
Responses to Salinity in Four Plantago Species from Tunisia


 ,
,  ,
, 

 and
and
Abstract
:1. Introduction
2. Results
2.1. Substrate Analysis
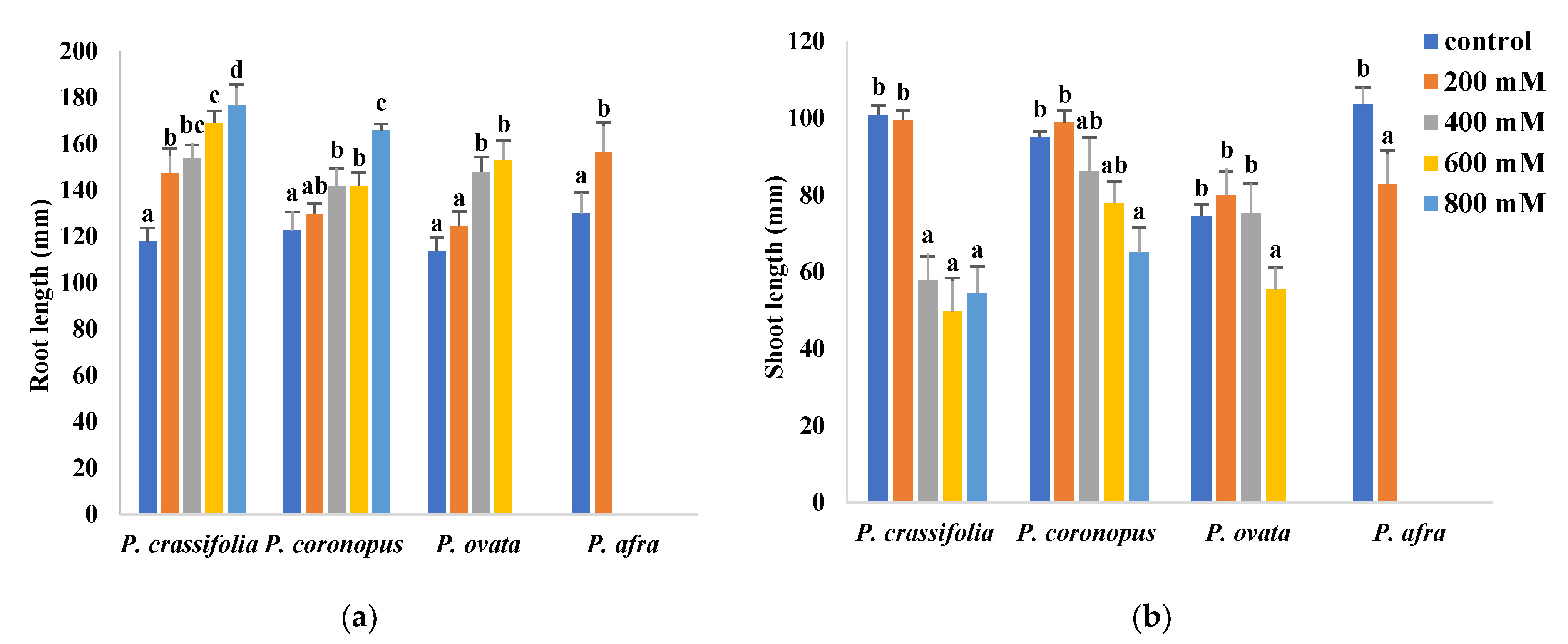
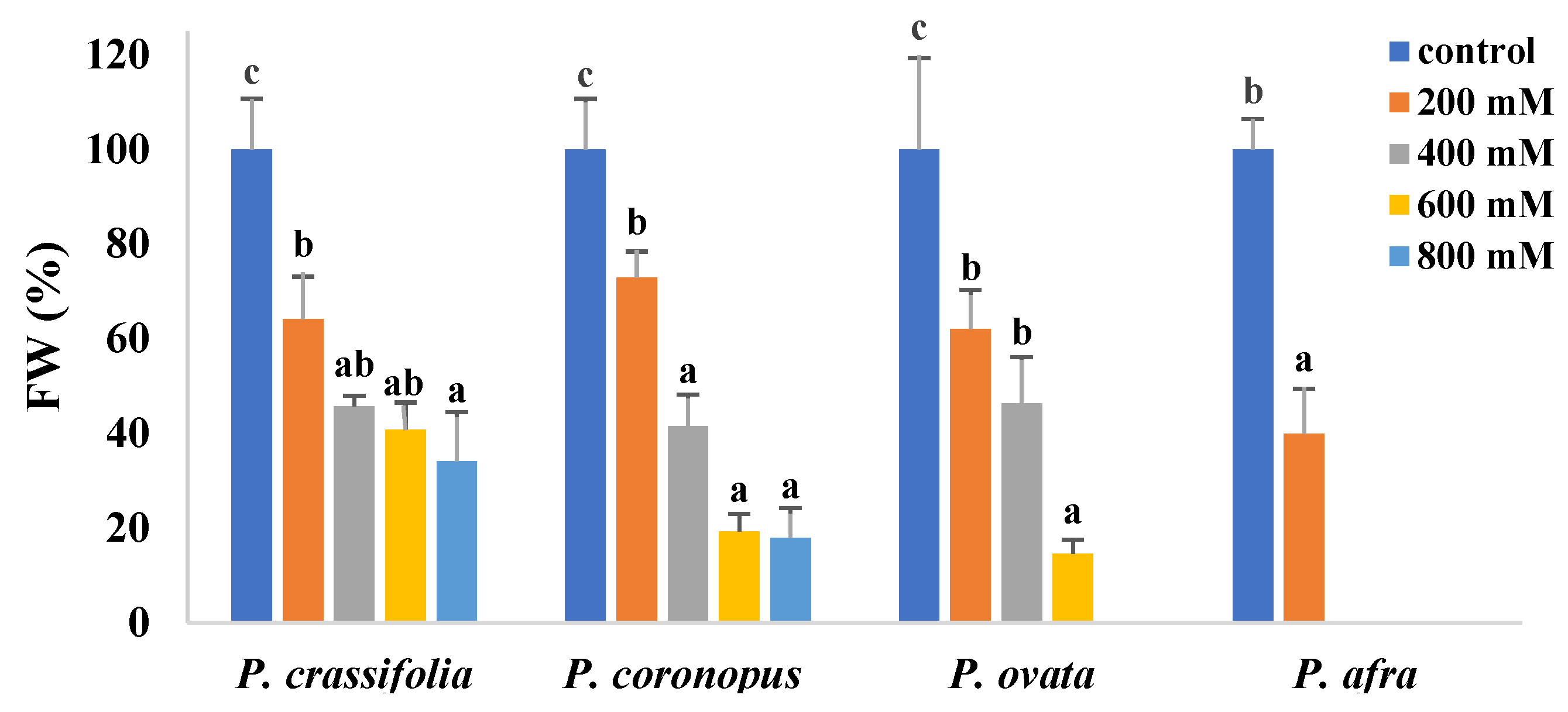
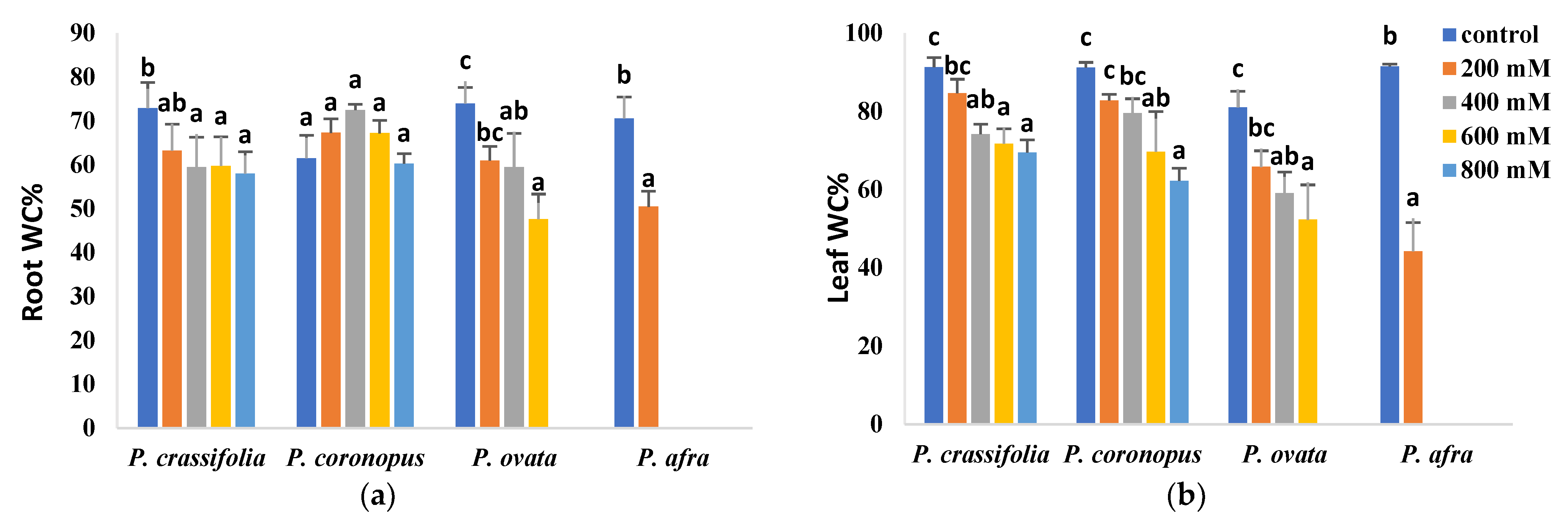
2.2. Effects of Salt Stress on Plant Growth
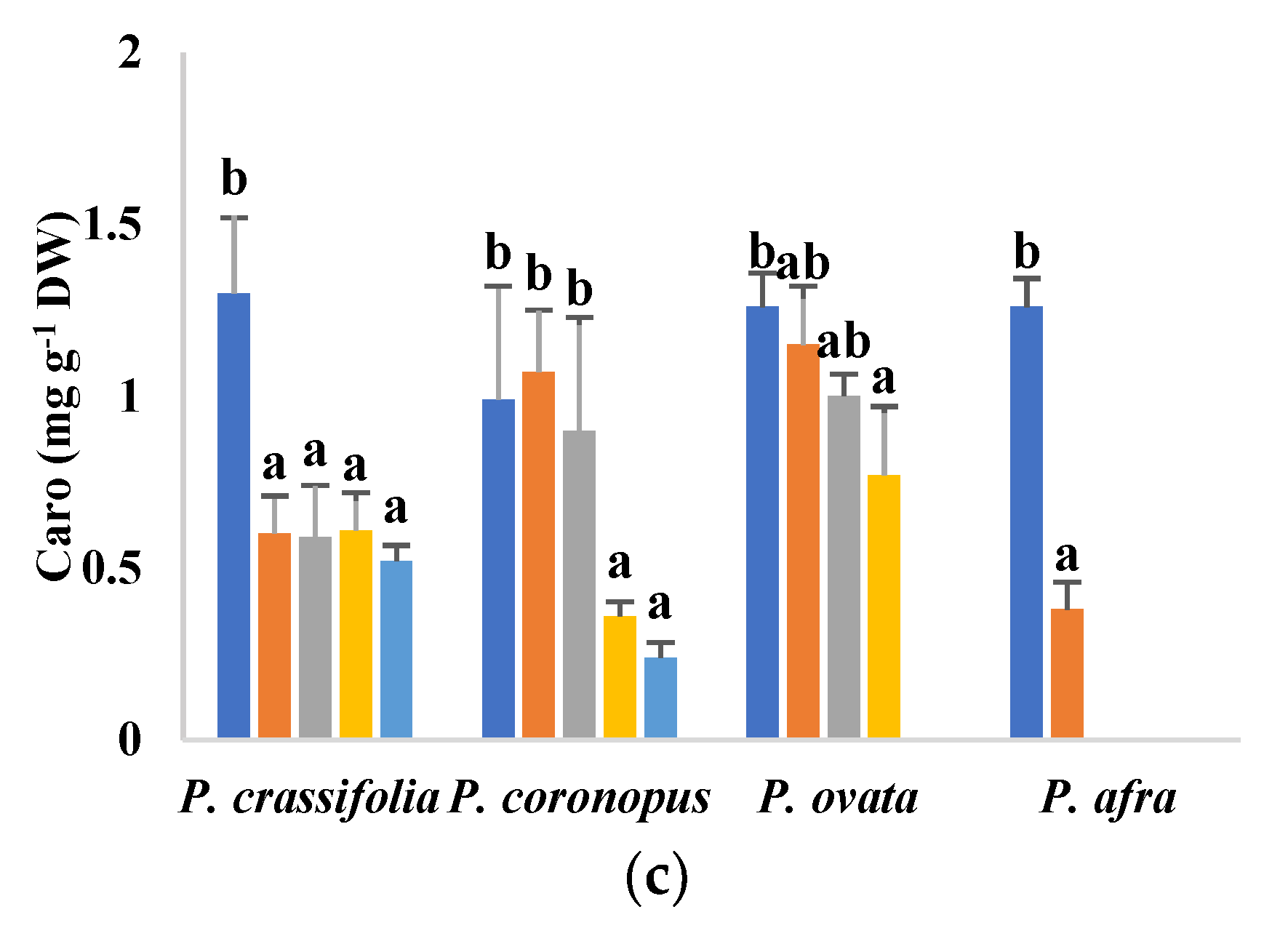
2.3. Effects of Salt Stress on Photosynthetic Pigment Levels
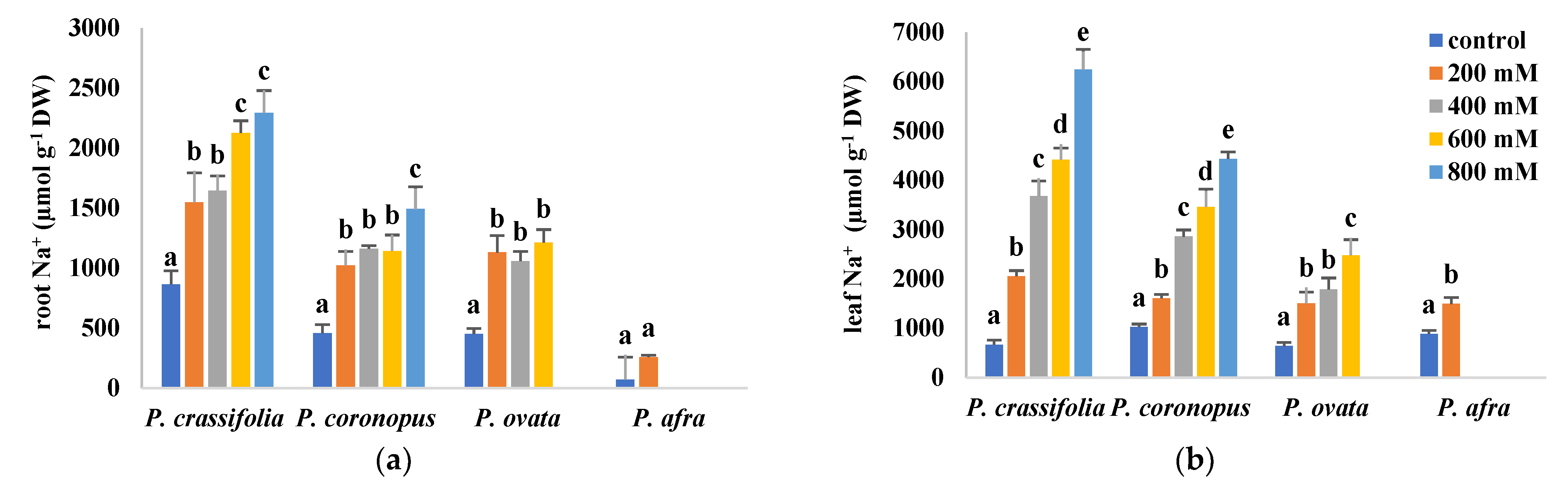
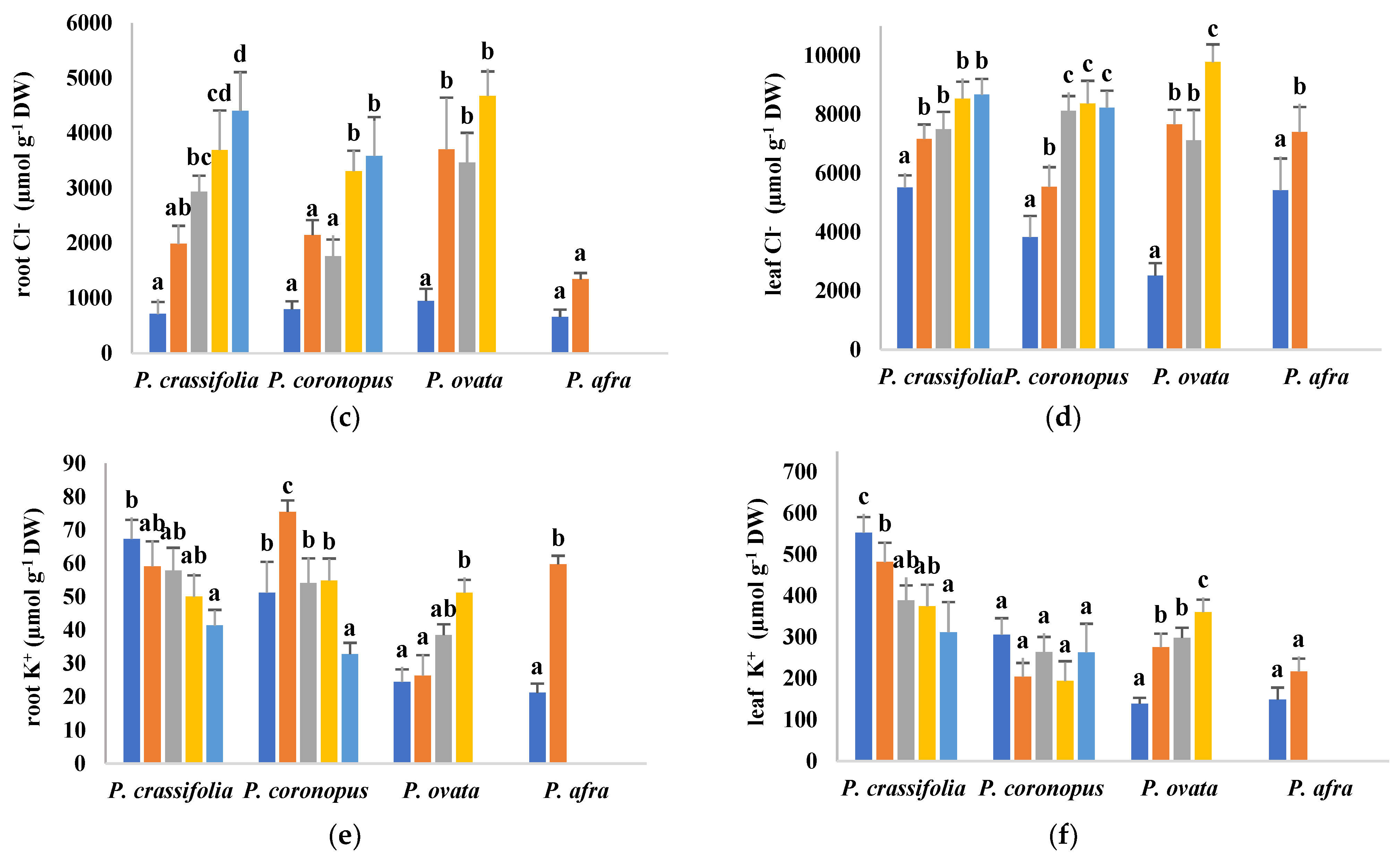
2.4. Ion Accumulation
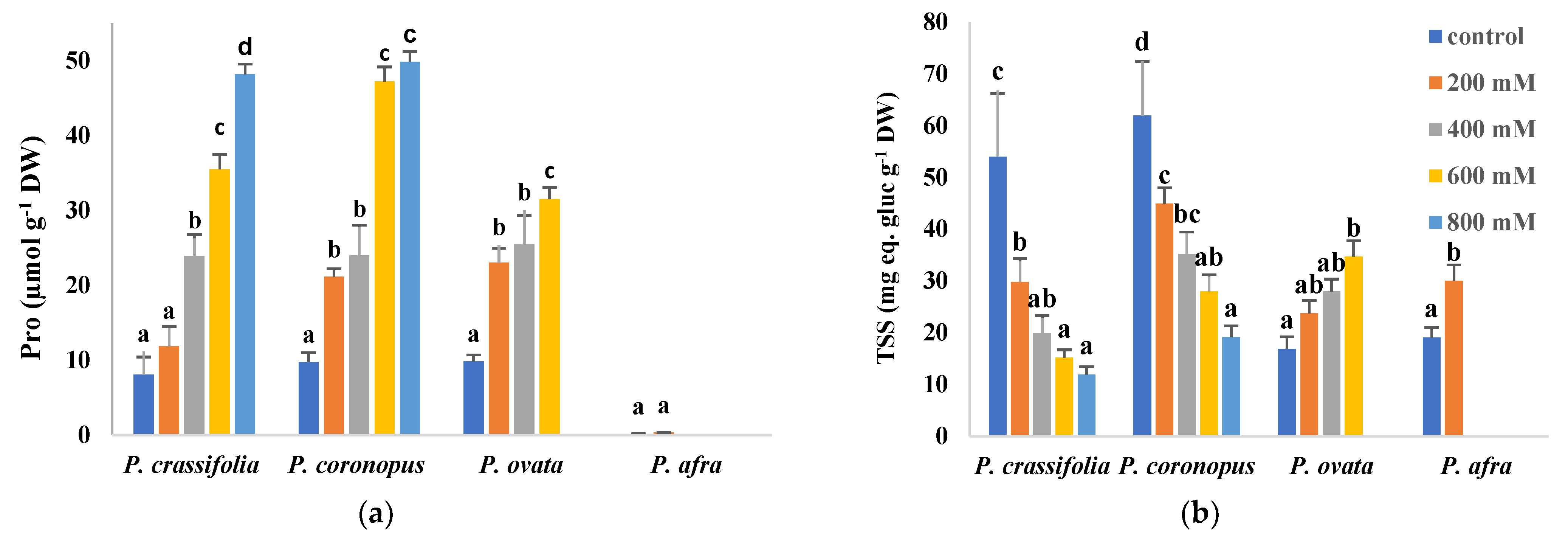
2.5. Salt Stress Effect on Osmolyte Contents
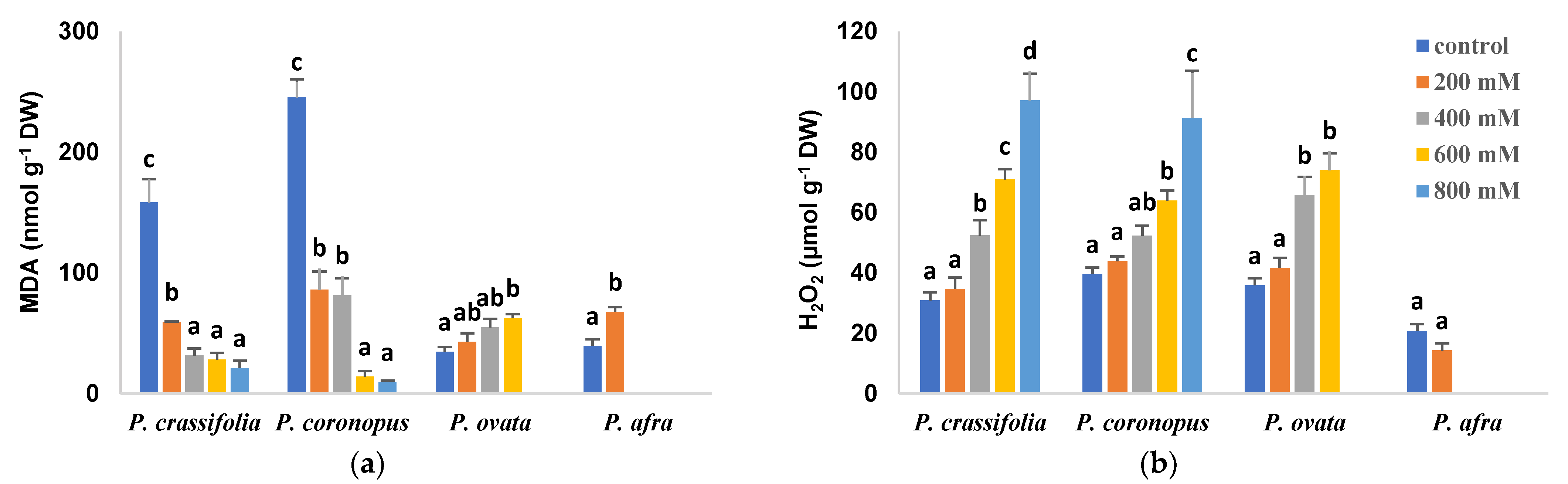
2.6. Oxidative Stress Biochemical Markers
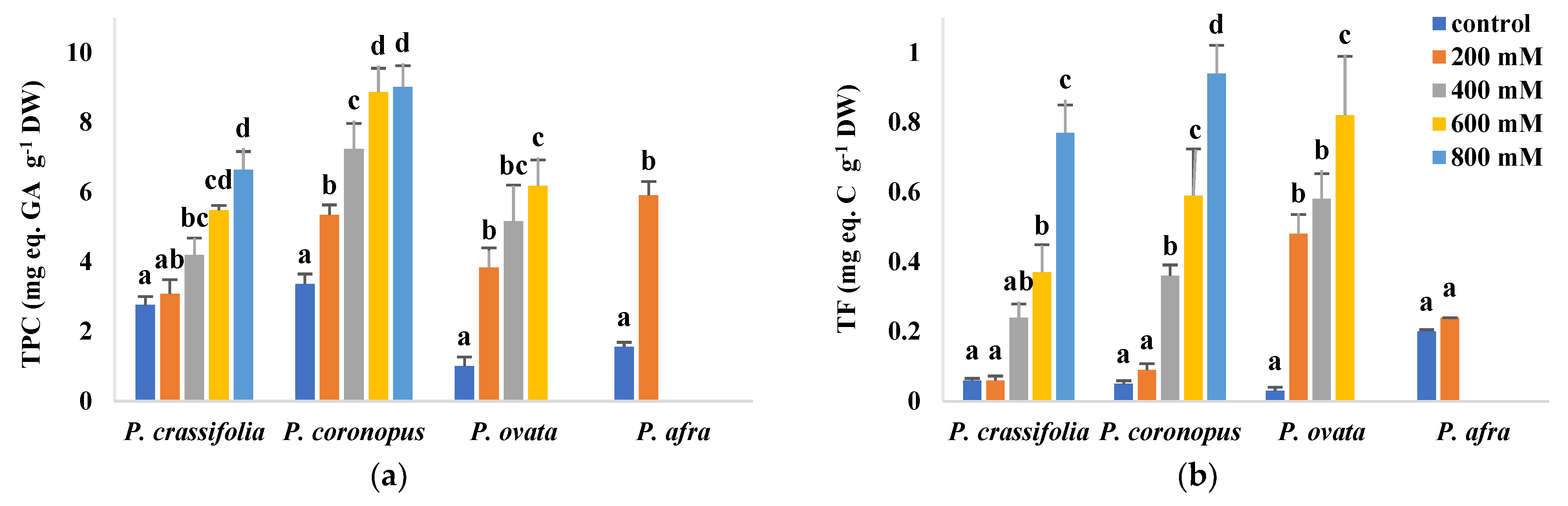
2.7. Antioxidant Compounds
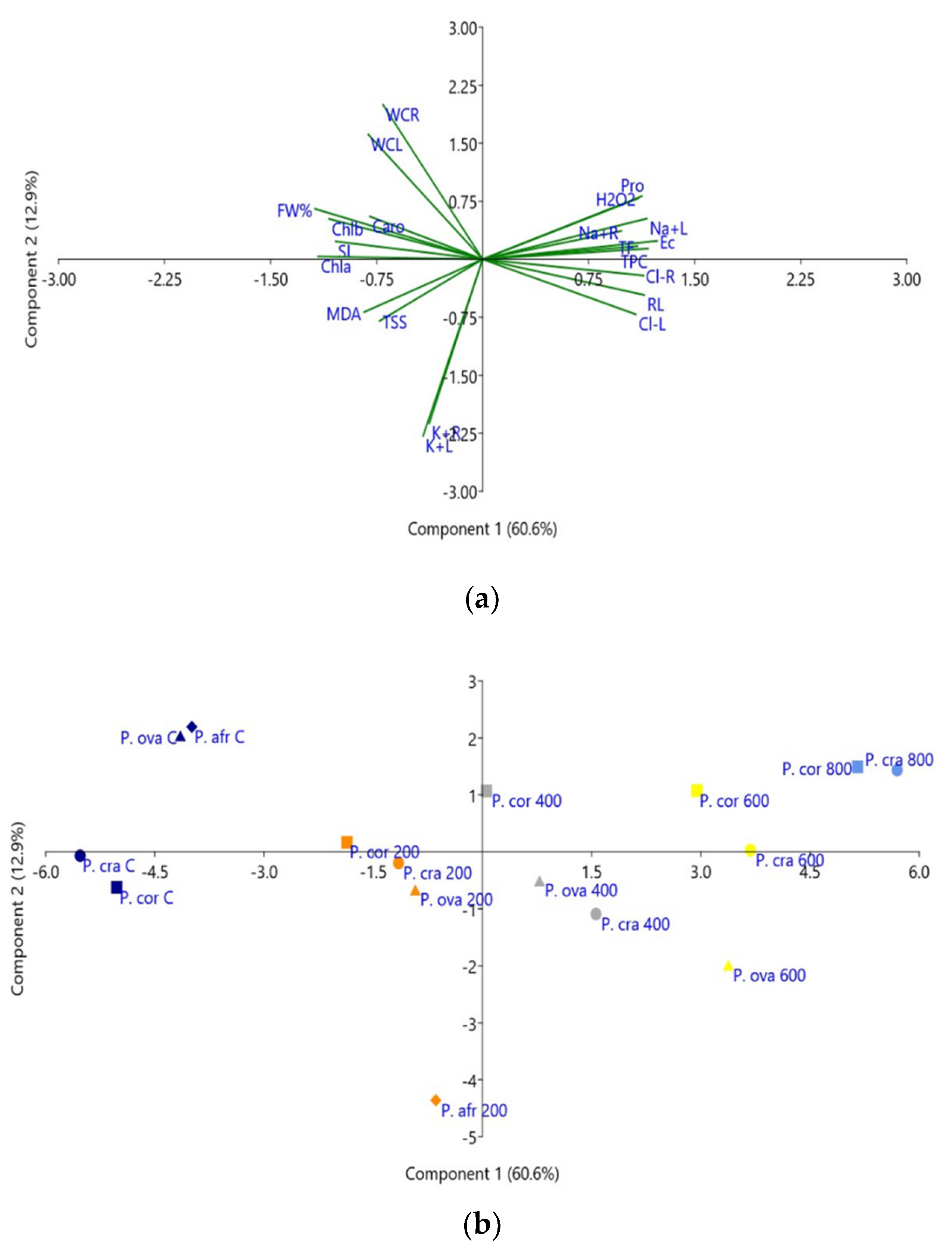
2.8. Principal Component Analysis
3. Discussion
4. Material and Methods
4.1. Plant Material
4.2. Plant Growth, Salt Treatments and Plant Sampling
4.3. Electrical Conductivity of the Substrate
4.4. Photosynthetic Pigments Determination
4.5. Ion Content Measurements
4.6. Proline and Total Soluble Sugars Quantification
4.7. Oxidative Stress Markers
4.8. Non-Enzymatic Antioxidants
