
The Tripartite Rhizobacteria-AM Fungal-Host Plant Relationship in Winter Wheat: Impact of Multi-Species Inoculation, Tillage Regime and Naturally Occurring Rhizobacteria Species

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Sites
2.2. Rhizobacteria and AM Fungi Inoculum
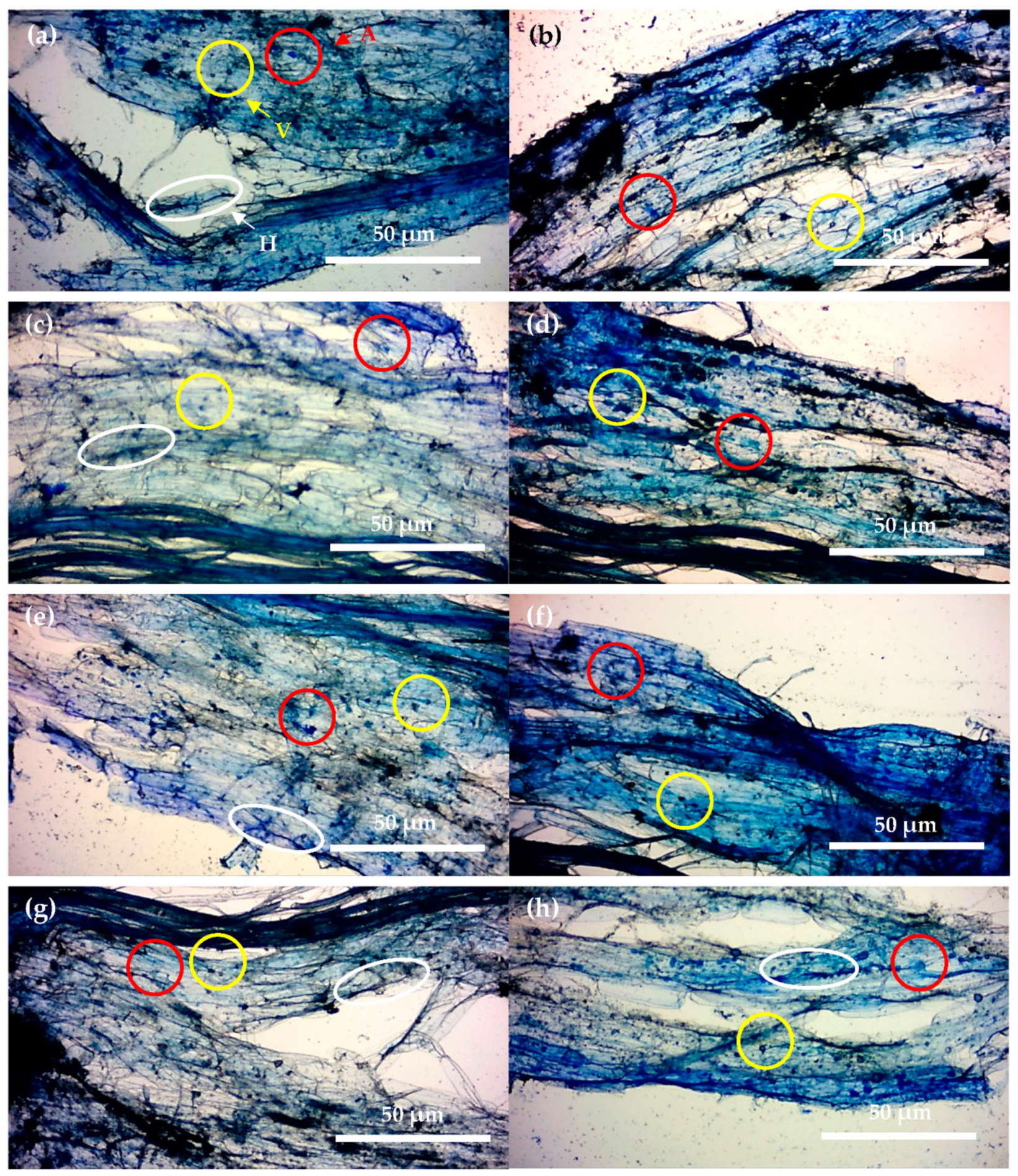
2.3. Impact of Rhizobacteria on AM Fungi Root Arbuscule Number and Winter Wheat Growth
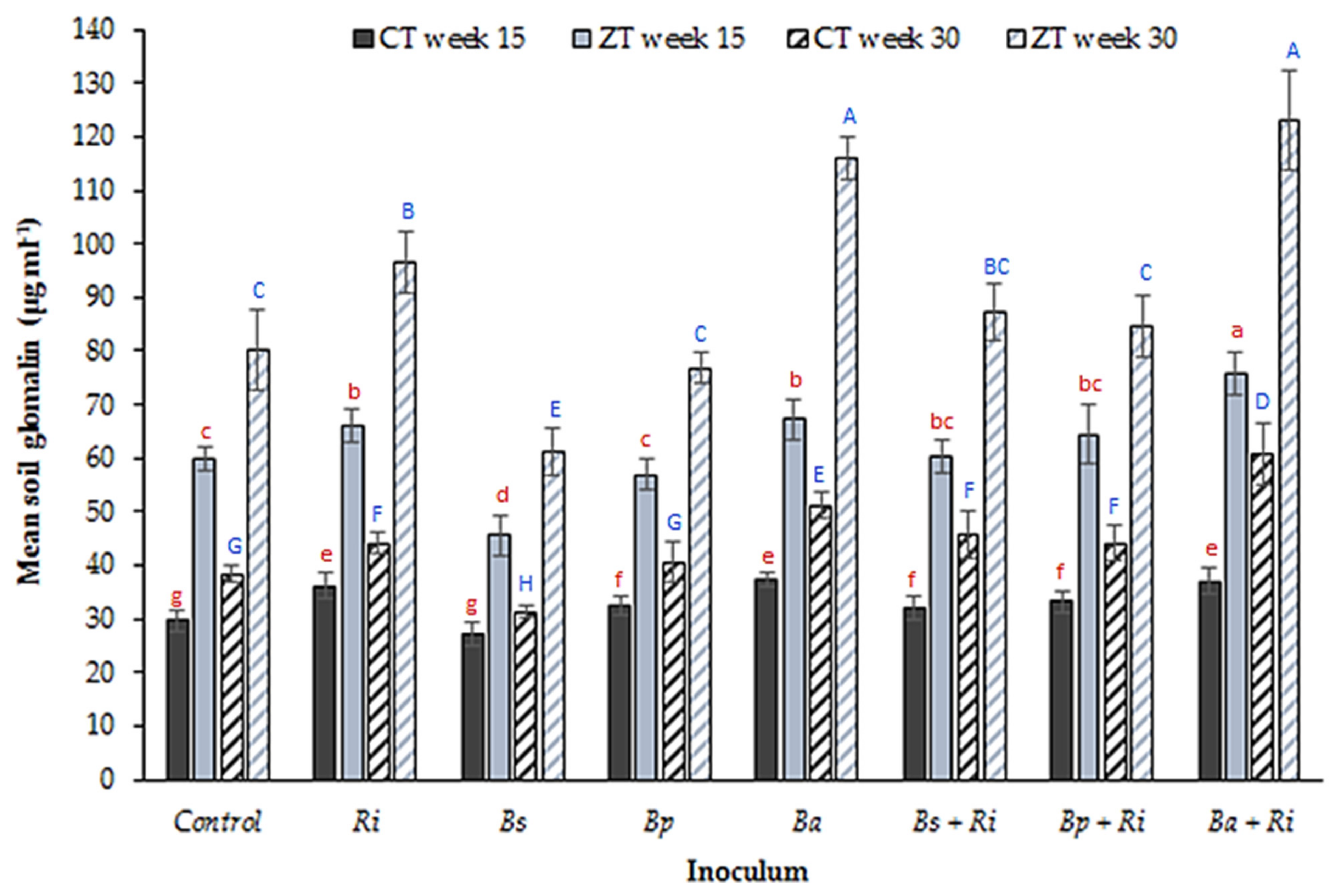
2.4. Impact of Rhizobacteria on Soil Glomalin
2.5. Soil Nutrient Testing
2.6. Molecular Quantification of AM Fungi and Rhizobacteria Species with qPCR
2.7. Statistical Analysis
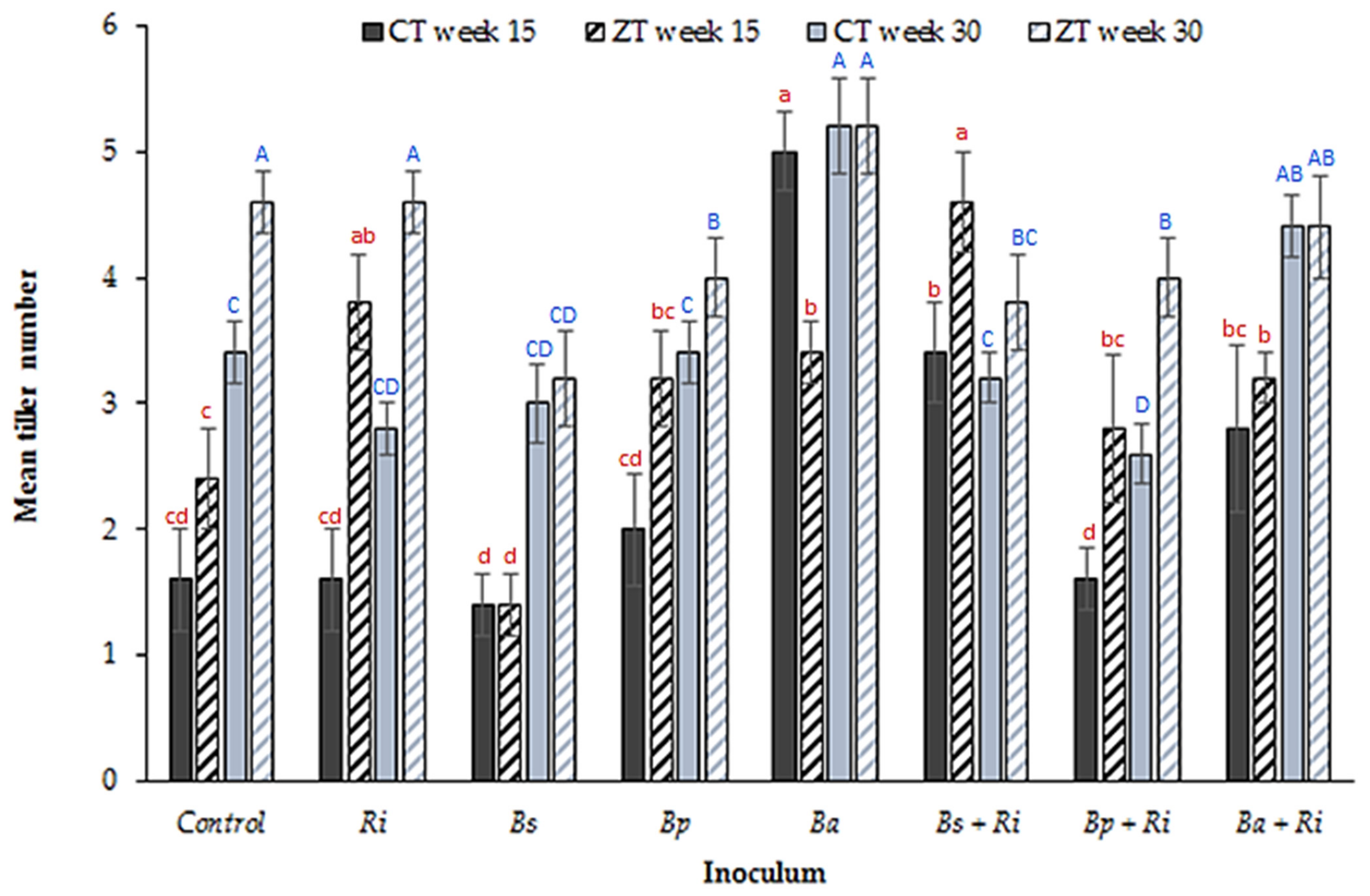
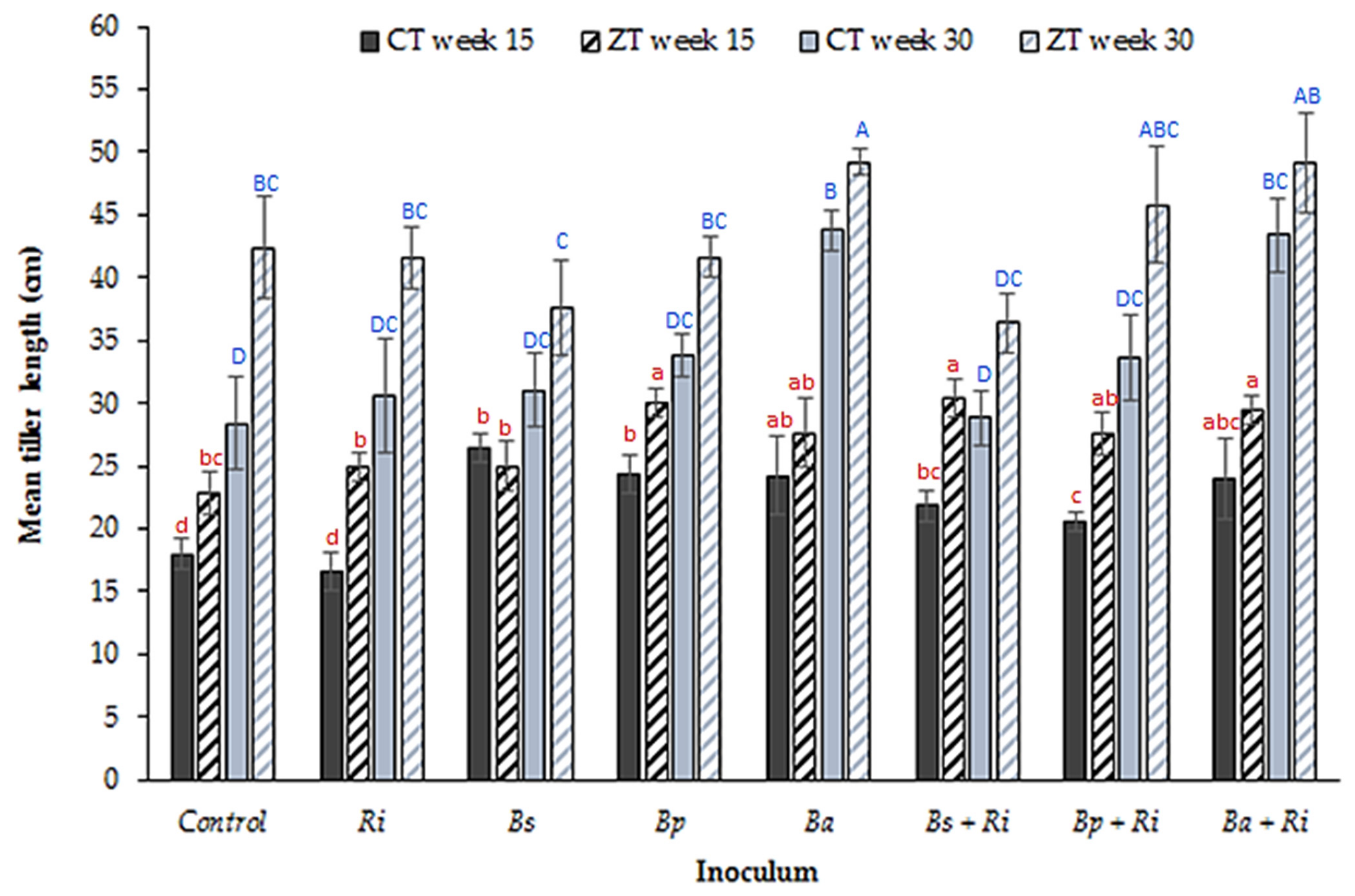
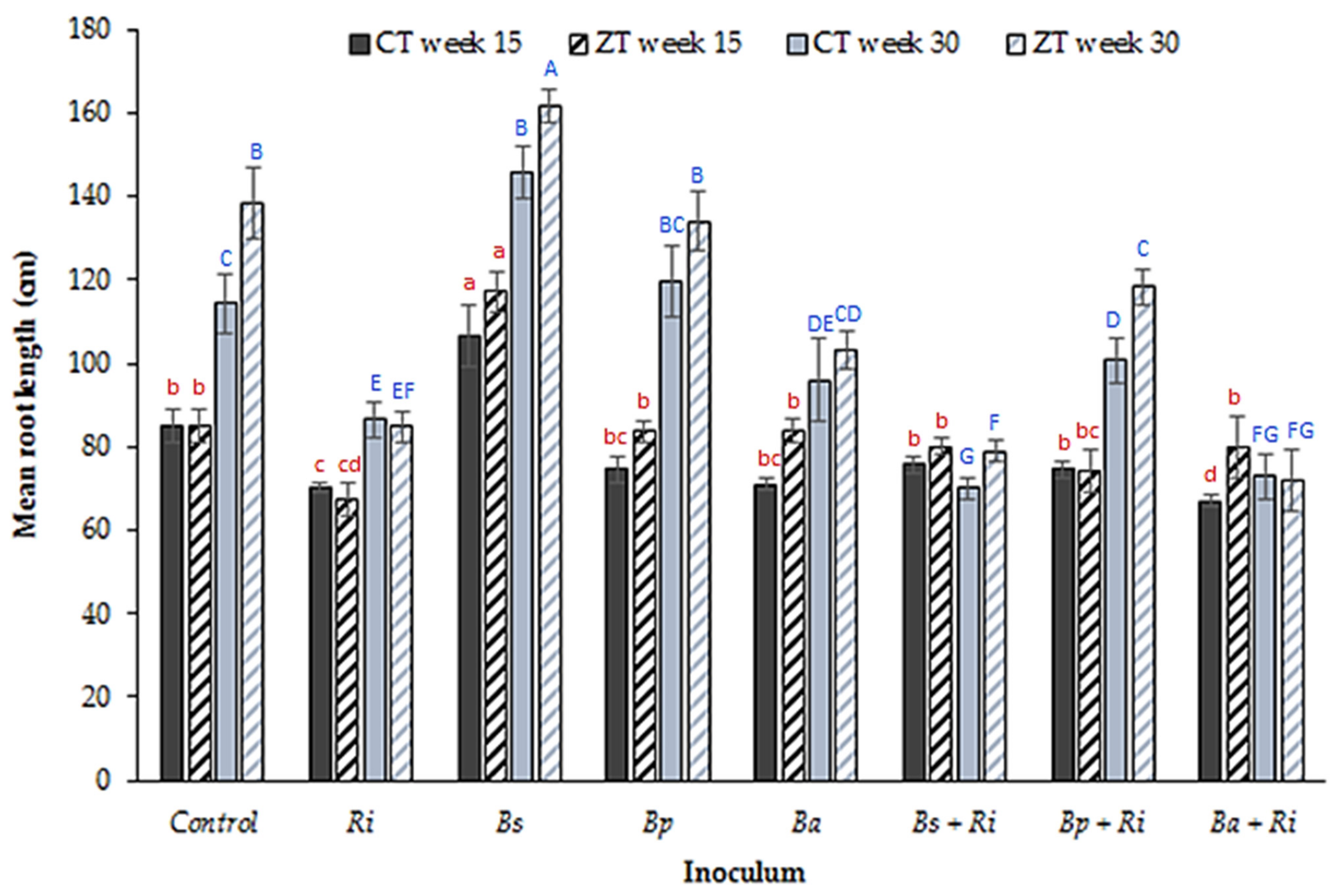
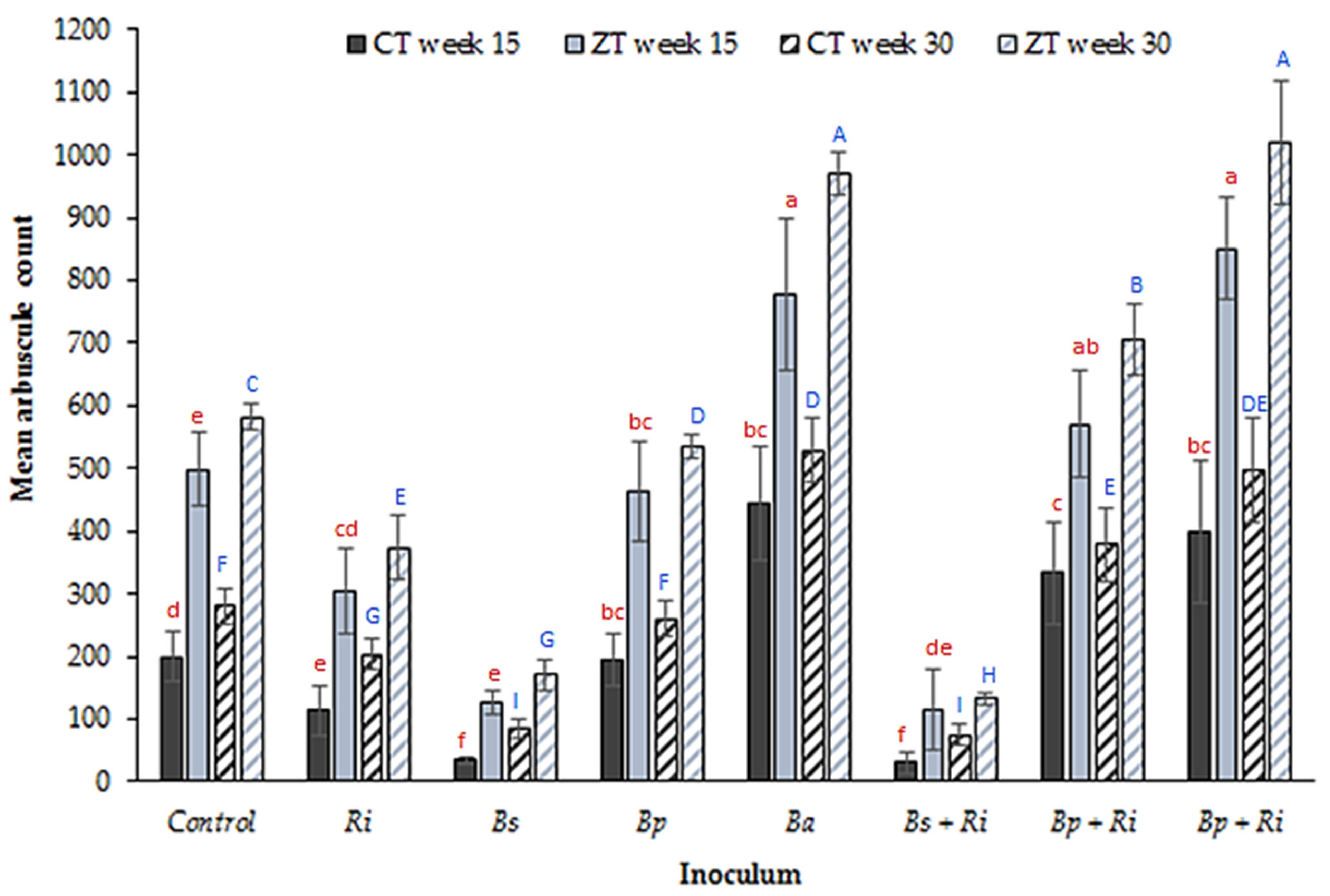
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kottke, I.; Nebel, M. The evolution of mycorrhiza-like associations in liverworts: An update. New Phytol. 2005, 167, 330–334. [Google Scholar] [CrossRef] [PubMed]
- Wilkes, T.I.; Warner, D.J.; Edmonds-Brown, V.; Davies, K.G.; Denholm, I. Zero Tillage Systems Conserve Arbuscular Mycorrhizal Fungi, Enhancing Soil Glomalin and Water Stable Aggregates with Implications for Soil Stability. Soil Syst. 2021, 5, 4. [Google Scholar] [CrossRef]
- Frew, A.; Powell, J.R.; Hiltpold, I.; Allsopp, P.G.; Sallam, N.; Johnson, S.N. Host plant colonisation by arbuscular mycorrhizal fungi stimulates immune function whereas high root silicon concentrations diminish growth in a soil-dwelling herbivore. Soil Biol. Biochem. 2017, 112, 117–126. [Google Scholar] [CrossRef]
- Begum, N.; Qin, C.; Ahanger, M.A.; Raza, S.; Khan, M.I.; Ashraf, M.; Ahmed, N.; Zhang, L. Role of Arbuscular Mycorrhizal Fungi in Plant Growth Regulation: Implications in Abiotic Stress Tolerance. Front. Plant Sci. 2019, 10, 1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berruti, A.; Lumini, E.; Balestrini, R.; Bianciotto, V. Arbuscular Mycorrhizal Fungi as Natural Biofertilizers: Let’s Benefit from Past Successes. Front. Microbiol. 2016, 6, 1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selosse, M.; Strullu-Derrien, C.; Martin, F.M.; Kamoun, S.; Kenrick, P. Plants, fungi and oomycetes: A 400-million years affair that shapes the biosphere. New Phytol. 2015, 206, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.; Read, D. Mycorrhiza Symbiosis, 3rd ed.; Academic Press: San Diego, CA, USA, 2008. [Google Scholar]
