
Morpho-Physiological Responses of Arabidopsis thaliana L. to the LED-Sourced CoeLux® System

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Results
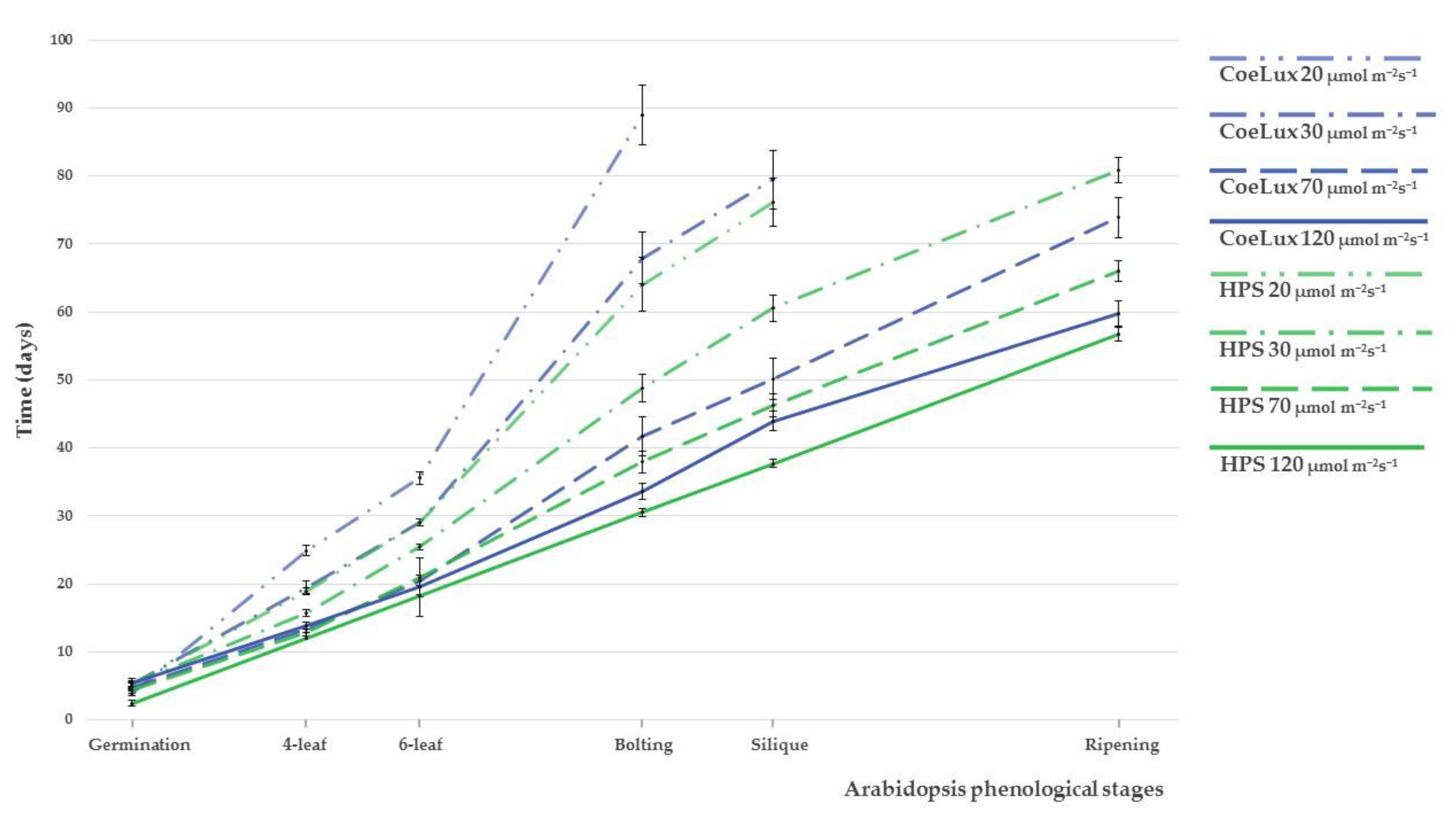
2.1. Phenological Analysis
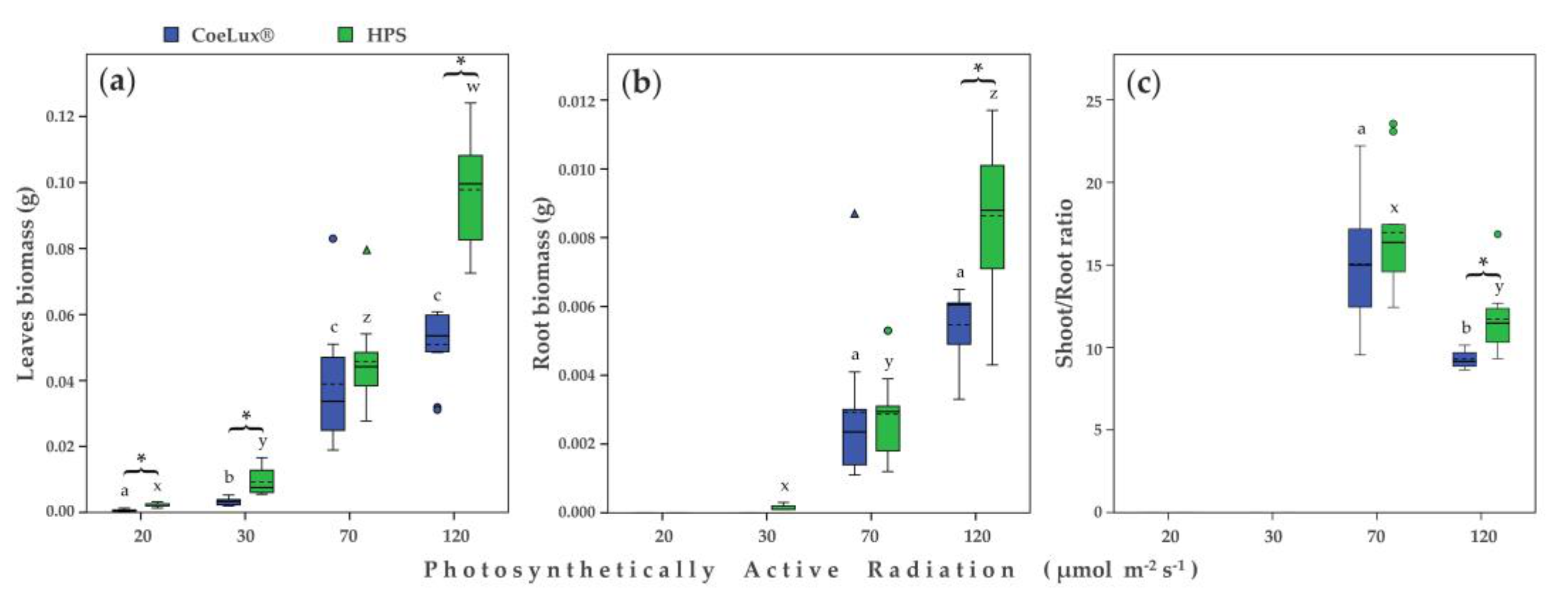
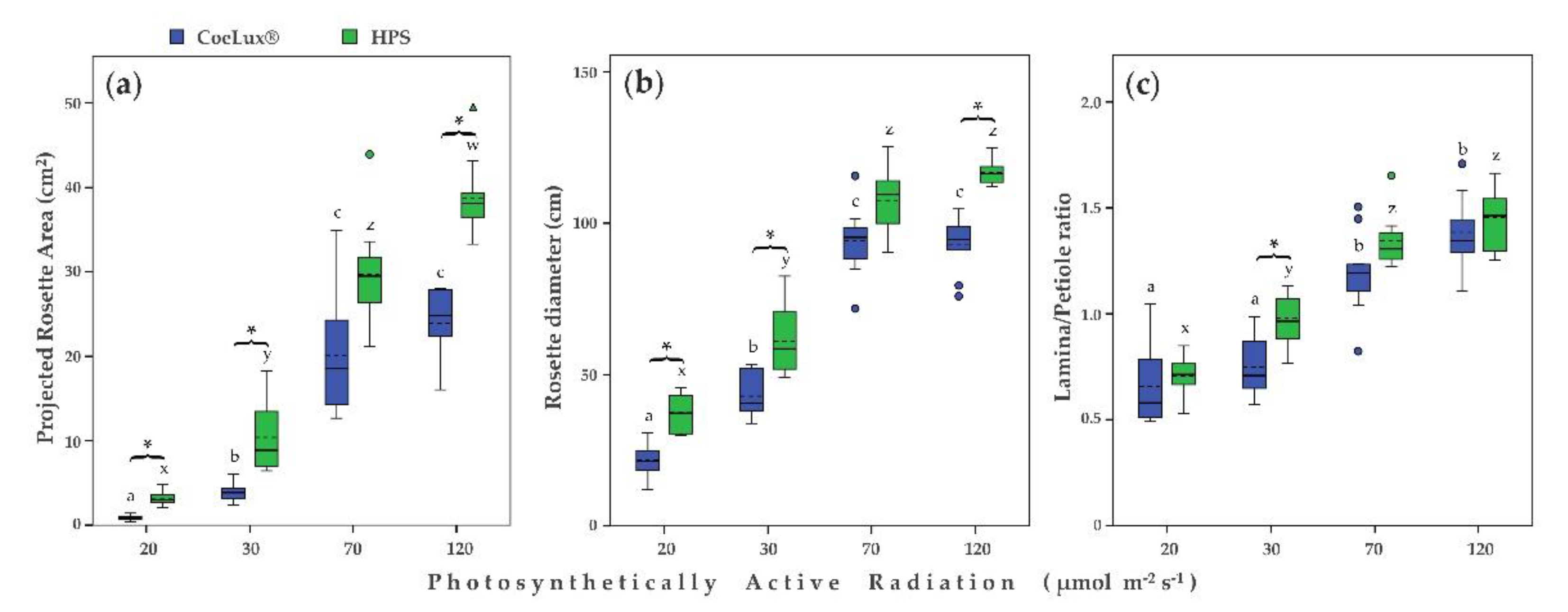
2.2. Morphological Traits
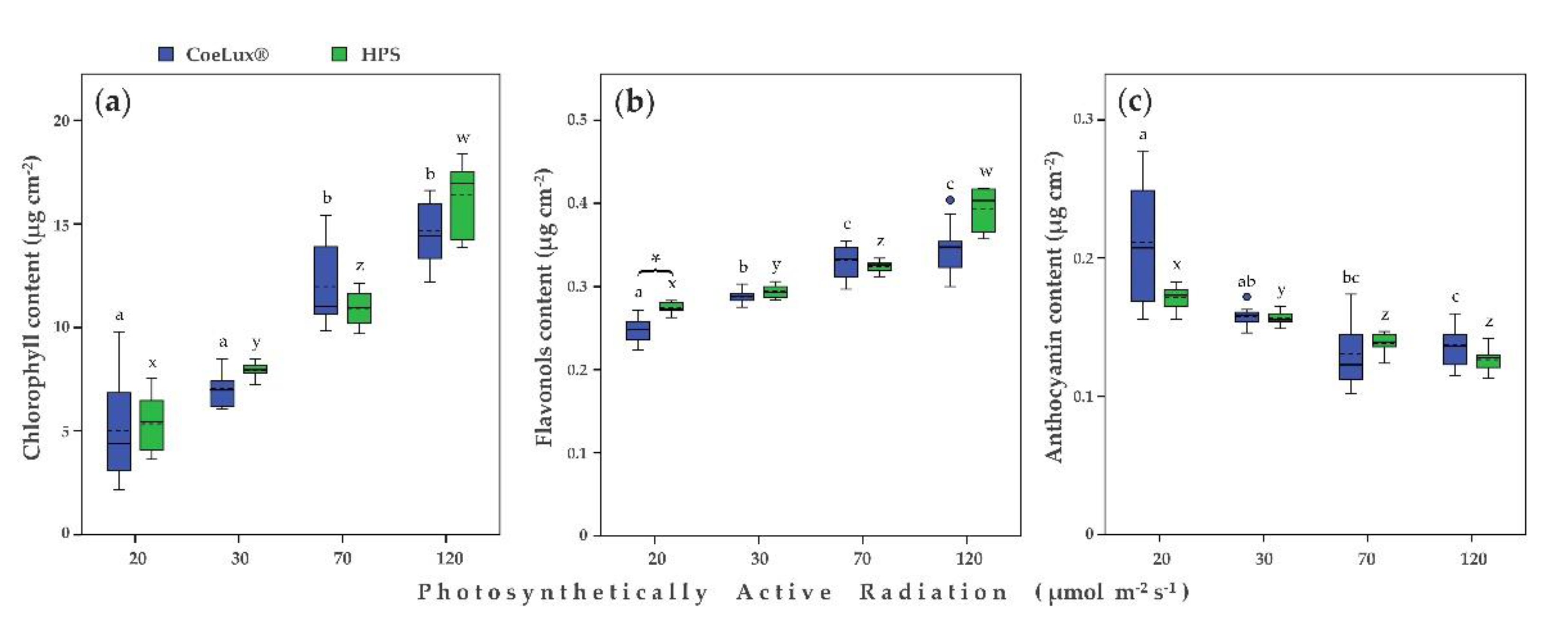
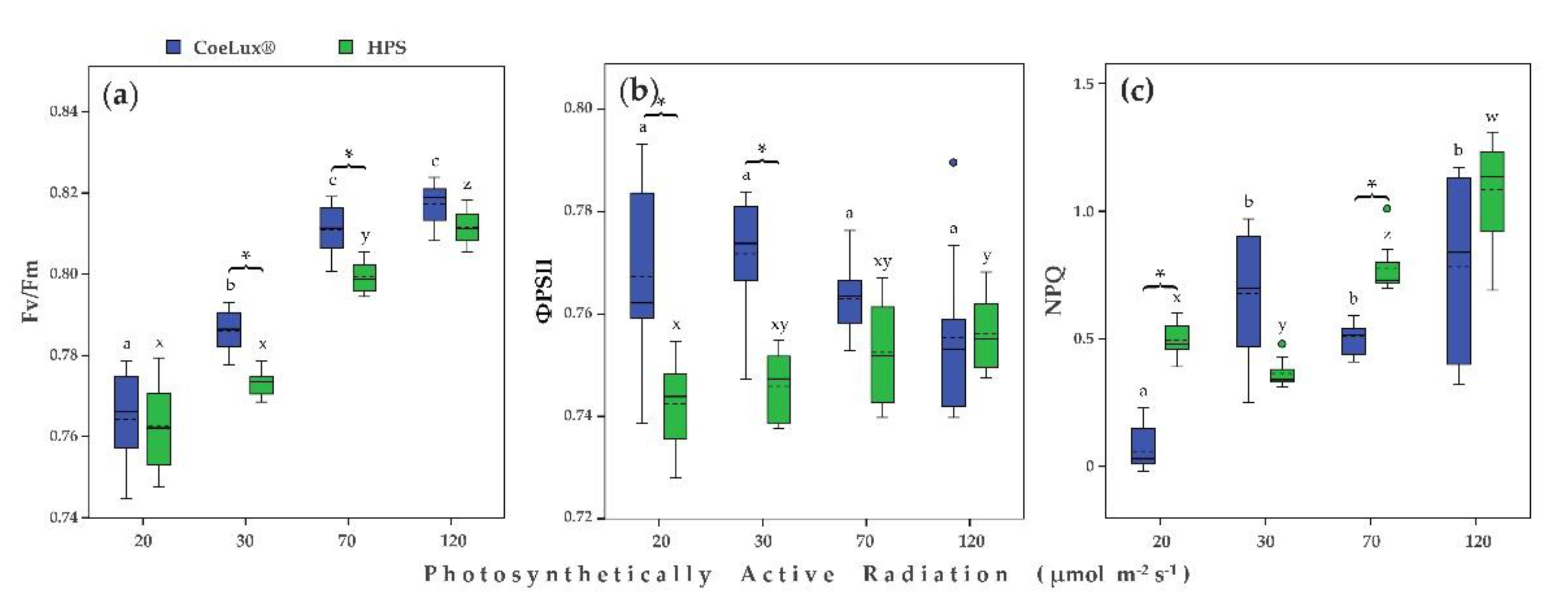
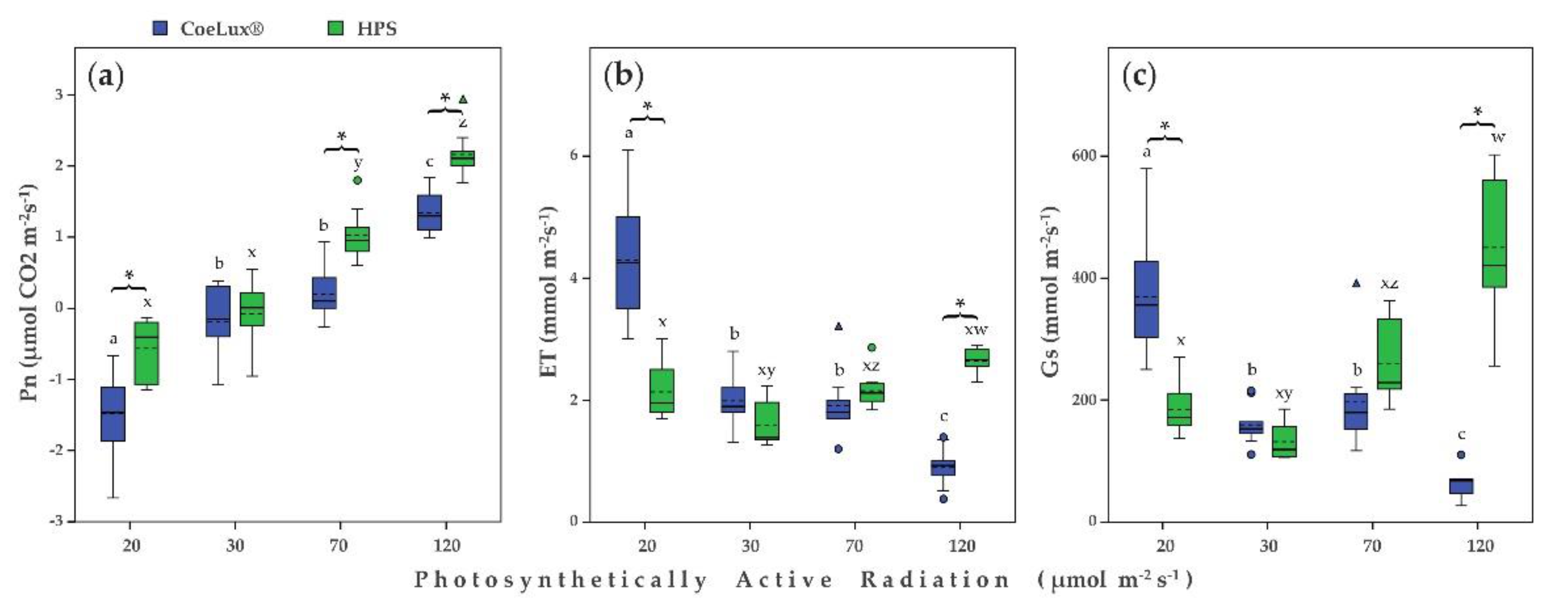
2.3. Physiological Measurements
3. Discussion
4. Materials and Methods
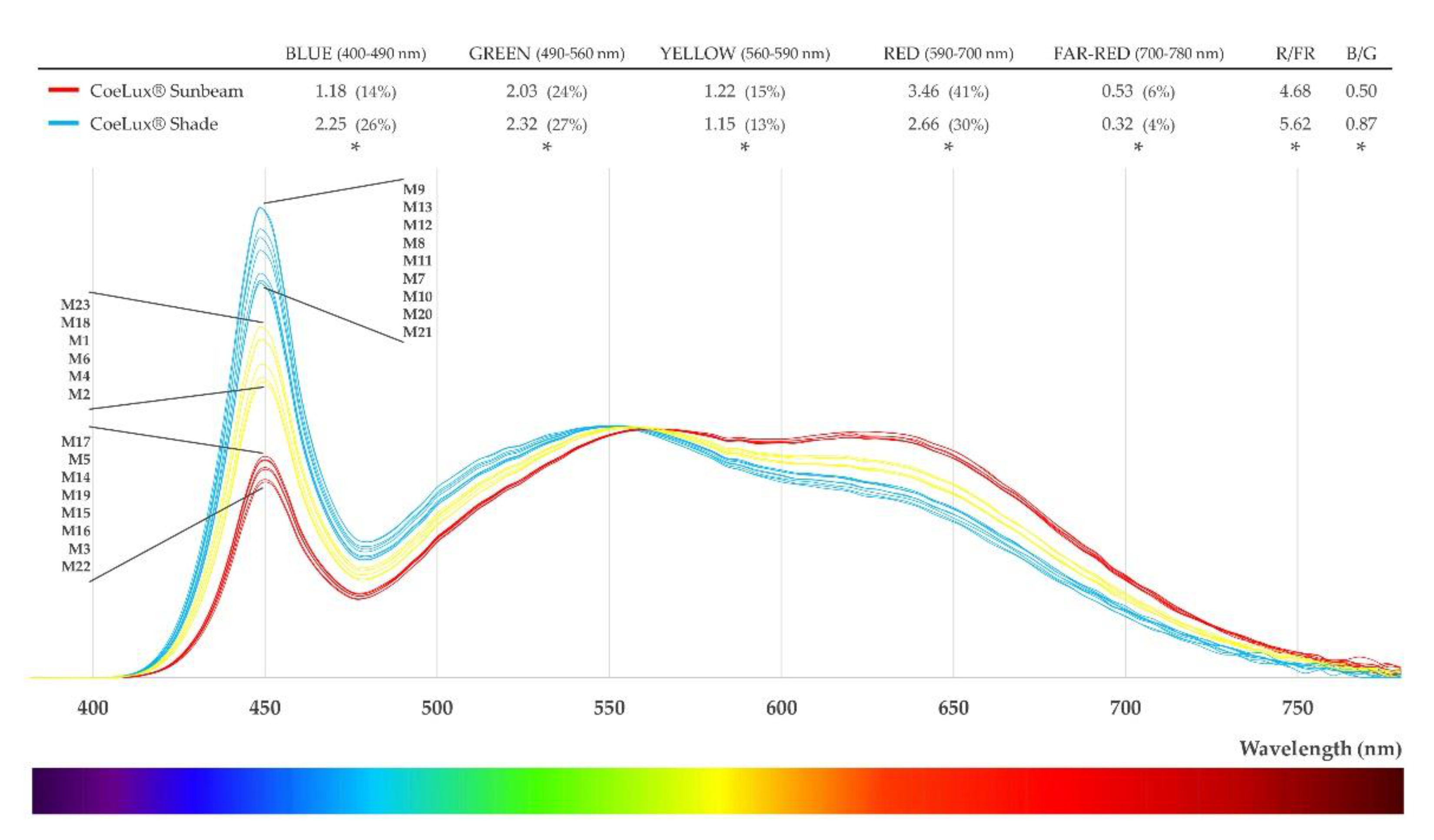
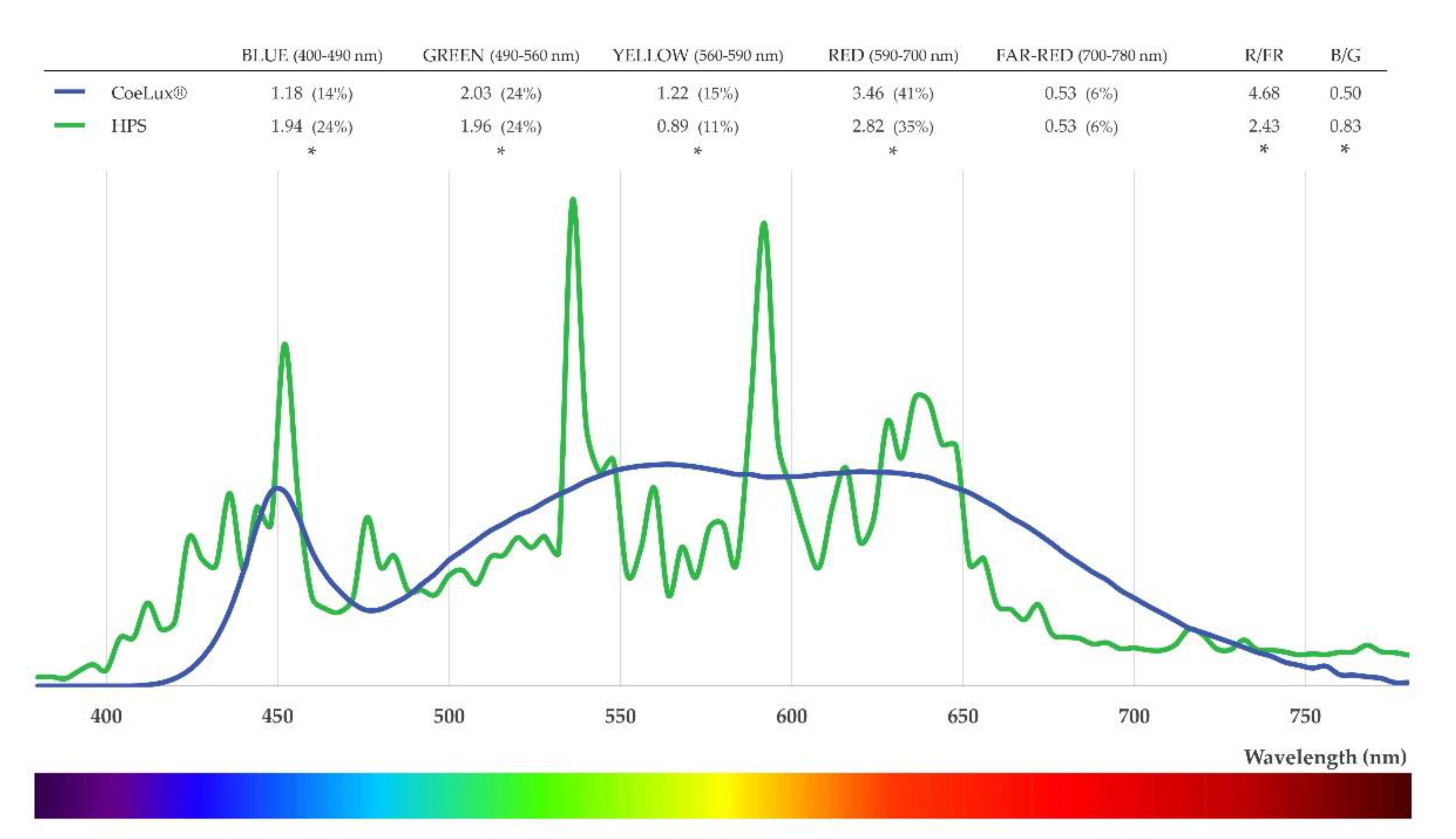
4.1. Light Characterization
4.2. Plant Material and Growth Conditions
4.3. Plant Analysis
4.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dutta Gupta, S.; Agarwal, A. Artificial Lighting System for Plant Growth and Development: Chronological Advancement, Working Principles, and Comparative Assessment. In Light Emitting Diodes for Agriculture: Smart Lighting; Springer: Singapore, 2017; pp. 1–334. ISBN 9789811058073. [Google Scholar]
- Seiler, F.; Soll, J.; Bölter, B. Comparative phenotypical and molecular analyses of arabidopsis grown under fluorescent and LED Light. Plants 2017, 6, 24. [Google Scholar] [CrossRef] [Green Version]
