
Antifungal Activity against Botryosphaeriaceae Fungi of the Hydro-Methanolic Extract of Silybum marianum Capitula Conjugated with Stevioside

 ,
,  ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Material, Reagents, and Fungal Isolates
2.2. Preparation of Plant Extracts
2.3. Physicochemical Characterization of S. marianum Extracts
2.4. Preparation of Bioactive Formulations
2.5. Antifungal Activity Assessment
2.5.1. In Vitro Tests of Mycelial Growth Inhibition
2.5.2. Greenhouse Bioassays on Grafted Plants
2.6. Statistical Analyses
3. Results
3.1. Vibrational Characterization
3.2. Gas Chromatography–Mass Spectrometry (GC-MS)
3.3. Antifungal Activity
3.3.1. In Vitro Growth Inhibition Tests
3.3.2. Greenhouse Bioassays
4. Discussion
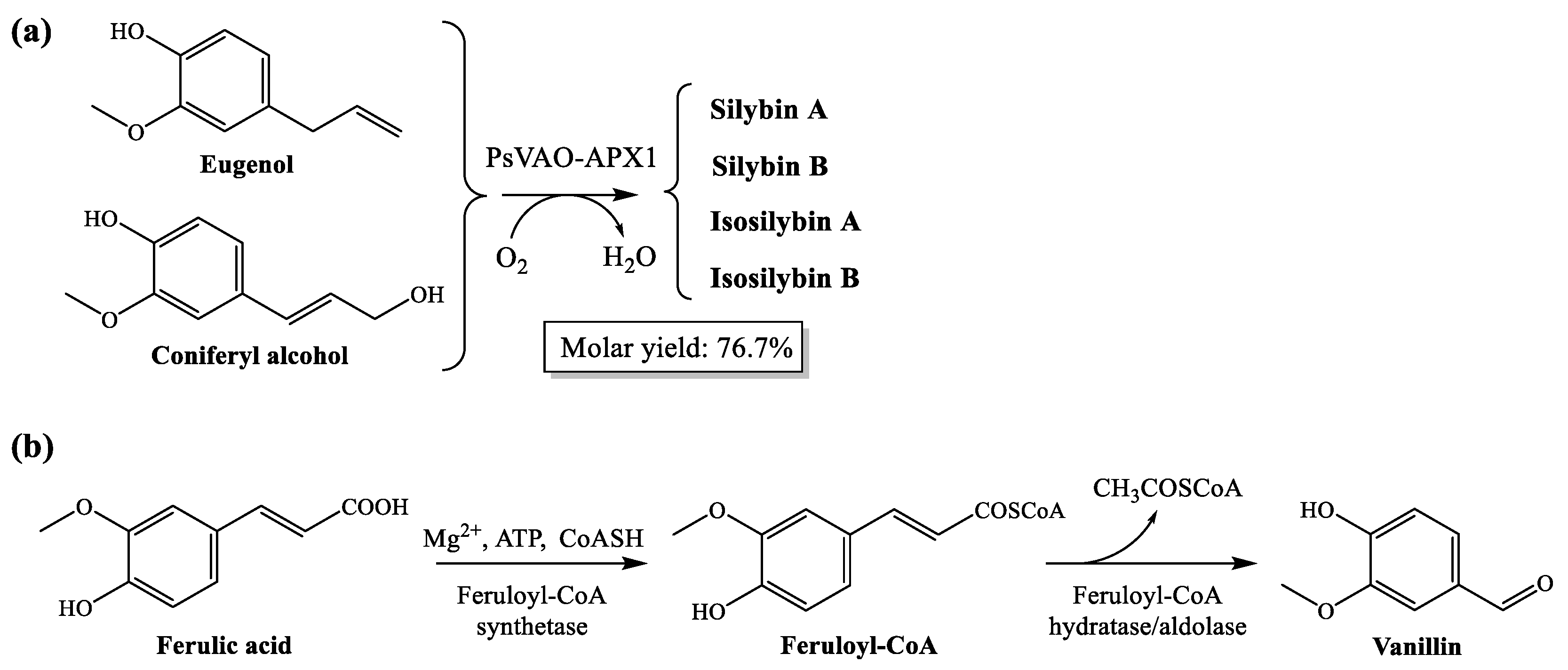
4.1. Valorization of Coniferyl Alcohol and Ferulic Acid
4.2. Efficacy of the Treatments
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Das, S.K.; Mukherjee, S.; Vasudevan, D. Medicinal properties of milk thistle with special reference to silymarin—An overview. Nat. Prod. Radiance 2008, 7, 182–192. [Google Scholar]
- Gazak, R.; Walterova, D.; Kren, V. Silybin and silymarin—New and emerging applications in medicine. Curr. Med. Chem. 2007, 14, 315–338. [Google Scholar] [CrossRef]
- Šimánek, V.; Kren, V.; Ulrichová, J.; Vicar, J.; Cvak, L. Silymarin: What is in the name…? An appeal for a change of editorial policy. Hepatology 2000, 32, 442–444. [Google Scholar] [CrossRef]
- ElSayed, A.I.; El-hamahmy, M.A.M.; Rafudeen, M.S.; Mohamed, A.H.; Omar, A.A. The impact of drought stress on antioxidant responses and accumulation of flavonolignans in milk thistle (Silybum marianum (L.) Gaertn). Plants 2019, 8, 611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AbouZid, S.; Ahmed, O.M. Silymarin flavonolignans: Structure–activity relationship and biosynthesis. In Studies in Natural Products Chemistry; Atta-ur-Rahman, F.R.S., Ed.; Elsevier: Amsterdam, The Netherlands, 2013; Volume 40, pp. 469–484. [Google Scholar]
- AbouZid, S.F.; Chen, S.-N.; McAlpine, J.B.; Friesen, J.B.; Pauli, G.F. Silybum marianum pericarp yields enhanced silymarin products. Fitoterapia 2016, 112, 136–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habán, M.; Otepka, P.; Kobida, L.; Habánová, M. Production and quality of milk thistle (Silybum marianum [L.] Gaertn.) cultivated in cultural conditions of warm agri-climatic macroregion. Hortic. Sci. 2009, 36, 69–74. [Google Scholar] [CrossRef] [Green Version]
- Lv, Y.; Gao, S.; Xu, S.; Du, G.; Zhou, J.; Chen, J. Spatial organization of silybin biosynthesis in milk thistle [Silybum marianum (L.) Gaertn]. Plant J. 2017, 92, 995–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drouet, S.; Tungmunnithum, D.; Lainé, É.; Hano, C. Gene expression analysis and metabolite profiling of silymarin biosynthesis during milk thistle (Silybum marianum (L.) Gaertn.) fruit ripening. Int. J. Mol. Sci. 2020, 21, 4730. [Google Scholar] [CrossRef] [PubMed]