- Wright, S.F.; Frankee-Snyder, M.; Morton, J.B. Time-course study and partial characterization of a protein on hyphae of arbuscular mycorrhizal fungi during active colonization of roots. Plant Soil. 1996, 181, 193–203. [Google Scholar] [CrossRef]
- Raklami, A.; Bechtaoui, N.; Tahiri, A.; Anil, M.; Meddich, A.; Oufdou, K. Use of Rhizobacteria and Mycorrhizae Consortium in the Open Field as a Strategy for Improving Crop Nutrition, Productivity and Soil Fertility. Front. Microbiol. 2019, 10, 1106. [Google Scholar] [CrossRef] [Green Version]
- Labbe, J.; Weston, D.; Dunkirk, N.; Pelletier, D.; Tuskan, G. Newly identified helper bacteria stimulate ectomycorrhizal formation in Populus. Front. Plant Sci. 2014, 10, 579. [Google Scholar] [CrossRef]
- Nanjundappa, A.; Bagyaraj, D.J.; Saxena, A.K.; Kumar, M.; Chakdar, H. Interaction between arbuscular mycorrhizal fungi and Bacillus spp. in soil enhancing growth of crop plants. Fungal. Biol. Biotechnol. 2019, 6, 23. [Google Scholar] [CrossRef] [Green Version]
- Munkvold, L.; Kjøller, R.; Vestberg, M.; Rosendahl, S.; Jakobsen, I. High functional diversity within species of arbuscular mycorrhizal fungi. New Phytol. 2004, 164, 357–364. [Google Scholar] [CrossRef]
- Nuccio, E.E.; Hodge, A.; Pett-Ridge, J.; Herman, D.J.; Weber, P.K.; Firestone, M.K. An arbuscular mycorrhizal fungus significantly modifies the soil bacterial community and nitrogen cycling during litter decomposition. Environ. Microbiol. 2013, 15, 1870–1881. [Google Scholar] [CrossRef]
- Svenningsen, N.B.; Watts-Williams, S.J.; Joner, E.J.; Battini, F.; Efthymiou, A.; Cruz-Paredes, C.; Nybroe, O.; Jakobsen, I. Suppression of the activity of arbuscular mycorrhizal fungi by the soil microbiota. ISME J. 2018, 12, 1296–1307. [Google Scholar] [CrossRef] [Green Version]
- Borriss, R. Use of plant-associated Bacillus strains as biofertilizers and biocontrol agents in agriculture. In Bacteria in Agrobiology: Plant Growth Responses; Springer: Berlin/Heidelberg, Germany, 2011; pp. 41–76. [Google Scholar]
- Xu, S.J.; Kim, B.S. Biocontrol of fusarium crown and root rot and promotion of growth of tomato by paenibacillus strains isolated from soil. Mycobiology 2014, 42, 158–166. [Google Scholar] [CrossRef] [Green Version]