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parvaiz, A.; Satyawati, S. Salt stress and phyto-biochemical responses of plants—A review. Plant Soil Environ. 2008, 54, 88–99. [Google Scholar] [CrossRef]
- Arzani, A.; Ashraf, M. Smart engineering of genetic resources for enhanced salinity tolerance in crop plants. Crit. Rev. Plant Sci. 2016, 35, 146–189. [Google Scholar] [CrossRef]
- Shahid, S.A.; Zaman, M.; Heng, L. Soil salinity: Historical perspectives and a world overview of the problem. In Guideline for Salinity Assessment, Mitigation and Adaptation Using Nuclear and Related Techniques; Zaman, M., Shahid, S.A., Heng, L., Eds.; Springer: Cham, Germany, 2018. [Google Scholar] [CrossRef] [Green Version]
- Pitman, M.G.; Láuchli, A. Global impact of salinity and agricultural ecosystems. In Salinity: Environment-Plants–Molecules; Läuchli, A., Lüttge, U., Eds.; Kluver Academic Publishers: Dordrecht, The Netherlands, 2002. [Google Scholar] [CrossRef]
- IPCC. Intergovernmental panel on climate change. In Proceedings of the 5th Assessment Report, WGII, Climate Change: Impacts, Adaptation, and Vulnerability. Available online: http://www.ipcc.ch/report/ar5/wg2/ (accessed on 4 March 2021).