- Saidi, I.; Bessam, H.M.; Benchaachoua, A. Effects of different extraction methods and solvents on the phenolic composition and antioxidant activity of Silybum marianum leaves extracts. Int. J. Med. Sci. Clin. Invent. 2018, 5, 3641–3647. [Google Scholar] [CrossRef]
- Mani, P.; Senthilkumar, S.; Sasikala, P.; Azhagumadhavan, S.; Jayaseelan, T.; Ganesan, S.; Padma, M. Phytochemical screening and GC-MS analysis of bioactive compounds present in ethanolic leaves extract of Silybum marianum (L). J. Drug Deliv. Ther. 2019, 9, 85–89. [Google Scholar] [CrossRef] [Green Version]
- Sulas, L.; Re, G.A.; Bullitta, S.; Piluzza, G. Chemical and productive properties of two Sardinian milk thistle (Silybum marianum (L.) Gaertn.) populations as sources of nutrients and antioxidants. Genet. Resour. Crop. Evol. 2015, 63, 315–326. [Google Scholar] [CrossRef]
- Vanholme, R.; Demedts, B.; Morreel, K.; Ralph, J.; Boerjan, W. Lignin biosynthesis and structure. Plant Physiol. 2010, 153, 895–905. [Google Scholar] [CrossRef] [Green Version]
- Daurade-Le Vagueresse, M.H.; Romiti, C.; Grosclaude, C.; Bounias, M. Coevolutionary toxicity as suggested by differential coniferyl alcohol inhibition of ceratocystis species growth. Toxicon 2001, 39, 203–208. [Google Scholar] [CrossRef]
- Kalinova, J.; Triska, J.; Vrchotova, N. Occurence of eugenol, coniferyl alcohol and 3,4,5-trimethoxyphenol in common buck-wheat (Fagopyrum esculentum Moench) and their biological activity. Acta Physiol. Plant. 2011, 33, 1679–1685. [Google Scholar] [CrossRef]
- de Oliveira Silva, E.; Batista, R. Ferulic acid and naturally occurring compounds bearing a feruloyl moiety: A review on their structures, cccurrence, and potential health benefits. Compr. Rev. Food Sci. Food Saf. 2017, 16, 580–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammed, F.S.; Pehlivan, M.; Sevindik, M. Antioxidant, antibacterial and antifungal activities of different extracts of Silybum marianum collected from Duhok (Iraq). Int. J. Second. Metab. 2019, 317–322. [Google Scholar] [CrossRef]
- Yun, D.G.; Lee, D.G. Silymarin exerts antifungal effects via membrane-targeted mode of action by increasing permeability and inducing oxidative stress. Biochim. Biophys. Acta Biomembr. 2017, 1859, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Fuzzati, N. Phenylpropane derivatives from roots of Cosmos caudatus. Phytochemistry 1995, 39, 409–412. [Google Scholar] [CrossRef]
- Tawata, S.; Taira, S.; Kobamoto, N.; Zhu, J.; Ishihara, M.; Toyama, S. Synthesis and antifungal activity of cinnamic acid esters. Biosci. Biotechnol. Biochem. 2014, 60, 909–910. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.-s.; Luo, J.; Raza, W.; Liu, Y.-x.; Gu, M.; Chen, G.; Hu, X.-f.; Wang, J.-h.; Mao, Z.-s.; Shen, Q.-r. Effect of exogenously added ferulic acid on in vitro Fusarium oxysporum f. sp. niveum. Sci. Hortic. 2010, 124, 448–453. [Google Scholar] [CrossRef]
- Barral, B.; Chillet, M.; Minier, J.; Léchaudel, M.; Schorr-Galindo, S. Evaluating the response to Fusarium ananatum inoculation and antifungal activity of phenolic acids in pineapple. Fungal Biol. 2017, 121, 1045–1053. [Google Scholar] [CrossRef]
