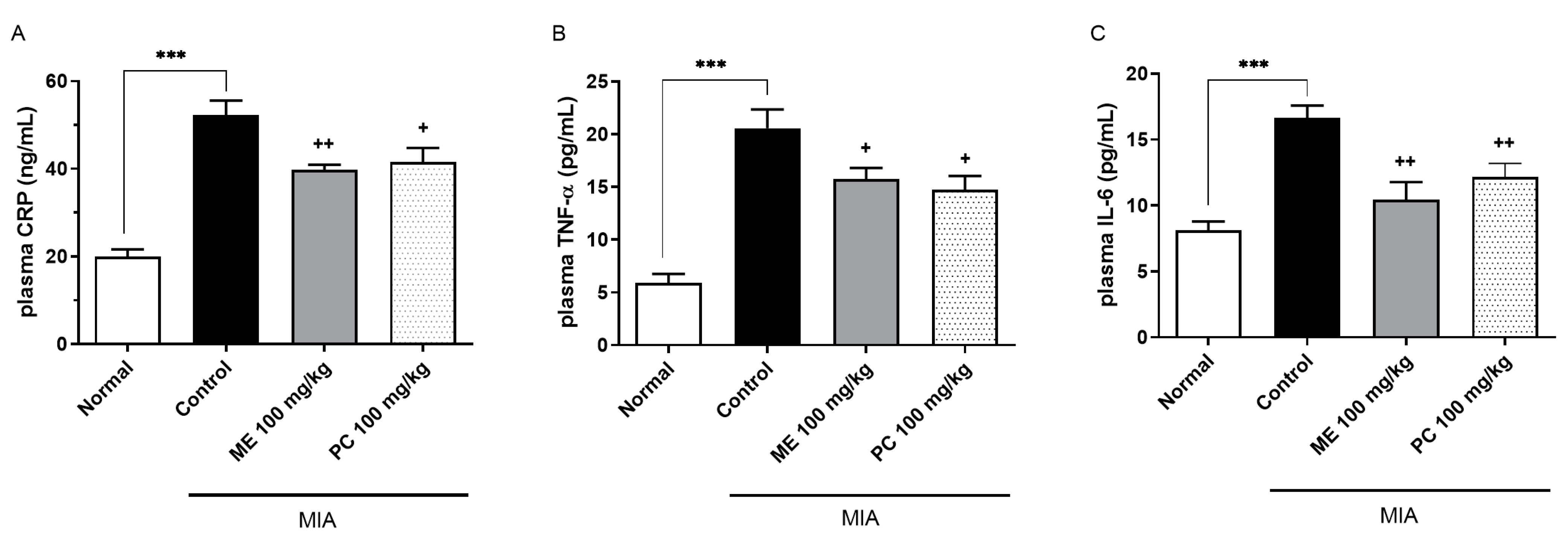
2.3. ME and PC Suppressed the Levels of Plasma CRP, TNF-α, and IL-6 in the MIA-Induced OA Model
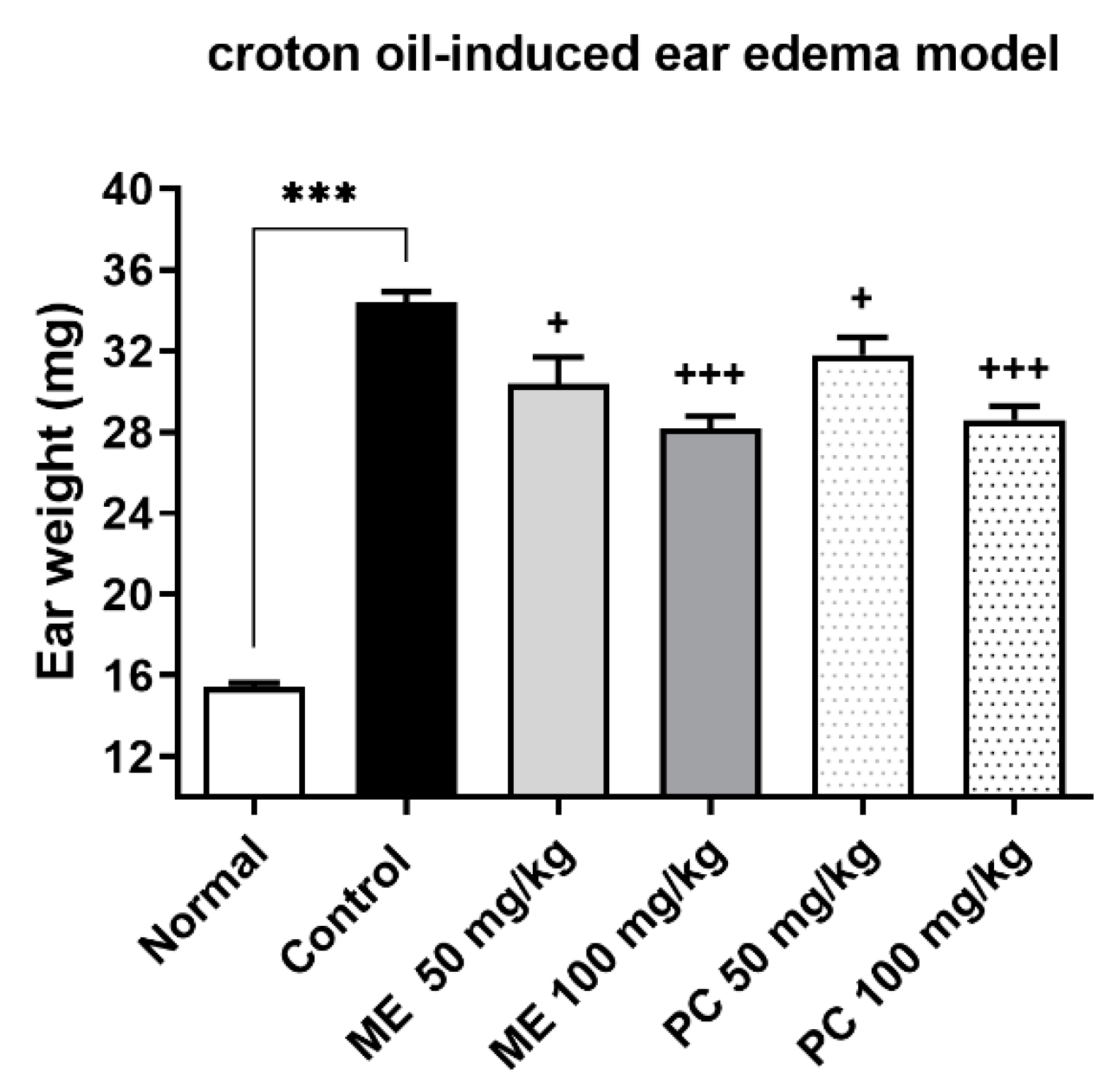
2.4. ME and PC Ameliorated Ear Edema Induced by Croton Oil
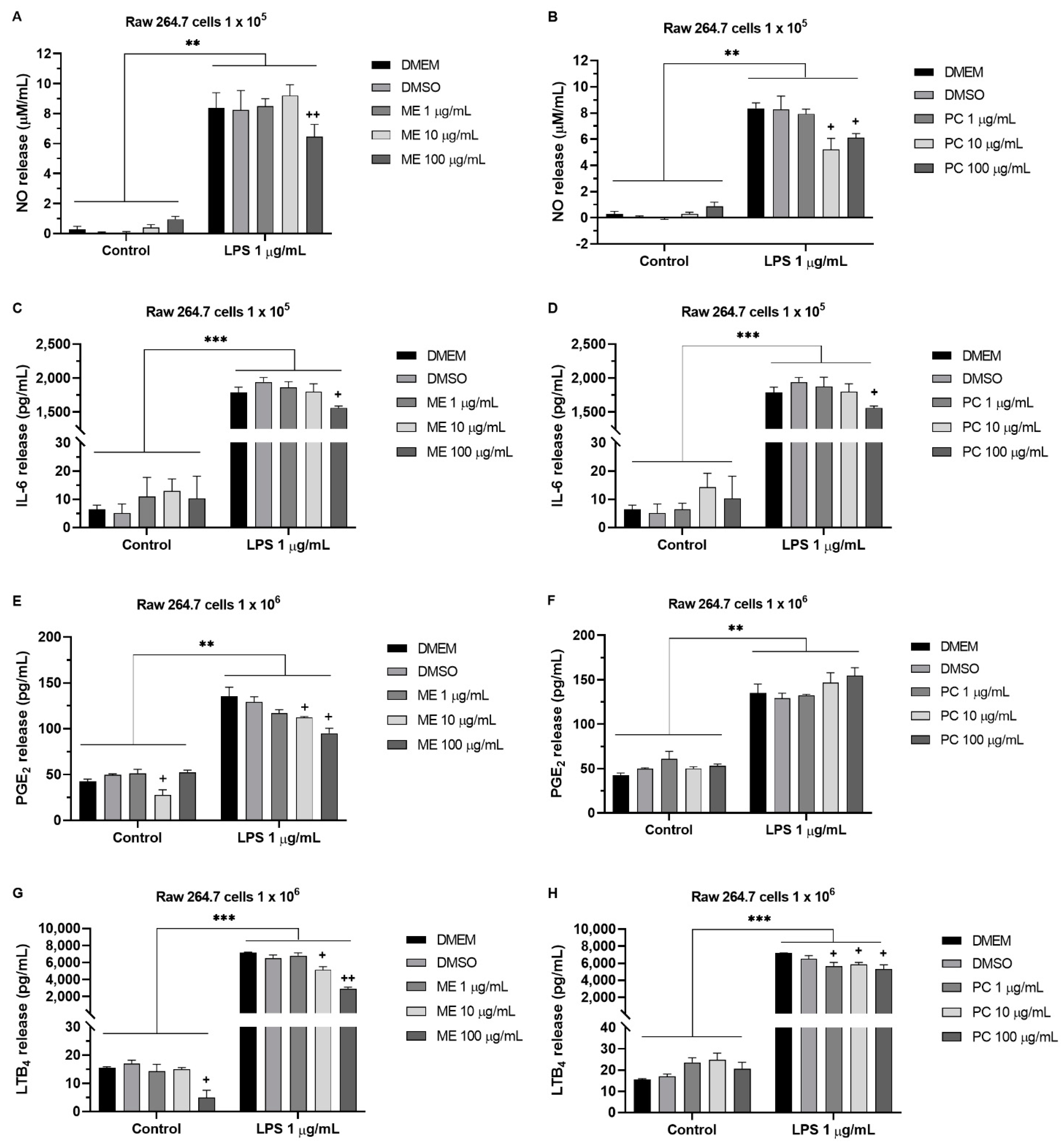
2.5. ME and PC Decreased the Nitric Oxide (NO), PGE2, LTB4, and IL-6 Release in RAW 264.7 Cells
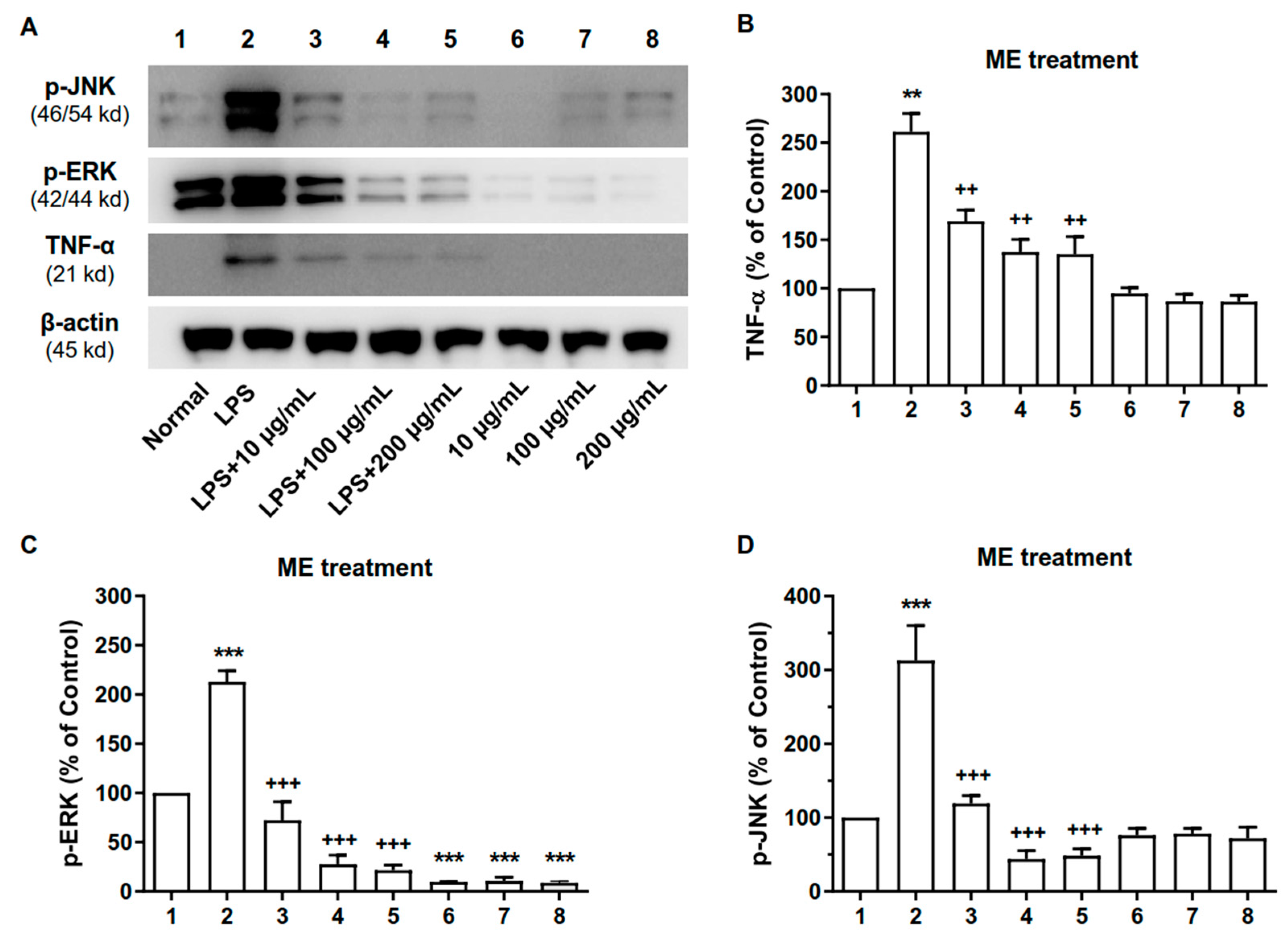
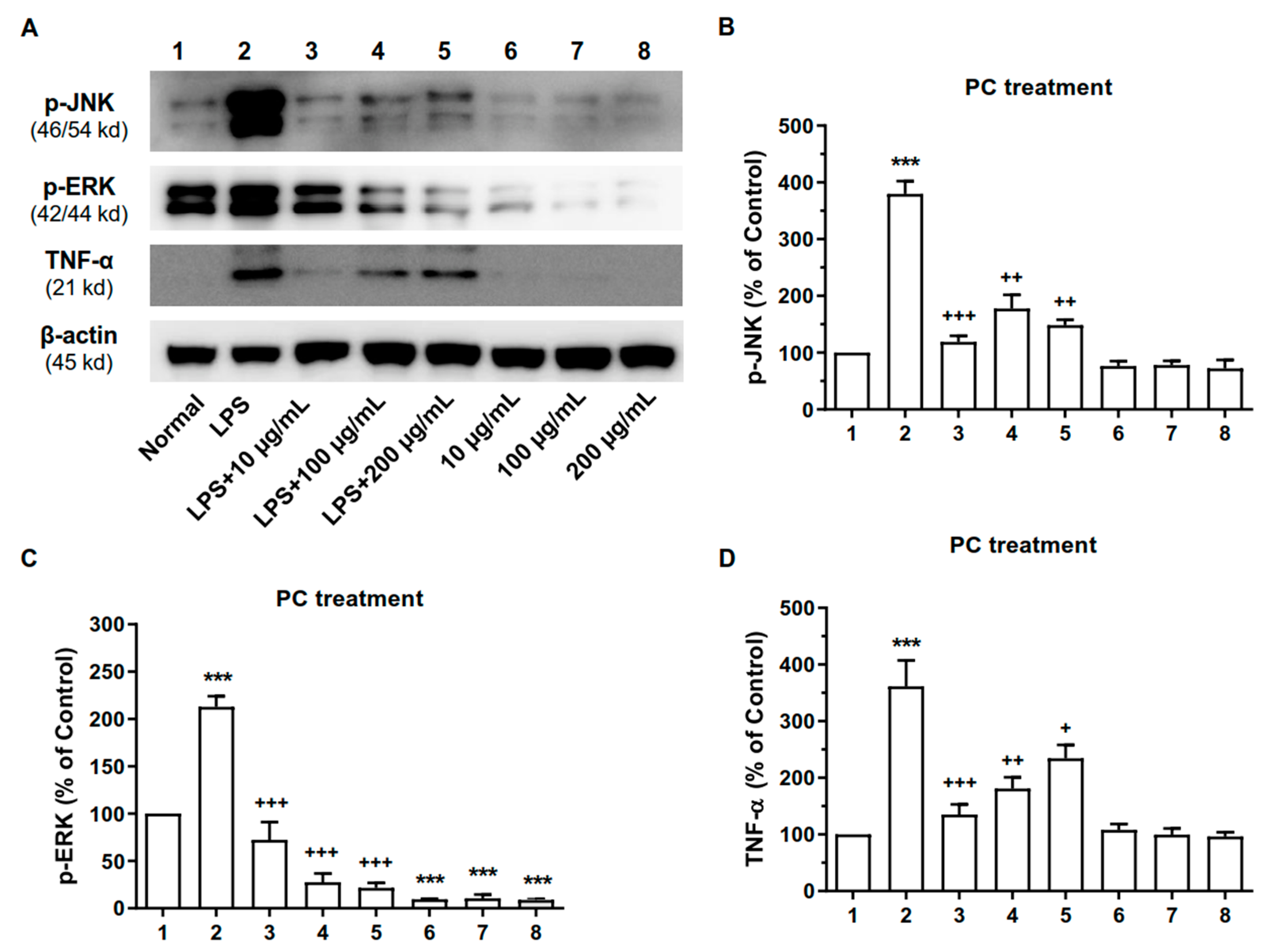
2.6. ME and PC Downregulated the Expression Levels of MAPK and TNF-α Proteins in RAW 264.7 Cells
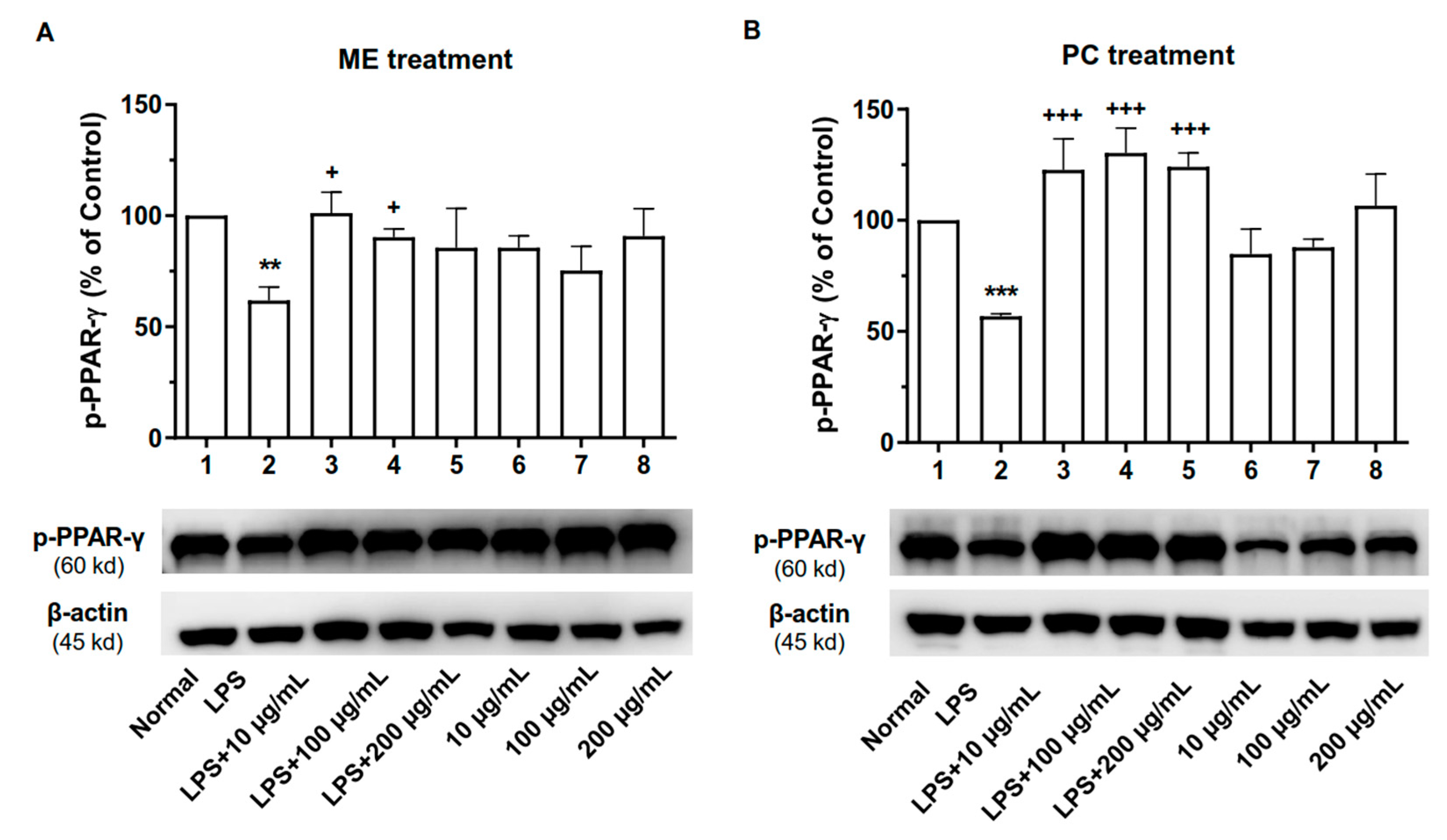
2.7. ME and PC Upregulated the Phosphorylated Level of PPAR-γ Protein in RAW 264.7 Cells
3. Discussion
4. Materials and Methods
4.1. Materials Preparation
4.2. Phytochemical Analysis Using HPLC
4.3. Experimental Animals
4.4. Production of MIA-Induced OA Model
4.5. Plasma Cytokines ELISA Assay in ICR Mice
4.6. The Measurement of Ear Edema
4.7. Cell Culture and Treatment
4.8. Measurement of Nitric Oxide
4.9. Determination of PGE2, LTB4, and IL-6 Production in the RAW 264.7 Cells
4.10. Western Blot Analysis
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wieland, H.A.; Michaelis, M.; Kirschbaum, B.J.; Rudolphi, K.A. Osteoarthritis—An untreatable disease? Nat. Rev. Drug Discov. 2005, 4, 331–344. [Google Scholar] [CrossRef]
- Goldring, M.B.; Goldring, S.R. Osteoarthritis. J. Cell Physiol. 2007, 213, 626–634. [Google Scholar] [CrossRef]
- Bijlsma, J.W.; Berenbaum, F.; Lafeber, F.P. Osteoarthritis: An update with relevance for clinical practice. Lancet 2011, 377, 2115–2126. [Google Scholar] [CrossRef]
- Kato, H.; Li, W.; Koike, M.; Wang, Y.; Koike, K. Phenolic glycosides from Agrimonia pilosa. Phytochemistry 2010, 71, 1925–1929. [Google Scholar] [CrossRef]
- Zhu, L.; Tan, J.; Wang, B.; He, R.; Liu, Y.; Zheng, C. Antioxidant activities of aqueous extract from Agrimonia pilosa Ledeb and its fractions. Chem. Biodivers. 2009, 6, 1716–1726. [Google Scholar] [CrossRef]