- Tetri, E.; Sarvaranta, A.; Syri, S. Potential of new lighting technologies in reducing household lighting energy use and CO2 emissions in Finland. Energy Effic. 2014, 7, 559–570. [Google Scholar] [CrossRef]
- Di Trapani, P.; Magatti, D. Artificial Lighting System for Simulating Natural Lighting. US Patent 2014/01,331,125 A1, 15 May 2014. [Google Scholar]
- Rayleigh, J.W. On the instability of jets. Proc. Lond. Math. Soc. 1878, 10, 4–13. [Google Scholar] [CrossRef] [Green Version]
- Di-Trapani, P.; Magatti, D. Artificial Illumination Device. US Patent 9,709,245 B2, 18 July 2017. [Google Scholar]
- Canazei, M.; Laner, M.; Staggl, S.; Pohl, W.; Ragazzi, P.; Magatti, D.; Martinelli, E.; Di Trapani, P. Room- and illumination-related effects of an artificial skylight. Light. Res. Technol. 2016, 48, 539–558. [Google Scholar] [CrossRef]
- Dong, C.; Fu, Y.; Liu, G.; Liu, H. Low light intensity effects on the growth, photosynthetic characteristics, antioxidant capacity, yield and quality of wheat (Triticum aestivum L.) at different growth stages in BLSS. Adv. Sp. Res. 2014, 53, 1557–1566. [Google Scholar] [CrossRef]