- Ribes, S.; Fuentes, A.; Talens, P.; Barat, J.M. Combination of different antifungal agents in oil-in-water emulsions to control strawberry jam spoilage. Food Chem. 2018, 239, 704–711. [Google Scholar] [CrossRef]
- Liu, H.; Zhao, H.; Lyu, L.; Huang, Z.; Fan, S.; Wu, W.; Li, W. Synergistic effect of natural antifungal agents for postharvest diseases of blackberry fruits. J. Sci. Food Agric. 2019, 99, 3343–3349. [Google Scholar] [CrossRef]
- Khatkar, A.; Nanda, A.; Kumar, P.; Narasimhan, B. Synthesis and antimicrobial evaluation of ferulic acid derivatives. Res. Chem. Intermed. 2013, 41, 299–309. [Google Scholar] [CrossRef]
- Mahiwal, K.; Kumar, P.; Narasimhan, B. Synthesis, antimicrobial evaluation, ot-QSAR and mt-QSAR studies of 2-amino benzoic acid derivatives. Med. Chem. Res. 2010, 21, 293–307. [Google Scholar] [CrossRef]
- Batista, E.; Lopes, A.; Alves, A. What do we know about Botryosphaeriaceae? An overview of a worldwide cured dataset. Forests 2021, 12, 313. [Google Scholar] [CrossRef]
- Lambert, C.; Bisson, J.; Waffo-Teguo, P.; Papastamoulis, Y.; Richard, T.; Corio-Costet, M.F.; Merillon, J.M.; Cluzet, S. Phenolics and their antifungal role in grapevine wood decay: Focus on the Botryosphaeriaceae family. J. Agric. Food. Chem. 2012, 60, 11859–11868. [Google Scholar] [CrossRef]
- Gómez, P.; Báidez, A.G.; Ortuño, A.; del Río, J.A. Grapevine xylem response to fungi involved in trunk diseases. Ann. Appl. Biol. 2016, 169, 116–124. [Google Scholar] [CrossRef]
- Dekker, R.F.H.; Barbosa, A.M.; Sargent, K. The effect of lignin-related compounds on the growth and production of laccases by the ascomycete, Botryosphaeria sp. Enzym. Microb. Technol. 2002, 30, 374–380. [Google Scholar] [CrossRef]
- Mondello, V.; Songy, A.; Battiston, E.; Pinto, C.; Coppin, C.; Trotel-Aziz, P.; Clement, C.; Mugnai, L.; Fontaine, F. Grapevine trunk diseases: A review of fifteen years of trials for their control with chemicals and biocontrol agents. Plant Dis. 2018, 102, 1189–1217. [Google Scholar] [CrossRef] [Green Version]
- Shakeel, F.; Salem-Bekhit, M.M.; Haq, N.; Siddiqui, N.A. Solubility and thermodynamics of ferulic acid in different neat sol-vents: Measurement, correlation and molecular interactions. J. Mol. Liq. 2017, 236, 144–150. [Google Scholar] [CrossRef]
- Nguyen, T.T.H.; Si, J.; Kang, C.; Chung, B.; Chung, D.; Kim, D. Facile preparation of water soluble curcuminoids extracted from turmeric (Curcuma longa L.) powder by using steviol glucosides. Food Chem. 2017, 214, 366–373. [Google Scholar] [CrossRef] [Green Version]
- Martinelli, T.; Andrzejewska, J.; Salis, M.; Sulas, L. Phenological growth stages of Silybum marianum according to the extended BBCH scale. Ann. Appl. Biol. 2015, 166, 53–66. [Google Scholar] [CrossRef]
- Martin, M.T.; Cobos, R. Identification of fungi associated with grapevine decline in Castilla y León (Spain). Phytopathol. Mediterr. 2007, 46, 18–25. [Google Scholar]
- Nunn, S.; Nishikida, K. Advanced ATR Correction Algorithm—Application Note 50581; ThermoScientific: Madison, WI, USA, 2008; p. 4. [Google Scholar]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectorscopy, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007; p. 804. [Google Scholar]
