
Modifying Carbohydrate Supply to Fruit during Development Changes the Composition and Flavour of Actinidia chinensis var. chinensis ‘Zesy002’ Kiwifruit

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
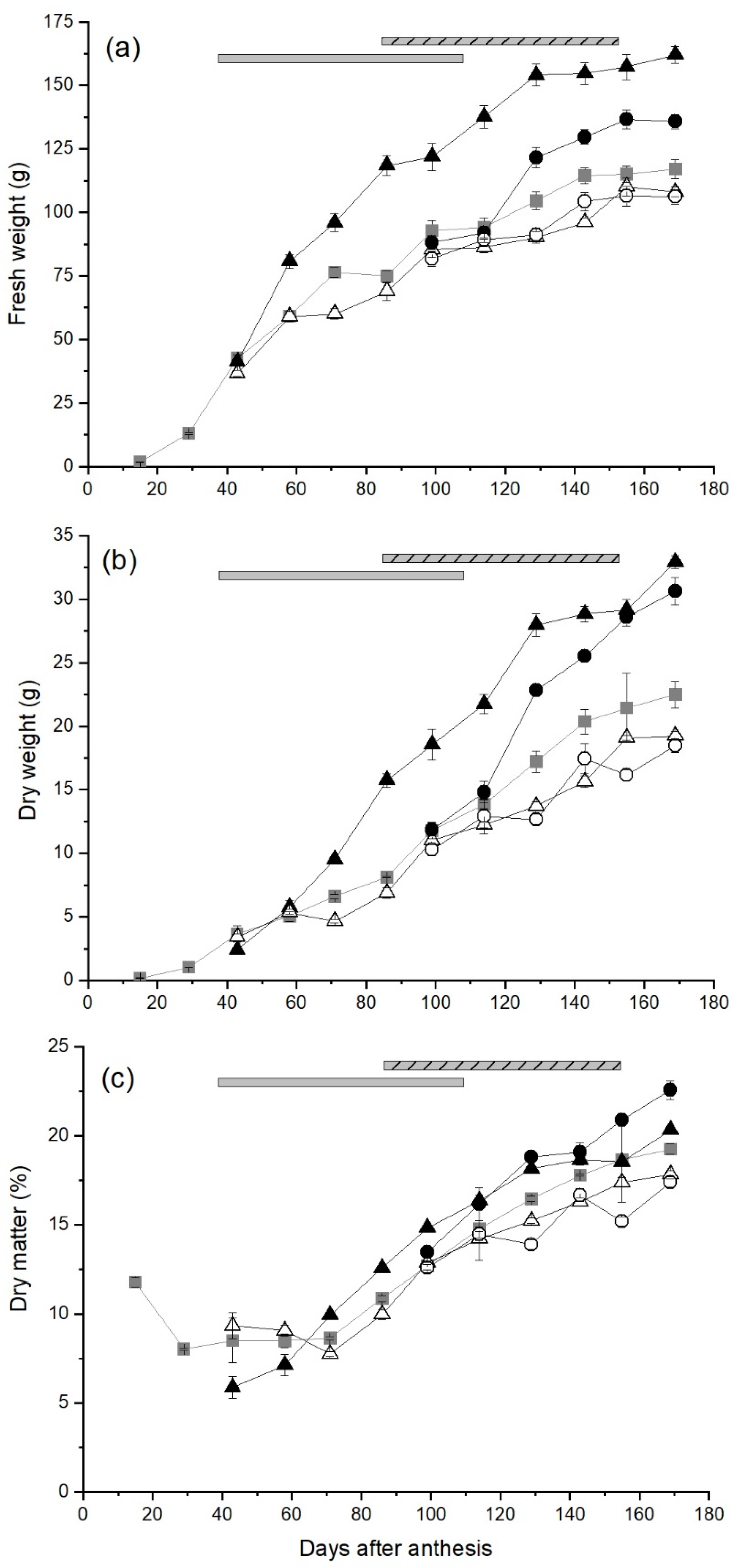
2.1. The Effect of Carbohydrate Supply on Fruit Growth
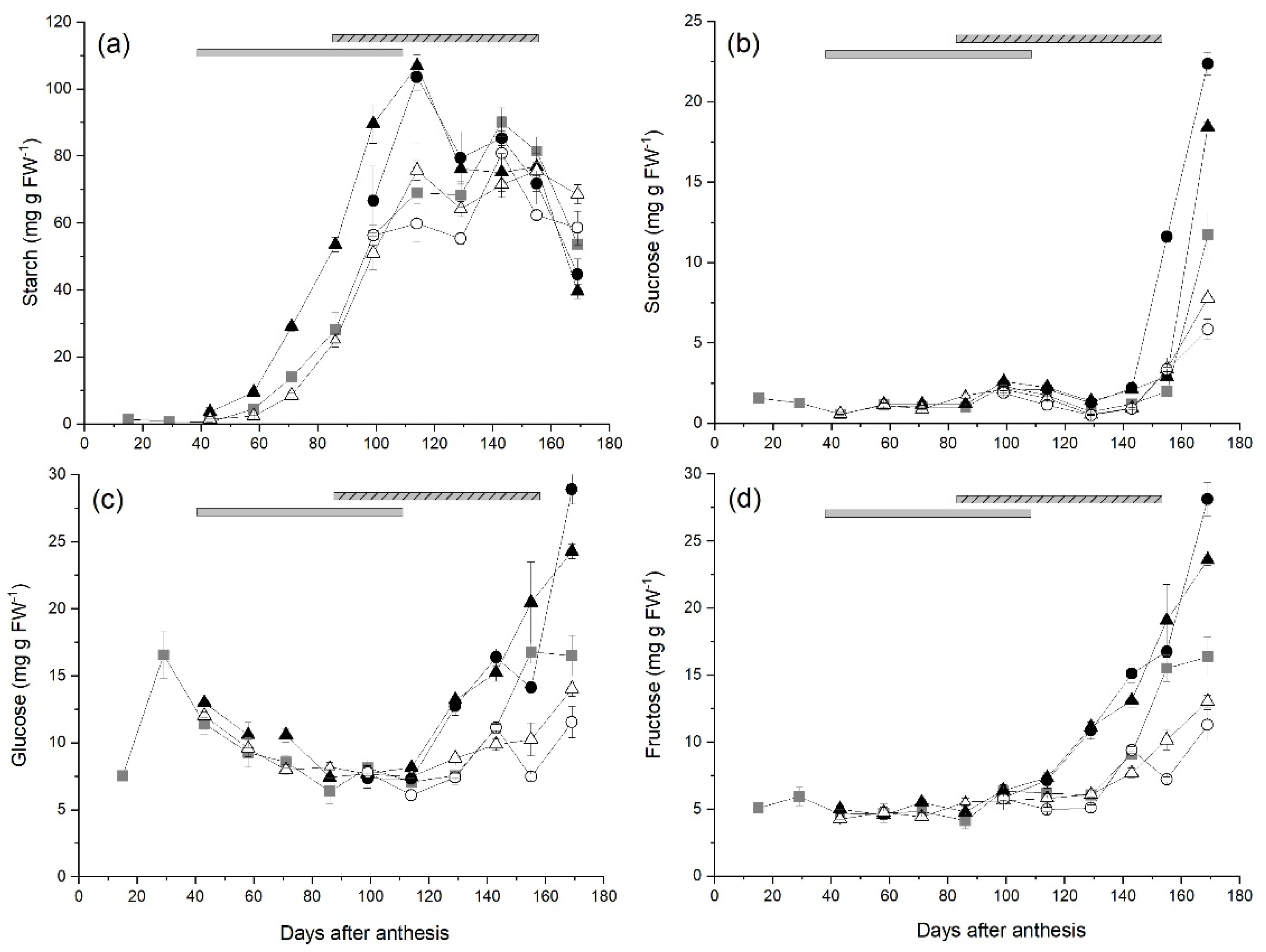
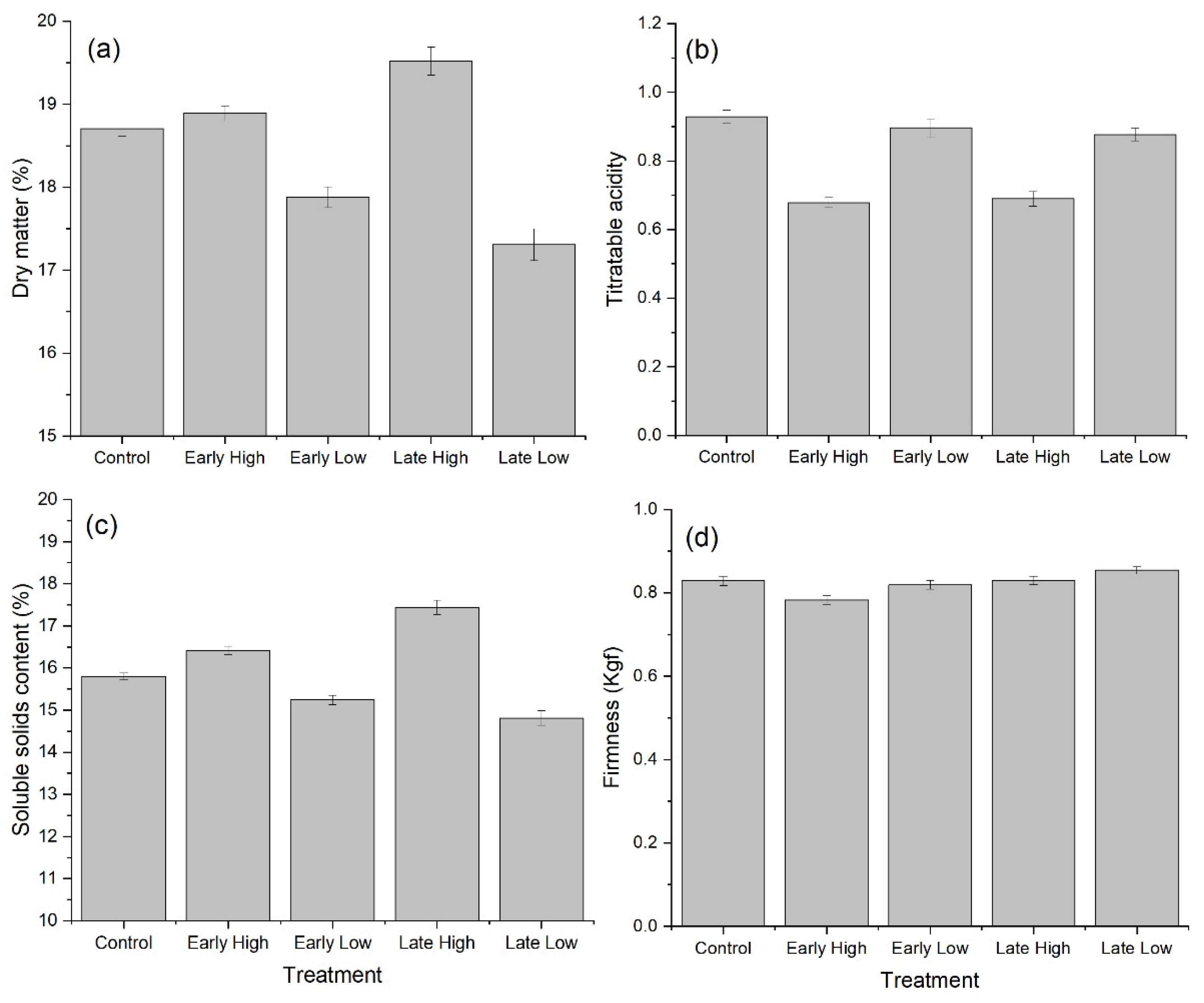
2.2. Starch and Sugar Concentrations in Fruit during Development
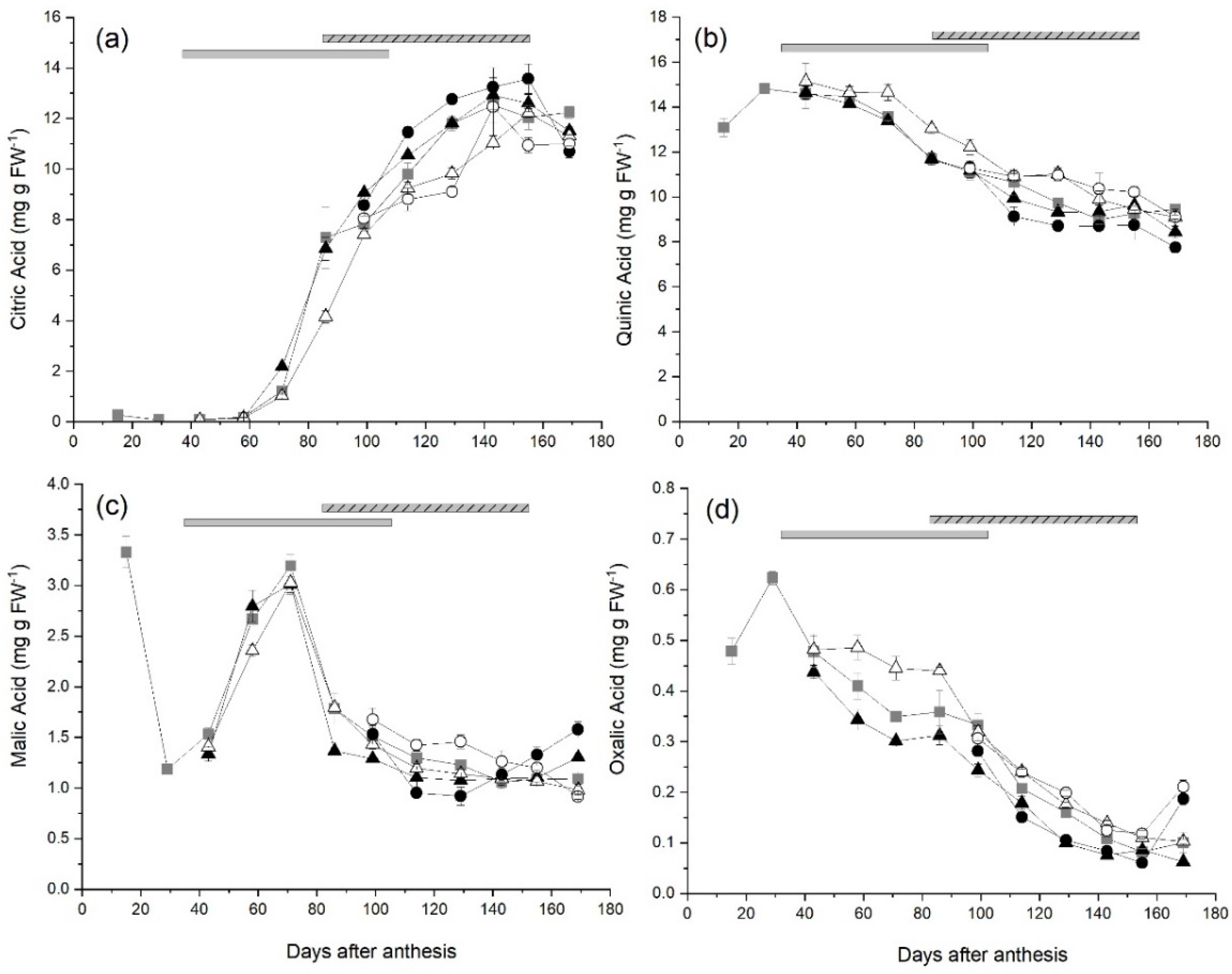
2.3. Changes in Organic Acid Concentrations in Fruit
2.4. Cytokinin Concentrations during Fruit Development
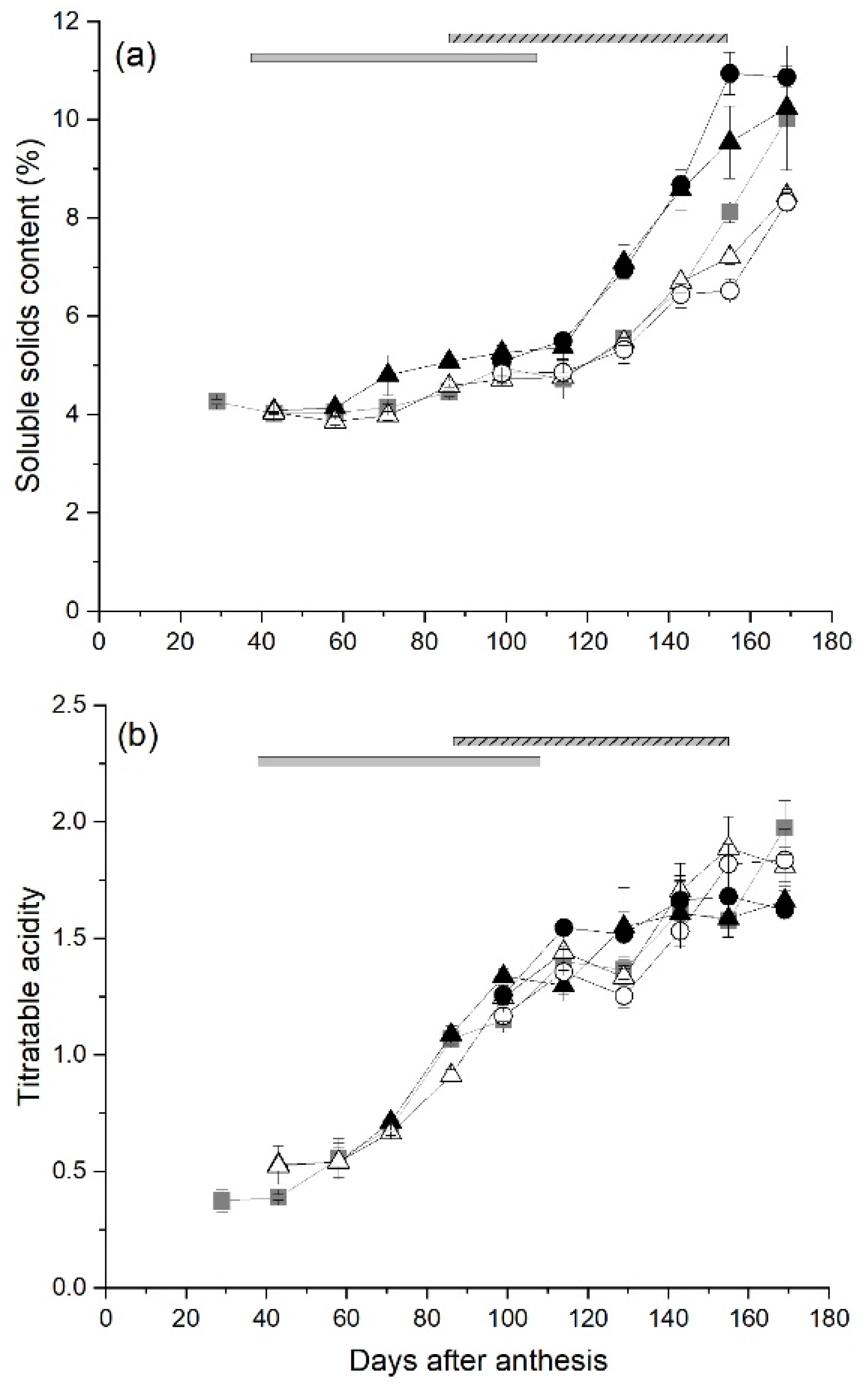
2.5. Maturation of Fruit
2.6. Sensory Evaluation of Fruit
3. Discussion
3.1. The Effect of Carbohydrate Source Manipulation on Fruit Growth
3.2. Fruit Metabolite Accumulation Is Affected by Carbohydrate Supply to Fruit
3.3. Effects of Carbohydrate Supply on Fruit Maturation
4. Materials and Methods
4.1. Plant Material
4.2. Experimental Design
4.3. Fruit Measurements
4.4. Soluble Carbohydrates, Starch, Organic Acid and Cytokinin Analyses
4.5. Fruit Maturity Assessments, Harvest and Fruit Ripening
4.6. Consumer Evaluation
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferguson, A.R. Kiwifruit in the World-2014. Acta Hortic. 2015, 1096, 33–46. [Google Scholar] [CrossRef]
- Richardson, A.; Boldingh, H.; Kashuba, P.; Knight, G.; Ellingham, D. Flowering time determines the weight and composition of Actinidia chinensis var. chinensis ‘Zesy002’ kiwifruit. Sci. Hortic. 2019, 246, 741–748. [Google Scholar] [CrossRef]
- Richardson, A.C.; McAneney, K.J. Influence of fruit number on fruit weight and yield of kiwifruit. Sci. Hortic. 1990, 42, 233–241. [Google Scholar] [CrossRef]
- Nardozza, S.; Boldingh, H.L.; Richardson, A.C.; Costa, G.; Marsh, H.; MacRae, E.A.; Clearwater, M.J. Variation in carbon content and size in developing fruit of Actinidia deliciosa genotypes. Funct. Plant Biol. 2010, 37, 545–554. [Google Scholar] [CrossRef]
- Howell, G.S. Sustainable grape productivity and the growth-yield relationship: A review. Am. J. Enol. Vitic. 2001, 52, 165–174. [Google Scholar]