- Bringslimark, T.; Hartig, T.; Patil, G.G. The psychological benefits of indoor plants: A critical review of the experimental literature. J. Environ. Psychol. 2009, 29, 422–433. [Google Scholar] [CrossRef]
- Montagnoli, A.; Dumroese, R.K.; Terzaghi, M.; Pinto, J.R.; Fulgaro, N.; Scippa, G.S.; Chiatante, D. Tree seedling response to LED spectra: Implications for forest restoration. Plant Biosyst. 2018, 152, 515–523. [Google Scholar] [CrossRef]
- Chiang, C.; Bånkestad, D.; Hoch, G. Reaching natural growth: The significance of light and temperature fluctuations in plant performance in indoor growth facilities. Plants 2020, 9, 1312. [Google Scholar] [CrossRef]
- Clementson, L.A.; Wojtasiewicz, B. Dataset on the absorption characteristics of extracted phytoplankton pigments. Data Br. 2019, 24, 103875. [Google Scholar] [CrossRef]
- Wahid, A.; Rasul, E. Handbook of Photosynthesis; CRC Press: Boca Raton, FL, USA, 2005; ISBN 9781420027877. [Google Scholar]
- Valladares, F.; Niinemets, Ü. Shade tolerance, a key plant feature of complex nature and consequences. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 237–257. [Google Scholar] [CrossRef] [Green Version]
- Gommers, C.M.M.; Visser, E.J.W.; Onge, K.R.S.; Voesenek, L.A.C.J.; Pierik, R. Shade tolerance: When growing tall is not an option. Trends Plant Sci. 2013, 18, 65–71. [Google Scholar] [CrossRef]