- Fita, A.; Rodríguez-Burruezo, A.; Boscaiu, M.; Prohens, J.; Vicente, O. Breeding and domesticating crops adapted to drought and salinity: A new paradigm for increasing food production. Front. Plant Sci. 2015, 6, 978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genomics 2014, 701596. [Google Scholar] [CrossRef] [PubMed]
- Isayenkov, S.V.; Maathuis, F.J.M. Plant salinity stress: Many unanswered questions remain. Front. Plant Sci. 2019, 1, 80. [Google Scholar] [CrossRef] [Green Version]
- Van Zelm, E.; Zhang, Y.; Testerink, Y. Salt tolerance mechanisms of plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef] [Green Version]
- Muhammad, Z.; Hussain, F. Effect of NaCl salinity on the germination and seedling growth of some medicinal plants. Pak. J. Bot. 2010, 42, 889–897. [Google Scholar]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Santos, J.; Al-Azzawi, M.; Aronson, J.; Flowers, T.J. eHALOPH a database of salt-tolerant plants: Helping put halophytes to work. Plant Cell Physiol. 2016, 57, e10. [Google Scholar] [CrossRef] [Green Version]
- Hassemer, G.; Shipunov, A.; Rønsted, N.A.H.; Meudt, H.M. Taxonomic and geographic novelties in the genus Plantago (Plantaginaceae) in Chile, including the description of a new species. Phytotaxa 2018, 340, 137–156. [Google Scholar] [CrossRef]
- Hassemer, G.; Bruun-Lund, S.; Shipunov, A.; Briggs, B.G.; Meudt, H.M.; Rønsted, N.A.H. The application of high-throughput sequencing for taxonomy: The case of Plantago subg. Plantago (Plantaginaceae). Mol. Phylogenet. Evol. 2019, 138, 156–173. [Google Scholar] [CrossRef] [PubMed]
- Chater, A.O.; Cartier, D. Plantago L. In Flora Europaea 4, Plantaginaceae to Compositae; Tutin, T.G., Heywood, V.H., Burges, N.A., Valentine., D.H., Eds.; Cambridge University Press: Cambridge, UK, 1976; Volume 4, pp. 38–44. [Google Scholar]
- Ksouri, R.; Ksouri, W.M.; Jallali, I.; Debez, A.; Magné, C.; Hiroko, I.; Abdelly, C. Medicinal halophytes: Potent source of health promoting biomolecules with medical, nutraceutical and food applications. Crit. Rev. Biotechnol. 2012, 32, 289–326. [Google Scholar] [CrossRef]