- Guardiola, J.L.; Garcia-Luis, A. Thinning effects on citrus yield and fruit size. In Eight International Symposium on Plant Bioregulators in Fruit Production; Guardiola, J.L., Martinez, J.L.G., Eds.; International Society for Horticultural Science: Leuven, Belgium, 1997; pp. 463–473. [Google Scholar]
- Palmer, J.; Diack, R.; Johnston, J.; Boldingh, H. Manipulation of fruit dry matter accumulation and fruit size in ‘Scifresh’ apple through alteration of the carbon supply, and its relationship with apoplastic sugar composition. J. Hortic. Sci. Biotechnol. 2013, 88, 483–489. [Google Scholar] [CrossRef]
- Seal, A.G.; Clark, C.J.; Sharrock, K.R.; de Silva, H.N.; Jaksons, P.; Wood, M.E. Choice of pollen donor affects weight but not composition of Actinidia chinensis var. chinensis ‘Zesy002’ (Gold3) kiwifruit. N. Z. J. Crop Hortic. Sci. 2018, 46, 133–143. [Google Scholar] [CrossRef]
- Harker, F.R.; Carr, B.T.; Lenjo, M.; MacRae, E.A.; Wismer, W.V.; Marsh, K.B.; Williams, M.; White, A.; Lund, C.M.; Walker, S.B.; et al. Consumer liking for kiwifruit flavour: A meta-analysis of five studies on fruit quality. Food Qual. Prefer. 2009, 20, 30–41. [Google Scholar] [CrossRef]
- Jaeger, S.R.; Rossiter, K.L.; Wismer, W.V.; Harker, F.R. Consumer-driven product development in the kiwifruit industry. Food Qual. Prefer. 2003, 14, 187–198. [Google Scholar] [CrossRef]
- Crisosto, C.H.; Crisosto, G.M. Understanding consumer acceptance of early harvested ‘Hayward’ kiwifruit. Postharvest Biol. Technol. 2001, 22, 205–213. [Google Scholar] [CrossRef] [Green Version]
- Marsh, K.B.; Friel, E.N.; Gunson, A.; Lund, C.; MacRae, E. Perception of flavour in standardised fruit pulps with additions of acids or sugars. Food Qual. Prefer. 2006, 17, 376–386. [Google Scholar] [CrossRef]
- Richardson, A.C.; McAneney, K.J.; Dawson, T.E. Carbohydrate dynamics in kiwifruit. J. Hortic. Sci. 1997, 72, 907–917. [Google Scholar] [CrossRef]
- Jordan, R.B.; Seelye, R.J. Relationship between taste perception, density and soluble solids concentration in kiwifruit (Actinidia deliciosa). N. Z. J. Crop Hortic. Sci. 2009, 37, 303–317. [Google Scholar] [CrossRef]
- Stec, M.G.H.; Hodgson, J.A.; Macrae, E.A.; Triggs, C.M. Role of fruit firmness in the sensory evaluation of kiwifruit (Actinidia deliciosa cv. Hayward). J. Sci. Food Agric. 1989, 47, 417–433. [Google Scholar] [CrossRef]