- Buzón-Durán, L.; Martín-Gil, J.; Pérez-Lebeña, E.; Ruano-Rosa, D.; Revuelta, J.L.; Casanova-Gascón, J.; Ramos-Sánchez, M.C.; Martín-Ramos, P. Antifungal agents based on chitosan oligomers, ε-polylysine and Streptomyces spp. secondary metabolites against three Botryosphaeriaceae species. Antibiotics 2019, 8, 99. [Google Scholar] [CrossRef] [Green Version]
- Arendrup, M.C.; Cuenca-Estrella, M.; Lass-Flörl, C.; Hope, W. EUCAST technical note on the EUCAST definitive document EDef 7.2: Method for the determination of broth dilution minimum inhibitory concentrations of antifungal agents for yeasts EDef 7.2 (EUCAST-AFST). Clin. Microbiol. Infect. 2012, 18, E246–E247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, Y.; Benderly, M.; Cohen, Y.; Gisi, U.; Bassand, D. The joint action of fungicides in mixtures: Comparison of two methods for synergy calculation. EPPO Bull. 1986, 16, 651–657. [Google Scholar] [CrossRef]
- Aroca, Á.; Gramaje, D.; Armengol, J.; García-Jiménez, J.; Raposo, R. Evaluation of the grapevine nursery propagation process as a source of Phaeoacremonium spp. and Phaeomoniella chlamydospora and occurrence of trunk disease pathogens in root-stock mother vines in Spain. Eur. J. Plant Pathol. 2010, 126, 165–174. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Lv, Y.; Cheng, X.; Wu, D.; Du, G.; Zhou, J.; Chen, J. Improving bioconversion of eugenol to coniferyl alcohol by in situ elimi-nating harmful H2O2. Bioresour. Technol. 2018, 267, 578–583. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Xu, S.; Lyu, Y.; Zhou, S.; Du, G.; Chen, J.; Zhou, J. Engineering enzymatic cascades for the efficient biotransformation of eugenol and taxifolin to silybin and isosilybin. Green Chem. 2019, 21, 1660–1667. [Google Scholar] [CrossRef]
- Zainal-Abidin, M.H.; Hayyan, M.; Hayyan, A.; Jayakumar, N.S. New horizons in the extraction of bioactive compounds using deep eutectic solvents: A review. Anal. Chim. Acta 2017, 979, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.V.; Reis, R.A.S.P.; Garbuio, D.C.; de Freitas, L.A.P. Dynamic maceration of Matricaria chamomilla inflorescences: Optimal conditions for flavonoids and antioxidant activity. Rev. Bras. Farmacogn. 2018, 28, 111–117. [Google Scholar] [CrossRef]
- Qi, X.-L.; Peng, X.; Huang, Y.-Y.; Li, L.; Wei, Z.-F.; Zu, Y.-G.; Fu, Y.-J. Green and efficient extraction of bioactive flavonoids from Equisetum palustre L. by deep eutectic solvents-based negative pressure cavitation method combined with macroporous resin enrichment. Ind. Crop. Prod. 2015, 70, 142–148. [Google Scholar] [CrossRef]
- Civolani, C.; Barghini, P.; Roncetti, A.R.; Ruzzi, M.; Schiesser, A. Bioconversion of ferulic acid into vanillic acid by means of a vanillate-negative mutant of Pseudomonas fluorescens strain BF13. Appl. Environ. Microbiol. 2000, 66, 2311–2317. [Google Scholar] [CrossRef] [Green Version]