- Feng, J.H.; Lee, H.J.; Kim, S.B.; Jung, J.S.; Lim, S.S.; Suh, H.W. Antinociceptive Effect of Single Components Isolated from Agrimonia pilosa Ledeb. Extract. Sci. Pharm. 2019, 87, 18. [Google Scholar] [CrossRef] [Green Version]
- Park, S.H.; Sim, Y.B.; Kang, Y.J.; Lee, J.K.; Lim, S.S.; Suh, H.W. Effect of Agrimonia pilosa Ledeb extract on the antinociception and mechanisms in mouse. Korean J. Physiol. Pharmacol. 2012, 16, 119–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.Y.; Yu, Q.M.; Kong, H.J.; Lee, J.Y.; Yang, K.M.; Seo, J.S. Antioxidant and anti-inflammatory activities of Agrimonia pilosa Ledeb. extract. Evid. Based Complement. Alternat. Med. 2020, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.J.; Jiang, J.; Shim, D.W.; Kwon, S.C.; Kim, T.J.; Ye, S.K.; Kim, M.K.; Shin, Y.K.; Koppula, S.; Kang, T.B. Anti-inflammatory and anti-allergic effects of Agrimonia pilosa Ledeb extract on murine cell lines and OVA-induced airway inflammation. J. Ethnopharmacol. 2012, 140, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.R.; Jeon, H.; Kwon, J.E.; Suh, H.; Kim, B.H.; Yun, M.K.; Lim, Y.J.; Kang, S.C. Anti-osteoporotic effects of Salvia miltiorrhiza Bunge EtOH extract both in ovariectomized and naturally menopausal mouse models. J. Ethnopharmacol. 2020, 258, 112874. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Zuo, Z.; Chow, M.S.S. Danshen: An overview of its chemistry, pharmacology, pharmacokinetics, and clinical use. J. Clin. Pharmacol. 2005, 45, 1345–1359. [Google Scholar] [CrossRef]
- Guo, Y.; Li, Y.; Xue, L.; Severino, R.P.; Gao, S.; Niu, J.; Qin, L.P.; Zhang, D.; Brömme, D. Salvia miltiorrhiza: An ancient Chinese herbal medicine as a source for anti-osteoporotic drugs. J. Ethnopharmacol. 2014, 155, 1401–1416. [Google Scholar] [CrossRef]
- Hwang, S.H.; Kim, S.B.; Jang, S.-P.; Wang, Z.; Suh, H.W.; Lim, S.S. Anti-Nociceptive Effect and Standardization from Mixture of Agrimonia pilosa Ledeb and Salvia miltiorrhiza Bunge Extracts. J. Med. Food 2018, 21, 596–604. [Google Scholar] [CrossRef]