- Gamble, J.; Harker, F.R.; Jaeger, S.R.; White, A.; Bava, C.; Beresford, M.; Stubbings, B.; Wohlers, M.; Hofman, P.J.; Marques, R.; et al. The impact of dry matter, ripeness and internal defects on consumer perceptions of avocado quality and intentions to purchase. Postharvest Biol. Technol. 2010, 57, 35–43. [Google Scholar] [CrossRef]
- McPherson, H.G.; Richardson, A.C.; Snelgar, W.P.; Patterson, K.J.; Currie, M.B. Flower quality and fruit size in kiwifruit (Actinidia deliciosa). N. Z. J. Crop Hortic. Sci. 2001, 29, 93–101. [Google Scholar] [CrossRef] [Green Version]
- Richardson, A.C.; Boldingh, H.L.; McAtee, P.A.; Gunaseelan, K.; Luo, Z.W.; Atkinson, R.G.; David, K.M.; Burdon, J.N.; Schaffer, R.J. Fruit development of the diploid kiwifruit, Actinidia chinensis ‘Hort16A’. BMC Plant Biol. 2011, 11. [Google Scholar] [CrossRef] [Green Version]
- Hopping, M.E. Structure and development of fruit and seeds in chinese gooseberry (Actinidia chinensis Planch.). N. Z. J. Bot. 1976, 14, 63–68. [Google Scholar] [CrossRef]
- Minchin, P.E.H.; Richardson, A.C.; Patterson, K.J.; Martin, P.J. Prediction of final weight for Actinidia chinensis ‘Hort16A’ fruit. N. Z. J. Crop Hortic. Sci. 2003, 31, 147–157. [Google Scholar] [CrossRef]
- Nardozza, S.; Boldingh, H.L.; Osorio, S.; Hohne, M.; Wohlers, M.; Gleave, A.P.; MacRae, E.A.; Richardson, A.C.; Atkinson, R.G.; Sulpice, R.; et al. Metabolic analysis of kiwifruit (Actinidia deliciosa) berries from extreme genotypes reveals hallmarks for fruit starch metabolism. J. Exp. Bot. 2013, 64, 5049–5063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boldingh, H.; Smith, G.S.; Klages, K.U. Seasonal concentrations of non-structural carbohydrates of five Actinidia species in fruit, leaf and fine root tissue. Ann. Bot. 2000, 85, 469–476. [Google Scholar] [CrossRef] [Green Version]
- Richardson, A.C.; Marsh, K.B.; Macrae, E.A. Temperature effects on satsuma mandarin fruit development. J. Hortic. Sci. 1997, 72, 919–929. [Google Scholar] [CrossRef]
- Walton, E.F.; De Jong, T.M. Growth and compositional changes in kiwifruit berries from three Californian locations. Ann. Bot. 1990, 66. [Google Scholar] [CrossRef]