- Oddou, J.; Stentelaire, C.; Lesage-Meessen, L.; Asther, M.; Colonna Ceccaldi, B. Improvement of ferulic acid bioconversion into vanillin by use of high-density cultures of Pycnoporus cinnabarinus. Appl. Microbiol. Biotechnol. 1999, 53, 1–6. [Google Scholar] [CrossRef]
- Barghini, P.; di Gioia, D.; Fava, F.; Ruzzi, M. Vanillin production using metabolically engineered Escherichia coli under non-growing conditions. Microb. Cell Factories 2007, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luziatelli, F.; Brunetti, L.; Ficca, A.G.; Ruzzi, M. Maximizing the efficiency of vanillin production by biocatalyst enhancement and process optimization. Front. Bioeng. Biotechnol. 2019, 7, 279. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Chen, P.; Zhang, S.; Li, S.; Yan, X.; Wang, N.; Liang, N.; Li, H. Biotransformation of ferulic acid to vanillin in the packed bed-stirred fermentors. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- Sulas, L.; Ventura, A.; Murgia, L. Phytomass production from Silybum marianum for bioenergy. In Sustainable Mediterranean Grasslands and Their Multi-Functions; Porqueddu, C., Tavares de Sousa, M.M., Eds.; Mediterranean Agronomic Institute of Zaragoza: Zaragoza, Spain, 2008; Volume 79, pp. 487–490. [Google Scholar]
- Domínguez, M.T.; Madejón, P.; Madejón, E.; Diaz, M.J. Novel energy crops for Mediterranean contaminated lands: Valoriza-tion of Dittrichia viscosa and Silybum marianum biomass by pyrolysis. Chemosphere 2017, 186, 968–976. [Google Scholar] [CrossRef] [PubMed]
- Hunce, S.Y.; Clemente, R.; Bernal, M.P. Energy production potential of phytoremediation plant biomass: Helianthus annuus and Silybum marianum. Ind. Crop. Prod. 2019, 135, 206–216. [Google Scholar] [CrossRef]
- Ghosh, S.; Subudhi, E.; Nayak, S. Antimicrobial assay of Stevia rebaudiana Bertoni leaf extracts against 10 pathogens. Int. J. Integr. Biol. 2008, 2, 27–31. [Google Scholar]
- Abou-Arab, E.; Abu-Salem, F. Evaluation of bioactive compounds of Stevia rebaudiana leaves and callus. J. Food Dairy Sci. 2010, 1, 209–224. [Google Scholar] [CrossRef]
- Arya, A.; Kumar, S.; Kasana, M. In vitro regeneration of Stevia and evaluation of antimicrobial and antiprotozoal properties of regenerated calli and plants. Electron. J. Plant Breed. 2012, 3, 916–924. [Google Scholar]
- Shukla, S.; Mehta, A.; Bajpai, V.K. Phytochemical screening and anthelmintic and antifungal activities of leaf extracts of Stevia rebaudiana. J. Biol. Act. Prod. Nat. 2013, 3, 56–63. [Google Scholar] [CrossRef]
- Guerra Ramírez, P.; Guerra Ramírez, D.; Zavaleta Mejía, E.; Aranda Ocampo, S.; Nava Díaz, C.; Rojas Martínez, R.I. Extracts of Stevia rebaudiana against Fusarium oxysporum associated with tomato cultivation. Sci. Hortic. 2020, 259, 108683. [Google Scholar] [CrossRef]
- Yun, D.G.; Lee, D.G. Assessment of silibinin as a potential antifungal agent and investigation of its mechanism of action. IUBMB Life 2017, 69, 631–637. [Google Scholar] [CrossRef] [Green Version]
- Janeczko, M.; Kochanowicz, E. Silymarin, a popular dietary supplement shows anti–Candida activity. Antibiotics 2019, 8, 206. [Google Scholar] [CrossRef] [Green Version]