- Abramson, S.B. Osteoarthritis and nitric oxide. Osteoarthr. Cartil. 2008, 16, S15–S20. [Google Scholar] [CrossRef] [Green Version]
- Goldring, M.B.; Otero, M. Inflammation in osteoarthritis. Curr. Opin. Rheumatol. 2011, 23, 471. [Google Scholar] [CrossRef] [PubMed]
- Choy, E.H.; Panayi, G.S. Cytokine pathways and joint inflammation in rheumatoid arthritis. N. Engl. J. Med. 2001, 344, 907–916. [Google Scholar] [CrossRef]
- McInnes, I.B.; Schett, G. Cytokines in the pathogenesis of rheumatoid arthritis. Nat. Rev. Immunol. 2007, 7, 429–442. [Google Scholar] [CrossRef]
- Libby, P. Role of inflammation in atherosclerosis associated with rheumatoid arthritis. Am. J. Med. 2008, 121, S21–S31. [Google Scholar] [CrossRef]
- Chen, L.; Teng, H.; Fang, T.; Xiao, J. Agrimonolide from Agrimonia pilosa suppresses inflammatory responses through down-regulation of COX-2/iNOS and inactivation of NF-κB in lipopolysaccharide-stimulated macrophages. Phytomedicine 2016, 23, 846–855. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.G.; Tran, P.T.; Lee, J.H.; Min, B.S.; Kim, J.A. Anti-inflammatory activity of caffeic acid derivatives isolated from the roots of Salvia miltiorrhiza Bunge. Arch. Pharm. Res. 2018, 41, 64–70. [Google Scholar] [CrossRef]
- Ngo, T.M.; Tran, P.T.; Hoang, L.S.; Lee, J.H.; Min, B.S.; Kim, J.A. Diterpenoids isolated from the root of Salvia miltiorrhiza and their anti-inflammatory activity. Nat. Product Res. 2019, 35, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Song, S.; Wang, J.; Zhang, Q.; Qiu, F.; Zhao, F. Tiliroside, the major component of Agrimonia pilosa Ledeb ethanol extract, inhibits MAPK/JNK/p38-mediated inflammation in lipopolysaccharide-activated RAW 264.7 macrophages. Exp. Ther. Med. 2016, 12, 499–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, T.; Zhu, L.; Tan, J.; Zhou, X.; Xiao, L.; Liu, X.; Wang, B. Promoting effect of triterpenoid compound from Agrimonia pilosa Ledeb on preadipocytes differentiation via up-regulation of PPAR-γ expression. Pharmacogn. Mag. 2015, 11, 219. [Google Scholar]