- Marsh, K.B.; Harker, F.R. Acid metabolism in kiwifruit. In the Kiwifruit Genome; Testolin, R., Huang, H.-W., Ferguson, A.R., Eds.; Springer International Publishing: Basel, Switzerland, 2016; pp. 179–188. [Google Scholar]
- Montefiori, M.; McGhie, T.K.; Hallett, I.C.; Costa, G. Changes in pigments and plastid ultrastructure during ripening of green-fleshed and yellow-fleshed kiwifruit. Sci. Hortic. 2009, 119, 377–387. [Google Scholar] [CrossRef]
- Pilkington, S.M.; Montefiori, M.; Jameson, P.E.; Allan, A.C. The control of chlorophyll levels in maturing kiwifruit. Planta 2012, 236, 1615–1628. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.H.; Seal, A.G.; MacRae, E.A.; Wang, M.Y. Identifying volatile compounds associated with sensory and fruit attributes in diploid Actinidia chinensis (kiwifruit) using multivariate analysis. Euphytica 2011, 181, 179–195. [Google Scholar] [CrossRef]
- Snelgar, W.P.; Manson, P.J.; Martin, P.J. Influence of time of shading on flowering and yield of kiwifruit vines. J. Hortic. Sci. 1992, 67, 481–487. [Google Scholar] [CrossRef]
- Hassall, A.K.; Pringle, G.J.; MacRae, E.A. Development, maturation, and postharvest responses of Actinidia arguta (Sieb. et Zucc.) Planch. ex Miq. fruit. N. Z. J. Crop Hortic. Sci. 1998, 26, 95–108. [Google Scholar] [CrossRef]
- Lai, R.; Woolley, D.J.; Lawes, G.S. Effect of leaf to fruit ratio on fruit-growth of kiwifruit (Actinidia deliciosa). Sci. Hortic. 1989, 39, 247–255. [Google Scholar] [CrossRef]
- Snelgar, W.P.; Minchin, P.E.H.; Blatmann, P.; Hall, A.J. Sink priority on ‘Hayward’ kiwifruit vines. N. Z. J. Crop Hortic. Sci. 2012, 40, 253–263. [Google Scholar] [CrossRef]
- Minchin, P.E.H.; Snelgar, W.P.; Blattmann, P.; Hall, A.J. Competition between fruit and vegetative growth in Hayward kiwifruit. N. Z. J. Crop Hortic. Sci. 2010, 38, 101–112. [Google Scholar] [CrossRef]
- Patterson, K.J.; Mason, K.A.; Gould, K.S. Effects of CPPU (N-(2-chloro-4-pyridyl)-N′-phenylurea) on fruit growth, maturity, and storage quality of kiwifruit. N. Z. J. Crop Hortic. Sci. 1993, 21, 253–261. [Google Scholar] [CrossRef] [Green Version]
- Antognozzi, E.; Battistelli, A.; Famiani, F.; Moscatello, S.; Stanica, F.; Tombesi, A. Influence of CPPU on carbohydrate accumulation and metabolism in fruits of Actinidia deliciosa (A Chev.). Sci. Hortic. 1996, 65, 37–47. [Google Scholar] [CrossRef]