- Wilkes, T.I.; Warner, D.J.; Edmonds-Brown, V.; Davies, K.G. Species-Specific Interactions of Bacillus Innocula and Arbuscular Mycorrhizal Fungi Symbiosis with Winter Wheat. Microorganisms 2020, 8, 1795. [Google Scholar] [CrossRef]
- Alam, M.; Khaliq, A.; Sattar, A.; Shukla, R.S.; Anwar, M.; Dharni, S. Synergistic effect of arbuscular mycorrhizal fungi and Bacillus subtilis on the biomass and essential oil yield of rose-scented geranium (Pelargonium graveolens). Arch. Agron. Soil Sci. 2011, 57, 889–898. [Google Scholar] [CrossRef]
- Pérez-De-Luque, A.; Tille, S.; Johnson, I.; Pascual-Pardo, D.; Ton, J.; Cameron, D.D. The interactive effects of arbuscular mycorrhiza and plant growth-promoting rhizobacteria synergistically enhance host plant defences against pathogens. Sci. Rep. 2019, 7, 16409. [Google Scholar] [CrossRef]
- Kabir, Z. Tillage or no-tillage: Impact on mycorrhizae. Can. J. Plant Sci. 2005, 85, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Brito, I.; Goss, M.J.; de Carvalho, M.; Chatagnier, O.; van Tuinen, D. Impact of tillage system on arbuscular mycorrhiza fungal communities in the soil under Mediterranean conditions. Soil Tillage Res. 2012, 121, 63–67. [Google Scholar] [CrossRef]
- Wilkes, T.; Warner, D.J.; Edmonds-Brown, V.; Davies, K.G.; Denholm, I. A comparison of methodologies for the staining and quantification of intracellular components of arbuscular mycorrhizal fungi in the root cortex of two varieties of winter wheat. Access Microbiol. 2020, 2, e000083. [Google Scholar] [CrossRef]
- Alguacil, M.M.; Lumini, E.; Roldán, A.; Salinas-García, J.R.; Bonfante, P.; Bianciotto, V. The impact of tillage practices on arbuscular mycorrhizal fungal diversity in subtropical crops. Ecol. Appl. 2008, 18, 527–536. [Google Scholar] [CrossRef]
- Driver, J.D.; Holben, W.E.; Rillig, M.C. Characterization of glomalin as a hyphal wall component of arbuscular mycorrhizal fungi. Soil Biol. Biochem. 2005, 37, 101–106. [Google Scholar] [CrossRef]
- Lovelock, C.E.; Wright, S.F.; Clark, D.A.; Ruess, R.W. Soil stocks of glomalin produced by arbuscular mycorrhizal fungi across a tropical rain forest landscape. J. Ecol. 2004, 92, 278–287. [Google Scholar] [CrossRef]