- Givnish, T.J. Adaptation to sun and shade: A whole-plant perspective. Aust. J. Plant Physiol. 1988, 139–143. [Google Scholar] [CrossRef] [Green Version]
- Sessa, G.; Carabelli, M.; Possenti, M.; Morelli, G.; Ruberti, I. Multiple pathways in the control of the shade avoidance response. Plants 2018, 7, 102. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.A.; Kuwar, G.; Clarke, J.L.; Blystad, D.R.; Gislerød, H.R.; Olsen, J.E.; Torre, S. Artificial light from light emitting diodes (LEDs) with a high portion of blue light results in shorter poinsettias compared to high pressure sodium (HPS) lamps. Sci. Hortic. 2012, 147, 136–143. [Google Scholar] [CrossRef]
- Pinho, P.; Jokinen, K.; Halonen, L. Horticultural lighting—Present and future challenges. Light. Res. Technol. 2012, 44, 427–437. [Google Scholar] [CrossRef]
- Shirley, H.L. The Influence of Light Intensity and Light Quality Upon the Growth of Plants. Am. J. Bot. 1929, 16, 354–390. [Google Scholar] [CrossRef]
- Horton, J.L.; Neufeld, H.S. Photosynthetic responses of Microstegium vimineum (Trin.) A. Camus, a shade-tolerant, C4 grass, to variable light environments. Oecologia 1998, 114, 11–19. [Google Scholar] [CrossRef]