- Boyd, L.M.; Barnett, A.M.; Johnstone, P.R.; Searle, B.P.; Catto, W.D.; Pentreath, R. Nitrogen fertiliser affects fresh weight and dry matter concentration in ‘Hayward’ kiwifruit. Acta Hortic. 2013, 984, 197–202. [Google Scholar] [CrossRef]
- Tombesi, A.; Antognozzi, E.; Palliotti, A. Influence of light exposure on characteristics and storage life of kiwifruit. N. Z. J. Crop Hortic. Sci. 1993, 21, 85–90. [Google Scholar] [CrossRef]
- Richardson, A.C.; Marsh, K.B.; Boldingh, H.L.; Pickering, A.H.; Bulley, S.M.; Frearson, N.J.; Ferguson, A.R.; Thornber, S.E.; Bolitho, K.M.; Macrae, E.A. High growing temperatures reduce fruit carbohydrate and vitamin C in kiwifruit. Plant Cell Environ. 2004, 27, 423–435. [Google Scholar] [CrossRef]
- Antognozzi, E.; Tombesi, A.; Ferranti, F.; Frenguelli, G. Influence of sink competition on peduncle histogenesis in kiwifruit. N. Z. J. Crop Hortic. Sci. 1991, 19, 433–439. [Google Scholar] [CrossRef]
- Nardozza, S.; Boldingh, H.L.; Wohlers, M.W.; Gleave, A.P.; Luo, Z.W.; Costa, G.; MacRae, E.A.; Clearwater, M.J.; Richardson, A.C. Exogenous cytokinin application to Actinidia chinensis var. deliciosa ‘Hayward’ fruit promotes fruit expansion through water uptake. Hortic. Res. 2017, 4. [Google Scholar] [CrossRef] [PubMed]
- Nardozza, S.; Cooney, J.; Boldingh, H.L.; Hewitt, K.G.; Trower, T.; Jones, D.; Thrimawithana, A.H.; Allan, A.C.; Richardson, A.C. Phytohormone and transcriptomic analysis reveals endogenous cytokinins affect kiwifruit growth under restricted carbon supply. Metabolites 2020, 10, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, J.W.; Giuliani, R.; Adams, H.M. Effect of crop load on fruiting and leaf photosynthesis of ‘Braeburn’/M.26 apple trees. Tree Physiol. 1997, 17, 741–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siham, M.; Bussi, C.; Lescourret, F.; Genard, M.; Habib, R.; Gilreath, J. Pruning intensity and fruit load influence on vegetative and fruit growth in ‘Alexandra’ peach. In Proceedings of the 118th Annual Meeting of the Florida State Horticultural Society; Florida State Horticultural Society: Alexandria, VA, USA, 2005; pp. 266–269. [Google Scholar]