- Flanagan, L.B.; Jefferies, R.L. Effect of increased salinity on CO2 assimilation, O2 evolution and the δ13C values of leaves of Plantago maritima L. developed at low and high NaC1 levels. Planta 1989, 78, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Maathuis, F.J.; Prins, H.B. Patch clamp studies on root cell vacuoles of a salt-tolerant and a salt-sensitive Plantago species: Regulation of channel activity by salt stress. Plant Physiol. 1990, 92, 23–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staal, M.; Maathuis, F.J.M.; Elzenga, J.T.M.; Overbeek, J.H.M.; Prins, H.B.A. Na+/H+ antiport activity in tonoplast vesicles from roots of the salt-tolerant Plantago maritima and the salt-sensitive Plantago media. Physiol. Plant. 1991, 82, 179–184. [Google Scholar] [CrossRef]
- Al Hassan, M.; Pacurar, A.; López-Gresa, M.P.; Donat-Torres, M.P.; Llinares, J.V.; Boscaiu, M.; Vicente, O. Effects of salt stress on three ecologically distinct Plantago Species. PLoS ONE 2016, 11, e0160236. [Google Scholar] [CrossRef] [PubMed]
- Hultén, E.; Fries, M. Atlas of North European Vascular Plants North of the Tropic of Cancer; Maps 1–996; Koeltz Scientific Books: Königstein, Germany, 1986. [Google Scholar]