- Brown, K.; Wherrett, A. Measuring Soil Texture in the Laboratory, 2019. Available online: http://soilquality.org.au/factsheets/soil-texture-measuring-in-the-lab (accessed on 25 March 2018).
- AHDB (Agriculture and Horticulture Development Board). Nutrient Management Guide (RB209), Section 4 Arable Crops; AHDB: Stoneleigh, UK, 2020. [Google Scholar]
- Sun, X.-G.; Hu, W.; Tang, M.; Chen, H. Characterizing and handling different kinds of AM fungal spores in the rhizosphere. World J. Microbiol. Biotechnol. 2016, 32, 97. [Google Scholar] [CrossRef]
- Lancashire, P.D.; Bleiholder, H.; Van Den Boom, T.; Langelüddeke, P.; Stauss, R.; Weber, E.; Witzenberger, A. A uniform decimal code for growth stages of crops and weeds. Ann. Appl. Biol. 1991, 119, 561–601. [Google Scholar] [CrossRef]
- Wright, S.; Upadhyaya, A. A survey of soils for aggregate stability and glomalin, a glycoprotein produced by hyphae of arbuscular mycorrhizal fungi. Plant Soil 1996, 198, 97–107. [Google Scholar] [CrossRef]
- Agnolucci, M.; Palla, M.; Cristani, C.; Cavallo, N.; Giovannetti, M.; De Angelis, M.; Minervini, F. Beneficial plant microorganisms affect the endophytic bacterial communities of durum wheat roots as detected by different molecular approaches. Front. Microbiol. 2019, 10, 2500. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Lu, X.; Liao, Y. Effect of Tillage Treatment on the Diversity of Soil Arbuscular Mycorrhizal Fungal and Soil Aggregate-Associated Carbon Content. Front. Microbiol. 2018, 9, 2986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhury, S.P.; Hartmann, A.; Gao, X.; Borriss, R. Biocontrol mechanism by root-associated Bacillus amyloliquefaciens FZB42—A review. Front. Microbiol. 2015, 6, 780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkes, T.I.; Warner, D.J.; Davies, K.G.; Edmonds-Brown, V.R. Tillage, glyphosate and beneficial Arbuscular Mycorrhizal fungi: Optimizing crop management for plant-fungal symbiosis. Agriculture 2020, 10, 520. [Google Scholar] [CrossRef]
- Palme, K.; Nagy, F. A new gene for auxin synthesis. Cell 2008, 133, 31–32. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.J.; Fang, X.; Li, C.Y.; Yang, L.; Chen, X.Y. General and specialized tyrosine metabolism pathways in plants. aBIOTECH 2019, 1, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Helander, M.; Saloniemi, I.; Omacini, M.; Druille, M.; Salminen, J.P.; Saikkonen, K. Glyphosate decreases mycorrhizal colonization and affects plant-soil feedback. Sci. Total Environ. 2018, 642, 285–291. [Google Scholar] [CrossRef]