- Eason, C.T.; Adams, S.L.; Puddick, J.; Romanazzi, D.; Miller, M.R.; King, N.; Johns, S.; Forbes-Blom, E.; Hessian, P.A.; Stamp, L.K. Greenshell™ Mussels: A Review of Veterinary Trials and Future Research Directions. Vet. Sci. 2018, 5, 36. [Google Scholar] [CrossRef] [Green Version]
- Siriarchavatana, P.; Kruger, M.C.; Miller, M.R.; Tian, H.S.; Wolber, F.M. The Preventive Effects of Greenshell Mussel (Perna canaliculus) on Early-Stage Metabolic Osteoarthritis in Rats with Diet-Induced Obesity. Nutrients 2019, 11, 1601. [Google Scholar] [CrossRef] [Green Version]
- Heming, M.; Gran, S.; Jauch, S.L.; Fischer-Riepe, L.; Russo, A.; Klotz, L.; Hermann, S.; Schäfers, M.; Roth, J.; Barczyk-Kahlert, K. Peroxisome proliferator-activated receptor-γ modulates the response of macrophages to lipopolysaccharide and glucocorticoids. Front. Immunol. 2018, 9, 893. [Google Scholar] [CrossRef]
- Lindler, B.N.; Long, K.E.; Taylor, N.A.; Lei, W. Use of Herbal Medications for Treatment of Osteoarthritis and Rheumatoid Arthritis. Medicines 2020, 7, 67. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Huang, L.; Ding, F.; Yang, K.; Feng, Y.; Tang, H.; Xu, Q.M.; Feng, J.; Yang, S. Simultaneous purification of dihydrotanshinone, tanshinone I, cryptotanshinone, and tanshinone IIA from Salvia miltiorrhiza and their anti-inflammatory activities investigation. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Federica, R.; Anella, S.; Claudia, C.; Marco, C.G.; Antonia, S.; Sonia, L.; Irene, D.; Giovanna, C.M.; Giuseppe, B.; Antonio, C. Pharmacological and molecular docking assessment of cryptotanshinone as natural-derived analgesic compound. Biomed. Pharmacother. 2020, 126, 110042. [Google Scholar]