- Villellas, J.; Ehrlén, J.; Olesen, J.M.; Braza, R.; García, M.B. Plant performance in central and northern peripheral populations of the widespread Plantago coronopus. Ecography 2013, 36, 36–145. [Google Scholar] [CrossRef] [Green Version]
- Koyro, H.W. Effect on salinity on growth, photosynthesis, water relations and solute composition of the potential cash crop halophyte Plantago coronopus (L.). Environ. Exp. Bot. 2006, 56, 136–146. [Google Scholar] [CrossRef]
- Ceccanti, C.; Landi, M.; Incrocci, L.; Pardossi, A.; Venturi, F.; Taglieri, I.; Ferroni, G.; Guidi, L. Comparison of three domestications and wild-harvested plants for nutraceutical properties and sensory profiles in five wild edible herbs: Is domestication possible? Foods 2020, 9, 1065. [Google Scholar] [CrossRef]
- Boestfleisch, C.; Wagenseil, N.B.; Buhmann, A.K.; Seal, C.E.; Wade, E.M.; Muscolo, A.; Papenbrock, J. Manipulating the an-tioxidant capacity of halophytes to increase their cultural and economic value through saline cultivation. AoB Plants 2014, 6, plu046. [Google Scholar] [CrossRef]
- Höpke, J.; Mucina, L.; Albach, D.C. Phylogenetic and morphometric analysis of Plantago section Coronopus (Plantaginaceae). Taxon 2019, 68, 315–339. [Google Scholar] [CrossRef]