- Cipollini, D. Interactive effects of lateral shading and jasmonic acid on morphology, phenology, seed production, and defense traits in Arabidopsis thaliana. Int. J. Plant Sci. 2005, 166, 955–959. [Google Scholar] [CrossRef]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef]
- Keller, M.M.; Jaillais, Y.; Pedmale, U.V.; Moreno, J.E.; Chory, J.; Ballaré, C.L. Cryptochrome 1 and phytochrome B control shade-avoidance responses in Arabidopsis via partially independent hormonal cascades. Plant J. 2011, 67, 195–207. [Google Scholar] [CrossRef]
- Sellaro, R.; Crepy, M.; Trupkin, S.A.; Karayekov, E.; Buchovsky, A.S.; Rossi, C.; Casal, J.J. Cryptochrome as a sensor of the blue/green ratio of natural radiation in Arabidopsis. Plant Physiol. 2010, 154, 401–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, H.; Whitelam, G.C. The shade avoidance syndrome: Multiple responses mediated by multiple phytochromes. Plant Cell Environ. 1997, 20, 840–844. [Google Scholar] [CrossRef] [Green Version]
- Brouwer, B.; Gardeström, P.; Keech, O. In response to partial plant shading, the lack of phytochrome A does not directly induce leaf senescence but alters the fine-tuning of chlorophyll biosynthesis. J. Exp. Bot. 