- Helander, M.; Saloniemi, I.; Saikkonen, K. Trends in Northern ecosystems. Trends Plant Sci. 2012, 17, 569–574. [Google Scholar] [CrossRef]
- Escobar Ortega, J.S.; Aguilar Vásquez, N.N.; Ávila Alba, T.; García de Salamone, I.E. Impact of management of cover crop–soybean agroecosystems on rhizosphere microbial communities. Eur. J. Soil Sci. 2020, 72, 1154–1176. [Google Scholar] [CrossRef]
- Zablotowicz, R.M.; Reddy, K.N. Impact of glyphosate on the Bradyrhizobium japonicum symbiosis with glyphosate-resistant transgenic soybean: A minireview. J. Environ. Qual. 2004, 33, 825–831. [Google Scholar] [CrossRef]
- Lewis, K.A.; Tzilivakis, J.; Warner, D.J.; Green, A. An international database for pesticide risk assessments and management. Hum. Ecol. Risk Assess. Int. J. 2004, 22, 1050–1064. [Google Scholar] [CrossRef] [Green Version]
- Partoazar, M.; Hoodaji, M.; Tahmourespour, A. The effect of glyphosate application on soil microbial activities in agricultural land. Afr. J. Biotechnol. 2011, 10, 19419–19424. [Google Scholar]
- Martinez, D.A.; Loening, U.E.; Graham, M.C. Impacts of glyphosate-based herbicides on disease resistance and health of crops: A review. Environ. Sci. Eur. 2018, 30, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Newman, M.M.; Hoilett, N.; Lorenz, N.; Dick, R.P.; Liles, M.R.; Ramsier, C.; Kloepper, J.W. Glyphosate effects on soil rhizosphere-associated bacterial communities. Sci. Total Environ. 2016, 543, 155–160. [Google Scholar] [CrossRef] [Green Version]
- Wijekoon, N.; Yapa, N. Assessment of plant growth promoting rhizobacteria (PGPR) on potential biodegradation of glyphosate in contaminated soil and aquifers. Groundw. Sustain. Dev. 2018, 7, 465–469. [Google Scholar] [CrossRef]
- Fisher, P.J.; Petrini, O.; Scott, H.M.L. The distribution of some fungal and bacterial endophytes in maize (Zea mays L.). New Phytol. 1992, 122, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Medina, A.; Probanza, A.; Mañero, F.G.; Azcón, R. Interactions of arbuscular-mycorrhizal fungi and Bacillus strains and their effects on plant growth, microbial rhizosphere activity (thymidine and leucine incorporation) and fungal biomass (ergosterol and chitin). Appl. Soil Ecol. 2003, 22, 15–28. [Google Scholar] [CrossRef]