- Usenik, V.; Orazem, P.; Stampar, F. Low leaf to fruit ratio delays fruit maturity of ‘Lapins’ sweet cherry on Gisela 5. Sci. Hortic. 2010, 126, 33–36. [Google Scholar] [CrossRef]
- Boyd, L.M.; Barnett, A.M. Manipulation of whole-vine carbon allocation using girdling, pruning, and fruit thinning affects fruit numbers and quality in kiwifruit. Hortscience 2011, 46, 590–595. [Google Scholar] [CrossRef] [Green Version]
- Burdon, J.; Lallu, N.; Pidakala, P.; Barnett, A.M. Is the 6.2 degrees brix soluble solids harvest index suitable for ‘Hayward’ kiwifruit from high productivity orchard management systems? Acta Hortic. 2011, 913, 539–546. [Google Scholar] [CrossRef]
- Burdon, J.; Pidakala, P.; Martin, P.; Billing, D. Softening of ‘Hayward’ kiwifruit on the vine and in storage: The effects of temperature. Sci. Hortic. 2017, 220, 176–182. [Google Scholar] [CrossRef]
- Montefiori, M.; Espley, R.V.; Stevenson, D.D.; Cooney, J.M.; Datson, P.M.; Saiz, A.; Atkinson, R.G.; McGhie, T.; Hellens, R.P.; Allan, A.C. The control of kiwifruit red flesh colour. Acta Hortic. 2011, 913, 103–109. [Google Scholar] [CrossRef]
- Nardozza, S.; Boldingh, H.L.; Kashuba, M.P.; Feil, R.; Jones, D.; Thrimawithana, A.H.; Ireland, H.S.; Philippe, M.; Wohlers, M.W.; McGhie, T.K.; et al. Carbon starvation reduces carbohydrate and anthocyanin accumulation in red-fleshed fruit via trehalose 6-phosphate and MYB27. Plant Cell Environ. 2020, 43, 819–835. [Google Scholar] [CrossRef] [PubMed]
- Judd, M.J.; McAneney, K.J.; Wilson, K.S. Influence of water-stress on kiwifruit growth. Irrig. Sci. 1989, 10, 303–311. [Google Scholar] [CrossRef]
- Alspach, P.A.; Pyke, N.B.; Morgan, C.G.T.; Ruth, J.E. Influence of application rates of bee-collected pollen on the fruit size of kiwifruit. N. Z. J. Crop Hortic. Sci. 1991, 19, 19–24. [Google Scholar] [CrossRef]
- Burge, G.K.; Spence, C.B.; Marshall, R.R. Kiwifruit—Effects of thinning on fruit size, vegetative growth, and return bloom. N. Z. J. Exp. Agric. 1987, 15, 317–324. [Google Scholar] [CrossRef]
- Boldingh, H.; Richardson, A.; Marsh, K.B. The flavour seesaw: Balancing sugars and acids. N. Z. Kiwifruit J. 2008, 185, 42–44. [Google Scholar]
- Lewis, D.H.; Burge, G.K.; Schmierer, D.M.; Jameson, P.E. Cytokinins and fruit development in the kiwifruit (Actinidia deliciosa). 1. Changes during fruit development. Physiol. Plant. 1996, 98, 179–186. [Google Scholar] [CrossRef]