2014, 65, 4037–4049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichtenthaler, H.K.; Ač, A.; Marek, M.V.; Kalina, J.; Urban, O. Differences in pigment composition, photosynthetic rates and chlorophyll fluorescence images of sun and shade leaves of four tree species. Plant Physiol. Biochem. 2007, 45, 577–588. [Google Scholar] [CrossRef]
- Owens, D.K.; Alerding, A.B.; Crosby, K.C.; Bandara, A.B.; Westwood, J.H.; Winkel, B.S.J. Functional analysis of a predicted flavonol synthase gene family in arabidopsis. Plant Physiol. 2008, 147, 1046–1061. [Google Scholar] [CrossRef] [Green Version]
- Zeng, X.Q.; Chow, W.S.; Su, L.J.; Peng, X.X.; Peng, C.L. Protective effect of supplemental anthocyanins on Arabidopsis leaves under high light. Physiol. Plant. 2010, 138, 215–225. [Google Scholar] [CrossRef]
- Bailey, S.; Horton, P.; Walters, R.G. Acclimation of Arabidopsis thaliana to the light environment: The relationship between photosynthetic function and chloroplast composition. Planta 2004, 218, 793–802. [Google Scholar] [CrossRef]
- Karageorgou, P.; Manetas, Y. The importance of being red when young: Anthocyanins and the protection of young leaves of Quercus coccifera from insect herbivory and excess light. Tree Physiol. 2006, 26, 613–621. [Google Scholar] [CrossRef] [Green Version]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [Green Version]