- Zhang, W.; Suo, M.; Yu, G.; Zhang, M. Antinociceptive and anti-inflammatory effects of cryptotanshinone through PI3K/Akt signaling pathway in a rat model of neuropathic pain. Chem. Biol. Interact. 2019, 305, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Ri-Ge-le, A.; Guo, Z.L.; Wang, Q.; Zhang, B.-J.; Kong, D.W.; Yang, W.Q.; Yu, Y.B.; Zhang, L. Tanshinone IIA improves painful diabetic neuropathy by suppressing the expression and activity of voltage-gated Sodium Channel in rat dorsal root ganglia. Exp. Clin. Endocrinol. Diabetes 2018, 126, 632–639. [Google Scholar] [CrossRef]
- Mannelli, L.D.C.; Piccolo, M.; Maione, F.; Ferraro, M.G.; Irace, C.; De Feo, V.; Ghelardini, C.; Mascolo, N. Tanshinones from Salvia miltiorrhiza Bunge revert chemotherapy-induced neuropathic pain and reduce glioblastoma cells malignancy. Biomed. Pharmacother. 2018, 105, 1042–1049. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Lv, H.; Li, X.; Su, H.; Zhang, X.; Yang, J. Danshen attenuates cartilage injuries in osteoarthritis in vivo and in vitro by activating JAK2/STAT3 and AKT pathways. Exp. Anim. 2018, 67, 127–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Wang, J.; Ju, J.; Dai, J. Salvianolic acid B and sodium tanshinone II A sulfonate prevent pulmonary fibrosis through anti-inflammatory and anti-fibrotic process. Eur. J. Pharmacol. 2020, 883, 173352. [Google Scholar] [CrossRef]
- Yi, Y.S. Regulatory roles of flavonoids on inflammasome activation during inflammatory responses. Mol. Nutr. Food Res. 2018, 62, 1800147. [Google Scholar] [CrossRef]
- Kim, S.B.; Hwang, S.H.; Suh, H.W.; Lim, S.S. Phytochemical analysis of Agrimonia pilosa Ledeb, its antioxidant activity and aldose reductase inhibitory potential. Int. J. Mol. Sci. 2017, 18, 379. [Google Scholar] [CrossRef]
- Rylski, M.; Duriasz-Rowińska, H.; Rewerski, W. The analgesic action of some flavonoids in the hot plate test. Acta Physiol. Pol. 1979, 30, 385–388. [Google Scholar] [PubMed]
- Selvaraj, G.; Kaliamurthi, S.; Thirungnasambandam, R.; Vivekanandan, L.; Balasubramanian, T. Anti-nociceptive effect in mice of thillai flavonoid rutin. Biomed. Environ. Sci. 2014, 27, 295–299. [Google Scholar]