- Smith, G.S.; Clark, C.J.; Boldingh, H.L. Seasonal accumulation of starch by components of the kiwifruit vine. Ann. Bot. 1992, 70, 19–25. [Google Scholar] [CrossRef]
- Nardozza, S.; Kashuba, P.; McCaughan, L.; Philippe, M.; Wohlers, M.; Montefiori, M.; Currie, M.; Richardson, A. Leaves are important to obtain consistent red flesh pigmentation in Actinidia chinensis fruit. Sci. Hortic. 2015, 197, 496–503. [Google Scholar] [CrossRef]
- Marsh, K.B.; Boldingh, H.L.; Shilton, R.S.; Laing, W.A. Changes in quinic acid metabolism during fruit development in three kiwifruit species. Funct. Plant Biol. 2009, 36, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.Q.; Huang, Q.; Jia, D.F.; Zhong, M.; Tao, J.J.; Liao, G.L.; Huang, C.H.; Xu, X.B. Characterization of organic acid metabolism and expression of related genes during fruit development of Actinidia eriantha ‘Ganmi 6’. Plants 2020, 9, 332. [Google Scholar] [CrossRef] [Green Version]
- Hunter, D.C.; Beresford, M.; White, A.; Jaeger, S.R.; Wohlers, M.; Richards, K.; Chheang, S.L.; Jin, D.; Fullerton, C.G.; Clark, C.J. Measuring perception of acidity and sweetness in kiwifruit. Acta Hortic. 2018, 1218, 191–198. [Google Scholar] [CrossRef]
- Burdon, J.; Pidakala, P.; Martin, P.; McAtee, P.A.; Boldingh, H.L.; Hall, A.; Schaffer, R.J. Postharvest performance of the yellow-fleshed ‘Hort16A’ kiwifruit in relation to fruit maturation. Postharvest Biol. Technol. 2014, 92, 98–106. [Google Scholar] [CrossRef]
- Pilkington, S.M.; Montefiori, M.; Galer, A.L.; Emery, R.J.N.; Allan, A.C.; Jameson, P.E. Endogenous cytokinin in developing kiwifruit is implicated in maintaining fruit flesh chlorophyll levels. Ann. Bot. 2013, 112, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Montefiori, M.; McGhie, T.K.; Costa, G.; Ferguson, A.R. Pigments in the fruit of red-fleshed kiwifruit (Actinidia chinensis and Actinidia deliciosa). J. Agric. Food Chem. 2005, 53, 9526–9530. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Descriptors | Control | Early High | Early Low | Late High | Late Low |
|---|---|---|---|---|---|
| Bland | 18.7a | 13.3a | 13.3a | 10.7a | 17.3a |