- Murchie, E.H.; Horton, P. Toward C4 rice: Learning from the acclimation of photosynthesis in the C3 leaf. Charting New Pathw. C4 Rice 2008, 333–350. [Google Scholar] [CrossRef]
- Boyes, D.C.; Zayed, A.M.; Ascenzi, R.; McCaskill, A.J.; Hoffman, N.E.; Davis, K.R.; Görlach, J. Growth stage-based phenotypic analysis of Arabidopsis: A model for high throughput functional genomics in plants. Plant Cell 2001, 13, 1499–1510. [Google Scholar] [CrossRef] [Green Version]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Kölling, K.; George, G.M.; Künzli, R.; Flütsch, P.; Zeeman, S.C. A whole-plant chamber system for parallel gas exchange measurements of Arabidopsis and other herbaceous species. Plant Methods 2015, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage | Description |
|---|---|
| Germination | Plants with fully expanded cotyledons |
| 4-leaf stage | Plants with the first two rosette leaves bigger than the cotyledons |
| 6-leaf stage | Plants with the second couple of rosette leaves bigger than the first one |
| Bolting and flowering | Plants with a floral stalk taller than 1 cm |
| Silique formation | Plants with at least one fully developed silique |
| Ripening and senescence | Plants with at least one silique totally brown or open with ripe seeds |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beatrice, P.; Terzaghi, M.; Chiatante, D.; Scippa, G.S.; Montagnoli, A. Morpho-Physiological Responses of Arabidopsis thaliana L. to the LED-Sourced CoeLux® System. Plants 2021, 10, 1310. https://doi.org/10.3390/plants10071310
Beatrice P, Terzaghi M, Chiatante D, Scippa GS, Montagnoli A. Morpho-Physiological Responses of Arabidopsis thaliana L. to the LED-Sourced CoeLux® System. Plants. 2021; 10(7):1310. https://doi.org/10.3390/plants10071310
Chicago/Turabian StyleBeatrice, Peter, Mattia Terzaghi, Donato Chiatante, Gabriella Stefania Scippa, and Antonio Montagnoli. 2021. "Morpho-Physiological Responses of Arabidopsis thaliana L. to the LED-Sourced CoeLux® System" Plants 10, no. 7: 1310. https://doi.org/10.3390/plants10071310
APA StyleBeatrice, P., Terzaghi, M., Chiatante, D., Scippa, G. S., & Montagnoli, A. (2021). Morpho-Physiological Responses of Arabidopsis thaliana L. to the LED-Sourced CoeLux® System. Plants, 10(7), 1310. https://doi.org/10.3390/plants10071310