- Vicente, O.; Boscaiu, M.; Naranjo, M.A.; Estrelles, E.; Bellés, J.M.; Soriano, P. Responses to salt stress in the halophyte Plantago crassifolia (Plantaginaceae). J. Arid Environ. 2004, 58, 463–481. [Google Scholar] [CrossRef]
- El Shaer, H.M.; Attia-Ismail, S.A. Halophytic and salt-tolerant feedstuffs in the Mediterranean Basin and Arab Region: An overview. In Halophytic and Salt-Tolerant Feedstuffs: Impacts on Nutrition, Physiology and Reproduction of Livestock; El Shaer, H.M., Squires, V.R., Eds.; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Rønsted, N.; Chase, M.W.; Albaxh, D.C.; Bello, M.A. Phylogenetic relationships within Plantago (Plantaginaceae): Evidence from nuclear ribosomal ITS and plastid trnL-F sequence data. Bot. J. Linn. Soc. 2002, 139, 323–338. [Google Scholar] [CrossRef]
- Pedról, J.; Plantago, L. Flora ibérica 13 Plantaginaceae-Scrophulariaceae; Benedí, C., Rico, E., Eds.; Real Jardín Botánico, CSIC: Madrid, Spain, 2009; Volume 13, pp. 4–38. [Google Scholar]
- Meyers, S.C.; Liston, A. The biogeography of Plantago ovata Forssk. (Plantaginaceae). Int. J. Plant Sci. 2008, 169, 954–962. [Google Scholar] [CrossRef] [Green Version]