- Requena, N.; Jimenez, I.; Toro, M.; Barea, J.M. Interactions between plant-growth-promoting rhizobacteria (PGPR), arbuscular mycorrhizal fungi and Rhizobium spp. in the rhizosphere of Anthyllis cytisoides, a model legume for revegetation in mediterranean semi-arid ecosystems. New Phytol. 1997, 136, 667–677. [Google Scholar] [CrossRef]
- Bashan, Y.; Levanony, H. Current status of Azospirillum inoculation technology: Azospirillum as a challenge for agriculture. Can. J. Microbiol. 1990, 36, 591–608. [Google Scholar] [CrossRef] [Green Version]
- McGonigle, T.P.; Fitter, A.H. Ecological specificity of vesicular-arbuscular mycorrhizal associations. Mycol. Res. 1990, 94, 120–122. [Google Scholar] [CrossRef]
- Compant, S.; Duffy, B.; Nowak, J.; Clément, C.; Barka, E.A. Use of plant growth-promoting bacteria for biocontrol of plant diseases: Principles, mechanisms of action, and future prospects. Appl. Environ. Microbiol. 2005, 71, 4951–4959. [Google Scholar] [CrossRef] [Green Version]
- Welbaum, G.E.; Sturz, A.V.; Dong, Z.; Nowak, J. Managing soil microorganisms to improve productivity of agro-ecosystems. Crit. Rev. Plant Sci. 2004, 23, 175–193. [Google Scholar] [CrossRef]
- Bais, H.P.; Park, S.W.; Weir, T.L.; Callaway, R.M.; Vivanco, J.M. How plants communicate using the underground information superhighway. Trends Plant Sci. 2004, 9, 26–32. [Google Scholar] [CrossRef]
- Bacilio, M.; Hernandez, J.P.; Bashan, Y. Restoration of giant cardon cacti in barren desert soil amended with common compost and inoculated with Azospirillum brasilense. Biol. Fertil. Soils 2006, 43, 112–119. [Google Scholar] [CrossRef]
- Reva, O.N.; Dixelius, C.; Meijer, J.; Priest, F.G. Taxonomic characterization and plant colonizing abilities of some bacteria related to Bacillus amyloliquefaciens and Bacillus subtilis. FEMS Microbiol. Ecol. 2004, 48, 249–259. [Google Scholar] [CrossRef]
- Dunstan, W.A.; Malajczuk, N.; Dell, B. Effects of bacteria on mycorrhizal development and growth of container grown Eucalyptus diversicolor F. Muell. seedlings. Plant Soil 1998, 201, 241–249. [Google Scholar] [CrossRef]