| Crunchy | 4a | 4a | 5.3a | 4a | 2.7a |
| Fresh | 36a | 29.3a | 36a | 36a | 36a |
| Grassy/green | 2.7a | 4a | 6.7a | 5.3a | 8a |
| Juicy | 49.3a | 54.7a | 49.3a | 60a | 50.7a |
| Lemon/lime | 20a | 9.3a | 22.7a | 20a | 24a |
| Melting/smooth | 44a | 44a | 37.3a | 48a | 41.3a |
| Metallic | 2.7a | 2.7a | 2.7a | 2.7a | 4a |
| Mushy | 20a | 34.7a | 24a | 25.3a | 20a |
| Off-flavour | 1.3a | 4a | 2.7a | 4a | 10.7a |
| Over-ripe | 13.3a | 21.3a | 13.3a | 10.7a | 10.7a |
| Soft | 69.3a | 73.3a | 62.7a | 72a | 61.3a |
| Sour/acidic | 38.7ab | 20.0c | 44.0a | 26.7bc | 44.0a |
| Sweet | 57.3a | 66.7a | 41.3b | 72.0a | 40.0b |
| Tropical | 21.3a | 18.7a | 18.7a | 20a | 20a |
| Under-ripe | 6.7ab | 1.3b | 4.0ab | 2.7a | 12.0a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lievre, D.L.; Anderson, R.; Boldingh, H.; Cooney, J.; Seelye, R.; Gould, N.; Hunter, D.; Jensen, D.; Pereira, T.; Wohlers, M.; et al. Modifying Carbohydrate Supply to Fruit during Development Changes the Composition and Flavour of Actinidia chinensis var. chinensis ‘Zesy002’ Kiwifruit. Plants 2021, 10, 1328. https://doi.org/10.3390/plants10071328
Lievre DL, Anderson R, Boldingh H, Cooney J, Seelye R, Gould N, Hunter D, Jensen D, Pereira T, Wohlers M, et al. Modifying Carbohydrate Supply to Fruit during Development Changes the Composition and Flavour of Actinidia chinensis var. chinensis ‘Zesy002’ Kiwifruit. Plants. 2021; 10(7):1328. https://doi.org/10.3390/plants10071328
Chicago/Turabian StyleLievre, Danielle Le, Rachelle Anderson, Helen Boldingh, Janine Cooney, Richard Seelye, Nick Gould, Denise Hunter, Dwayne Jensen, Trisha Pereira, Mark Wohlers, and et al. 2021. "Modifying Carbohydrate Supply to Fruit during Development Changes the Composition and Flavour of Actinidia chinensis var. chinensis ‘Zesy002’ Kiwifruit" Plants 10, no. 7: 1328. https://doi.org/10.3390/plants10071328
APA StyleLievre, D. L., Anderson, R., Boldingh, H., Cooney, J., Seelye, R., Gould, N., Hunter, D., Jensen, D., Pereira, T., Wohlers, M., Clearwater, M., & Richardson, A. (2021). Modifying Carbohydrate Supply to Fruit during Development Changes the Composition and Flavour of Actinidia chinensis var. chinensis ‘Zesy002’ Kiwifruit. Plants, 10(7), 1328. https://doi.org/10.3390/plants10071328