- Fernández, A.; Colombo, M.L.; Curto, L.M.; Gómez, G.E.; Delfino, J.M.; Guzmán, F.; Bakás, L.; Malbrán, I.; Vairo-Cavalli, S.E. Peptides derived from the α-core and γ-core regions of a putative Silybum marianum flower defensin show antifungal activity against Fusarium graminearum. Front. Microbiol. 2021, 12, 632008. [Google Scholar] [CrossRef]
- Safarpoor, M.; Ghaedi, M.; Asfaram, A.; Yousefi-Nejad, M.; Javadian, H.; Zare Khafri, H.; Bagherinasab, M. Ultra-sound-assisted extraction of antimicrobial compounds from Thymus daenensis and Silybum marianum: Antimicrobial activity with and without the presence of natural silver nanoparticles. Ultrason. Sonochem. 2018, 42, 76–83. [Google Scholar] [CrossRef]
- Keskin, D.; Oskay, D.; Oskay, M. Antimicrobial activity of selected plant spices marketed in the West Anatolia. Int. J. Agric. Biol. 2010, 12, 916–920. [Google Scholar]
- Kuc, J. Induced systemic resistance in plants to diseases caused by fungi and bacteria. In The Dynamics of Host Defence; Bailey, J., Deverall, B., Eds.; Academic Press: New York, NY, USA, 1983; pp. 191–221. [Google Scholar]
- Hammerschmidt, R.; Kuć, J. Lignification as a mechanism for induced systemic resistance in cucumber. Physiol. Plant Pathol. 1982, 20, 61–71. [Google Scholar] [CrossRef]
- Keen, N.T.; Littlefield, L.J. The possible association of phytoalexins with resistance gene expression in flax to Melampsora lini. Physiol. Plant Pathol. 1979, 14, 265–280. [Google Scholar] [CrossRef]
- Lambert, C.; Papastamoulis, Y.; Waffo-Téguo, P.; Mérillon, J.; Cluzet, S. Effects of phenolic compounds towards grapevine wood decay fungi. Proceedings of Oeno2011: Actes de colloques du 9e symposium international d’oenologie de Bordeaux, Bordeaux, France, 15–17 June 2011; pp. 90–94. [Google Scholar]
- Srivastava, P.; Andersen, P.C.; Marois, J.J.; Wright, D.L.; Srivastava, M.; Harmon, P.F. Effect of phenolic compounds on growth and ligninolytic enzyme production in Botryosphaeria isolates. Crop Prot. 2013, 43, 146–156. [Google Scholar] [CrossRef]
- Buzón-Durán, L.; Martín-Gil, J.; Ramos-Sánchez, M.d.C.; Pérez-Lebeña, E.; Marcos-Robles, J.L.; Fombellida-Villafruela, Á.; Martín-Ramos, P. Antifungal activity against Fusarium culmorum of stevioside, Silybum marianum seed extracts, and their conjugate complexes. Antibiotics 2020, 9, 440. [Google Scholar] [CrossRef] [PubMed]
- Matei, P.; Iacomi, B.; Martín-Gil, J.; Pérez-Lebeña, E.; Ramos-Sánchez, M.; Barrio-Arredondo, M.; Martín-Ramos, P. In vitro antifungal activity of composites of AgNPs and polyphenol inclusion compounds against Fusarium culmorum in different dis-persion media. Agronomy 2018, 8, 239. [Google Scholar] [CrossRef] [Green Version]
- Matei, P.; Martín-Gil, J.; Michaela Iacomi, B.; Pérez-Lebeña, E.; Barrio-Arredondo, M.; Martín-Ramos, P. Silver nanoparticles and polyphenol inclusion compounds composites for Phytophthora cinnamomi mycelial growth inhibition. Antibiotics 2018, 7, 76. [Google Scholar] [CrossRef] [Green Version]