- Andrade, G.; Mihara, K.L.; Linderman, R.G.; Bethlenfalvay, G.J. Soil aggregation status and rhizobacteria in the mycorrhizosphere. Plant Soil 1998, 202, 89–96. [Google Scholar] [CrossRef]
- Couillerot, O.; Prigent-Combaret, C.; Caballero-Mellado, J.; Moënne-Loccoz, Y. Pseudomonas fluorescens and closely-related fluorescent pseudomonads as biocontrol agents of soil-borne phytopathogens. Lett. Appl. Microbiol. 2009, 48, 505–512. [Google Scholar] [CrossRef]
- Loper, J.E.; Henkels, M.D. Utilization of heterologous siderophores enhances levels of iron available to Pseudomonas putida in the rhizosphere. Appl. Environ. Microbiol. 1999, 65, 5357–5363. [Google Scholar] [CrossRef] [Green Version]
- Antoun, H.; Prévost, D. Ecology of plant growth promoting rhizobacteria. In PGPR: Biocontrol and Biofertilization; Springer: Dordrecht, The Netherlands, 2005; pp. 1–38. [Google Scholar]
- Trivedi, P.; Pandey, A.; Palni LM, S. Bacterial inoculants for field applications under mountain ecosystem: Present initiatives and future prospects. In Bacteria in Agrobiology: Plant Probiotics; Springer: Berlin, Heidelberg, 2012; pp. 15–44. [Google Scholar]
- Pamp, S.J.; Tolker-Nielsen, T. Multiple roles of biosurfactants in structural biofilm development by Pseudomonas aeruginosa. J. Bacteriol. 2007, 189, 2531–2539. [Google Scholar] [CrossRef] [Green Version]
- Dewey, F.M.; Wong, Y.L.; Seery, R.; Hollins, T.W.; Gurr, S.J. Bacteria associated with Stagonospora (Septoria) nodorum increase pathogenicity of the fungus. New Phytol. 1999, 144, 489–497. [Google Scholar] [CrossRef]
- Jäderlund, L.; Arthurson, V.; Granhall, U.; Jansson, J.K. Specific interactions between arbuscular mycorrhizal fungi and plant growth-promoting bacteria: As revealed by different combinations. FEMS Microbiol. Lett. 2008, 287, 174–180. [Google Scholar] [CrossRef]
- Chen, X.H.; Koumoutsi, A.; Scholz, R.; Borriss, R. More than anticipated–production of antibiotics and other secondary metabolites by Bacillus amyloliquefaciens FZB42. J. Mol. Microbiol. Biotechnol. 2009, 16, 14–24. [Google Scholar] [CrossRef]
- Tahmatsidou, V.; O’Sullivan, J.; Cassells, A.C.; Voyiatzis, D.; Paroussi, G. Comparison of AMF and PGPR inoculants for the suppression of Verticillium wilt of strawberry (Fragaria× ananassa cv. Selva). Appl. Soil Ecol. 2006, 32, 316–324. [Google Scholar] [CrossRef]