- Gul, A.; Kunwar, B.; Mazhar, M.; Faizi, S.; Ahmed, D.; Shah, M.R.; Simjee, S.U. Rutin and rutin-conjugated gold nanoparticles ameliorate collagen-induced arthritis in rats through inhibition of NF-κB and iNOS activation. Int. Immunopharmacol. 2018, 59, 310–317. [Google Scholar] [CrossRef]
- Xu, X.; Lv, H.; Li, X.; Su, H.; Zhang, X.; Yang, J. Danshen attenuates osteoarthritis-related cartilage degeneration through inhibition of NF-κB signaling pathway in vivo and in vitro. Biochem. Cell Biol. 2017, 95, 644–651. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.B.; Yuan, Y.J.; Zhang, Q.H.; Li, H.; Dai, J.L.; Min, J.K. Salvianolic acid B suppresses inflammatory mediator levels by downregulating NF-κB in a rat model of rheumatoid arthritis. Med. Sci. Monit. 2018, 24, 2524. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.H.; Butt, Y.K.C.; Wong, M.S.; Lo, S.C.L. A lipid extract of Perna canaliculus affects the expression of pro-inflammatory cytokines in a rat adjuvant-induced arthritis model. Eur. Ann. Allergy Clin. Immunol. 2008, 40, 148. [Google Scholar]
- Lee, C.H.; Lum, J.H.K.; Ng, C.K.C.; McKay, J.; Butt, Y.K.C.; Wong, M.S.; Lo, S.C.L. Pain controlling and cytokine-regulating effects of lyprinol, a lipid extract of Perna canaliculus, in a rat adjuvant-induced arthritis model. Evid. Based Complement Alternat. 2009, 6, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Coulson, S.; Palacios, T.; Vitetta, L. Perna canaliculus (green-lipped mussel): Bioactive components and therapeutic evaluation for chronic health conditions. Prog. Drug Res. 2015, 70, 91–132. [Google Scholar]
- Lawson, B.R.; Belkowski, S.M.; Whitesides, J.F.; Davis, P.; Lawson, J.W. Immunomodulation of murine collagen-induced arthritis by N, N-dimethylglycine and a preparation of Perna canaliculus. BMC Complement Altern. Med. 2007, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Arthur, J.S.C.; Ley, S.C. Mitogen-activated protein kinases in innate immunity. Nat. Rev. Immunol. 2013, 13, 679–692. [Google Scholar] [CrossRef]