- Matei, P.M.; Buzón-Durán, L.; Pérez-Lebeña, E.; Martín-Gil, J.; Iacomi, B.M.; Ramos-Sánchez, M.C.; Martín-Ramos, P. In vitro antifungal activity of chitosan-polyphenol conjugates against Phytophthora cinnamomi. AgriEngineering 2020, 2, 72–77. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Code | Isolate | Binomial Nomenclature | Geographical Origin | Host/Date |
|---|---|---|---|---|
| ITACYL_F111 | Y-091-03-01c | Neofusicoccum parvum (Pennycook & Samuels) Crous, Slippers & A.J.L.Phillips | Spain (Navarra, nursery) | Grapevine (‘Verdejo’) 2006 |
| ITACYL_F118 | Y-103-08-01 | Dothiorella viticola A.J.L.Phillips & J.Luque | Spain (Extremadura) | Grapevine 2004 |
| ITACYL_F098 | Y-084-01-01a | Diplodia seriata De Not. | Spain (DO Toro) | Grapevine (‘Tempranillo’) 2004 |
| Silybum marianum | Silymarin | Ferulic Acid | Assignments |
|---|---|---|---|
| 3335 | 3331 | OH group in phenolic compounds | |
| 3279 | |||
| 3069 | |||
| 2918 | 2932 | 2926 | O–H stretching |
| 1651 1634 | 1649 | skeletal vibration due to aromatic C=C ring stretching and C=O stretching | |
| 1602 | 1605 | aromatic C=C stretching | |
| 1558 | >C=C< aromatic | ||
| 1515 1457 | 1458 | 1510 | symmetric aromatic ring stretching vibration (C=C ring) |
| 1429 | 1434 | olefinic C–H | |
| 1313 | 1329 | C–H vibration of the methyl group | |
| 1275 | Carboxylic acid C=O stretching | ||
| 1242 | 1257 | ||
| 1126 | in plane =C–H bending/C=C stretching | ||
| 1030 | 1076 | C–O stretching/O-H out plane bending | |
| 941 | |||
| 779 | 721 | 693 | C=C on the aromatic ring methylene rocking vibration |
| Peak | Rt (min) | Area (%) | Tentative Assignments |
|---|---|---|---|
| 1 | 4.8755 | 2.67 | methoxy-phenyl-oxime |
| 2 | 6.0099 | 3.50 | glycerin |
| 3 | 7.3293 | 2.62 | hexamethyl-cyclotrisiloxane; tris(tert-butyldimethylsilyloxy)arsane |
| 4 | 7.6360 | 7.31 | 2-methoxy-phenol |
| 5 | 9.4764 | 2.48 | 2,3-dihydro-benzofurane |
| 6 | 9.6516 | 1.95 | methenamine |
| 7 | 10.8737 | 3.92 | 4-hydroxy-3-methylacetophenone |
| 8 | 12.0275 | 1.64 | vanillin |
| 9 | 12.6653 | 1.51 | trans-isoeugenol |
| 10 | 13.0548 | 1.36 | 6-methoxy-3-methylbenzofuran |
| 11 | 15.3139 | 1.69 | 4-((1E)-3-hydroxy-1-propenyl)-2-methoxyphenol (also named coniferol or γ-hydroxyisoeugenol) |
| 12 | 15.5865 | 0.82 | 2-hydroxy-4-isopropyl-7-methoxytropone |
| 13 | 15.9370 | 1.68 | 4-hydroxy-3-methoxybenzeneacetic acid, -, methyl ester |
| 14 | 16.0636 | 45.64 | 4-((1E)-3-Hydroxy-1-propenyl)-2-methoxyphenol (also named coniferol or γ-hydroxyisoeugenol) |
| 15 | 17.1153 | 14.99 | 2-propenoic acid, 3-(4-hydroxy-3-methoxyphenyl)-, methyl ester (also named ferulic acid methyl ester) |
| 16 | 17.9234 | 0.49 | ethyl (2E)-3-(4-hydroxy-3-methoxyphenyl)-2-propenoate |
| 17 | 19.5447 | 0.67 | 9,15-octadecadienoic acid, methyl ester, (Z,Z)- |
| 18 | 21.1027 | 2.93 | 2-(1,4,4-trimethyl-cyclohex-2-enyl)-ethanol |
| 19 | 24.4377 | 2.13 | 9,12-octadecadienoic acid (Z,Z)-, 2,3-dihydroxypropyl ester |
| Pathogen | EC | Stevioside | S. marianum | Stevioside– S. marianum | Coniferyl Alcohol | Stevioside– Coniferyl Alcohol | Ferulic Acid | Stevioside– Ferulic Acid |
|---|---|---|---|---|---|---|---|---|