- Gamalero, E.; Berta, G.; Massa, N.; Glick, B.R.; Lingua, G. Interactions between Pseudomonas putida UW4 and Gigaspora rosea BEG9 and their consequences for the growth of cucumber under salt-stress conditions. J. Appl. Microbiol. 2010, 108, 236–245. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tillage Treatment | Inoculant | Week 15 | Week 30 |
|---|---|---|---|
| Conventional | Control | 102.5 | 102.52 |
| B subtilis | 102.52 | 102.49 | |
| B pumilis | 102.5 | 102.52 | |
| B amyloliquefaciens | 101.5 | 102.52 | |
| AM fungi | 102.5 | 102.52 | |
| B subtilis + AM fungi | 102.52 | 102.52 | |
| B pumilis + AM fungi | 102.5 | 102.49 | |
| B amyloliquefaciens + AM fungi | 102.52 | 102.52 | |
| Zero | Control | 0 | 0 |
| B subtilis | 101.5 | 102.49 | |
| B pumilis | 0 | 102.5 | |
| B amyloliquefaciens | 0 | 101.5 | |
| AM fungi | 0 | 102.49 | |
| B subtilis + AM fungi | 0 | 102.52 | |
| B pumilis + AM fungi | 0 | 102.5 | |
| B amyloliquefaciens + AM fungi | 0 | 102.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wilkes, T.I.; Warner, D.J.; Edmonds-Brown, V.; Davies, K.G.; Denholm, I. The Tripartite Rhizobacteria-AM Fungal-Host Plant Relationship in Winter Wheat: Impact of Multi-Species Inoculation, Tillage Regime and Naturally Occurring Rhizobacteria Species. Plants 2021, 10, 1357. https://doi.org/10.3390/plants10071357
Wilkes TI, Warner DJ, Edmonds-Brown V, Davies KG, Denholm I. The Tripartite Rhizobacteria-AM Fungal-Host Plant Relationship in Winter Wheat: Impact of Multi-Species Inoculation, Tillage Regime and Naturally Occurring Rhizobacteria Species. Plants. 2021; 10(7):1357. https://doi.org/10.3390/plants10071357
Chicago/Turabian StyleWilkes, Thomas I., Douglas J. Warner, Veronica Edmonds-Brown, Keith G. Davies, and Ian Denholm. 2021. "The Tripartite Rhizobacteria-AM Fungal-Host Plant Relationship in Winter Wheat: Impact of Multi-Species Inoculation, Tillage Regime and Naturally Occurring Rhizobacteria Species" Plants 10, no. 7: 1357. https://doi.org/10.3390/plants10071357
APA StyleWilkes, T. I., Warner, D. J., Edmonds-Brown, V., Davies, K. G., & Denholm, I. (2021). The Tripartite Rhizobacteria-AM Fungal-Host Plant Relationship in Winter Wheat: Impact of Multi-Species Inoculation, Tillage Regime and Naturally Occurring Rhizobacteria Species. Plants, 10(7), 1357. https://doi.org/10.3390/plants10071357