- Ni, L.; Zheng, Y.; Gong, T.; Xiu, C.; Li, K.; Li, B.; Yang, H.; Chen, J. Proinflammatory macrophages promote degenerative phenotypes in rat nucleus pulpous cells partly through ERK and JNK signaling. J. Cell Physiol. 2019, 234, 5362–5371. [Google Scholar] [CrossRef]
- Shin, M.J.; Kim, D.W.; Choi, Y.J.; Cha, H.J.; Lee, S.H.; Lee, S.; Park, J.; Han, K.H.; Eum, W.S.; Choi, S.Y. PEP-1-GLRX1 protein exhibits anti-inflammatory effects by inhibiting the activation of MAPK and NF-κB pathways in Raw 264.7 cells. BMB Rep. 2020, 53, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, H.; Liu, X.; Sun, W.; Kang, N.; Liu, Y.; Yang, S.; Xu, Q.M.; Wang, C.; Chen, X. Total tanshinones exhibits anti-inflammatory effects through blocking TLR4 dimerization via the MyD88 pathway. Cell Death Dis. 2017, 8, e3004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, R.; Huang, L.; Du, L.J.; Feng, J.F.; Li, J.; Luo, Y.Y.; Xu, Q.M.; Yang, S.L.; Gao, H.; Feng, Y.L. Dihydrotanshinone exhibits an anti-inflammatory effect in vitro and in vivo through blocking TLR4 dimerization. Pharmacol. Res. 2019, 142, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.H.; Kim, S.B.; Lee, H.J.; Sim, S.M.; Lim, S.S.; Suh, H.W. Effects of Salvia miltiorrhiza Bunge extract and its single components on monosodium urate-induced pain in vivo and lipopolysaccharide-induced inflammation in vitro. J. Tradit Chin. Med. 2021, 41, 219–226. [Google Scholar] [PubMed]
- Hong, J.S.; Feng, J.H.; Park, J.S.; Lee, H.J.; Lee, J.Y.; Lim, S.S.; Suh, H.W. Antinociceptive effect of chrysin in diabetic neuropathy and formalin-induced pain models. Anim. Cells Syst. 2020, 24, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Bonin, R.P.; Bories, C.; De Koninck, Y. A simplified up-down method (SUDO) for measuring mechanical nociception in rodents using von Frey filaments. Mol. Pain 2014, 10, 26. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Namgoong, S.Y.; Kim, H.P. Antiinflammatory activity of flavonoids: Mouse ear edema inhibition. Arch. Pharm. Res. 1993, 16, 18. [Google Scholar] [CrossRef]