| N. parvum | EC50 | 152.2 ± 13.4 | 677.2 ± 47.0 | 89.2 ± 15.3 | 214.3 ± 26.2 | 157.8 ± 16.6 | 1394.5 ± 63.0 | 465.9 ± 27.51 |
| EC90 | 824.1 ± 56.7 | 2938.3 ± 286.6 | 262.1 ± 19.2 | 1005.1 ± 71.3 | 384.9 ± 22.8 | 2948.6 ± 268.1 | 1132.7 ± 127.3 | |
| D. viticola | EC50 | 271.4 ± 26.6 | 1088.4 ± 93.8 | 148.3 ± 11.7 | 361.1 ± 38.8 | 156.5 ± 8.3 | 1387.2 ± 134.3 | 544.5 ± 24.4 |
| EC90 | 1017.0 ± 74.3 | 9943.2 ± 1038.6 | 360.7 ± 39.0 | 988.5 ± 88.6 | 368.2 ± 26.6 | 3921.3 ± 438.6 | 1183.2 ± 111.0 | |
| D. seriata | EC50 | 230.1 ± 15.3 | 703.0 ± 26.6 | 127.1 ± 15.5 | 370.3 ± 10.4 | 191.6 ± 12.6 | 433.0 ± 31.5 | 209.0 ± 18.1 |
| EC90 | 840.5 ± 62.3 | 1461.1 ± 111.8 | 355.4 ± 38.1 | 913.2 ± 65.6 | 360.5 ± 29.6 | 903.4 ± 74.4 | 465.9 ± 33.2 |
| Effective Concentration | N. parvum | D. viticola | D. seriata |
|---|---|---|---|
| EC50 | 2.8 | 2.9 | 2.7 |
| EC90 | 4.9 | 5.1 | 3.0 |
| Pathogen | Sample | Frequency | Sum of Ranks | Mean of Ranks | Groups | ||
|---|---|---|---|---|---|---|---|
| N. parvum | Stevioside–S. marianum negative control | 48 | 1275.500 | 26.573 | A | ||
| Stevioside–S. marianum | 64 | 5911.000 | 92.359 | B | |||
| Positive control | 64 | 8389.500 | 131.086 | C | |||
| D. viticola | Stevioside–S. marianum negative control | 48 | 2174.000 | 45.292 | A | ||
| Stevioside–S. marianum | 64 | 4272.000 | 66.750 | B | |||
| Positive control | 64 | 9130.000 | 142.656 | C | |||
| D. seriata | Stevioside–S. marianum negative control | 48 | 2062.500 | 42.969 | A | ||
| Stevioside–S. marianum | 72 | 5641.500 | 78.354 | B | |||
| Positive control | 56 | 7872.000 | 140.571 | C | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Langa-Lomba, N.; Buzón-Durán, L.; Sánchez-Hernández, E.; Martín-Ramos, P.; Casanova-Gascón, J.; Martín-Gil, J.; González-García, V. Antifungal Activity against Botryosphaeriaceae Fungi of the Hydro-Methanolic Extract of Silybum marianum Capitula Conjugated with Stevioside. Plants 2021, 10, 1363. https://doi.org/10.3390/plants10071363
Langa-Lomba N, Buzón-Durán L, Sánchez-Hernández E, Martín-Ramos P, Casanova-Gascón J, Martín-Gil J, González-García V. Antifungal Activity against Botryosphaeriaceae Fungi of the Hydro-Methanolic Extract of Silybum marianum Capitula Conjugated with Stevioside. Plants. 2021; 10(7):1363. https://doi.org/10.3390/plants10071363
Chicago/Turabian StyleLanga-Lomba, Natalia, Laura Buzón-Durán, Eva Sánchez-Hernández, Pablo Martín-Ramos, José Casanova-Gascón, Jesús Martín-Gil, and Vicente González-García. 2021. "Antifungal Activity against Botryosphaeriaceae Fungi of the Hydro-Methanolic Extract of Silybum marianum Capitula Conjugated with Stevioside" Plants 10, no. 7: 1363. https://doi.org/10.3390/plants10071363
APA StyleLanga-Lomba, N., Buzón-Durán, L., Sánchez-Hernández, E., Martín-Ramos, P., Casanova-Gascón, J., Martín-Gil, J., & González-García, V. (2021). Antifungal Activity against Botryosphaeriaceae Fungi of the Hydro-Methanolic Extract of Silybum marianum Capitula Conjugated with Stevioside. Plants, 10(7), 1363. https://doi.org/10.3390/plants10071363