- Li, C.Y.; Meng, Y.H.; Ying, Z.M.; Xu, N.; Hao, D.; Gao, M.Z.; Zhang, W.J.; Xu, L.; Gao, Y.C.; Ying, X.X. Three novel alkaloids from Portulaca oleracea L. and their anti-inflammatory effects. J. Agric. Food Chem. 2016, 64, 5837–5844. [Google Scholar] [CrossRef] [PubMed]
- Damodar, K.; Lee, J.T.; Kim, J.K.; Jun, J.G. Synthesis and in vitro evaluation of homoisoflavonoids as potent inhibitors of nitric oxide production in RAW-264.7 cells. Bioorg. Med. Chem. Lett. 2018, 28, 2098–2102. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, J.-H.; Kim, H.-Y.; Sim, S.-M.; Zuo, G.-L.; Jung, J.-S.; Hwang, S.-H.; Kwak, Y.-G.; Kim, M.-J.; Jo, J.-H.; Kim, S.-C.; et al. The Anti-Inflammatory and the Antinociceptive Effects of Mixed Agrimonia pilosa Ledeb. and Salvia miltiorrhiza Bunge Extract. Plants 2021, 10, 1234. https://doi.org/10.3390/plants10061234
Feng J-H, Kim H-Y, Sim S-M, Zuo G-L, Jung J-S, Hwang S-H, Kwak Y-G, Kim M-J, Jo J-H, Kim S-C, et al. The Anti-Inflammatory and the Antinociceptive Effects of Mixed Agrimonia pilosa Ledeb. and Salvia miltiorrhiza Bunge Extract. Plants. 2021; 10(6):1234. https://doi.org/10.3390/plants10061234
Chicago/Turabian StyleFeng, Jing-Hui, Hyun-Yong Kim, Su-Min Sim, Guang-Lei Zuo, Jeon-Sub Jung, Seung-Hwan Hwang, Youn-Gil Kwak, Min-Jung Kim, Jeong-Hun Jo, Sung-Chan Kim, and et al. 2021. "The Anti-Inflammatory and the Antinociceptive Effects of Mixed Agrimonia pilosa Ledeb. and Salvia miltiorrhiza Bunge Extract" Plants 10, no. 6: 1234. https://doi.org/10.3390/plants10061234
APA StyleFeng, J. -H., Kim, H. -Y., Sim, S. -M., Zuo, G. -L., Jung, J. -S., Hwang, S. -H., Kwak, Y. -G., Kim, M. -J., Jo, J. -H., Kim, S. -C., Lim, S. -S., & Suh, H. -W. (2021). The Anti-Inflammatory and the Antinociceptive Effects of Mixed Agrimonia pilosa Ledeb. and Salvia miltiorrhiza Bunge Extract. Plants, 10(6), 1234. https://doi.org/10.3390/plants10061234