- Shryock, D.F.; Washburn, L.K.; De Falco, L.A.; Esque, T.C. Genetic and habitat data for Plantago ovata in the Mojave Desert: U. S. Geological Survey data release 2020. [Google Scholar] [CrossRef]
- Talukder, P.; Talapatra, S.; Ghoshal, N.; Raychaudhuri, S. Antioxidant activity and high-performance liquid chromatographic analysis of phenolic compounds during in vitro callus culture of Plantago ovata Forsk. and effect of exogenous additives on accumulation of phenolic compounds. J. Sci. Food Agric 2016, 96, 232–244. [Google Scholar] [CrossRef] [PubMed]
- Izadi-Darbandi, E.; Mehdikhani, H. Salinity effect on some of the morphophysiological traits of three Plantago species (Plantago spp.). Sci. Hortic. 2018, 236, 43–51. [Google Scholar] [CrossRef]
- Rubinigg, M.; Wenisch, J.; Elzenga, J.T.M.; Stulen, I. NaCl salinity affects lateral root development in Plantago martitima. Funct. Plant Biol. 2004, 31, 775–780. [Google Scholar] [CrossRef]
- Al Hassan, M.; Pacurar, A.; Gaspar, A.; Vicente, O.; Boscaiu, M. Growth and reproductive success under saline conditions of three Plantago species with different levels of stress tolerance. Not. Bot. Horti Agrobot. Cluj Napoca 2014, 42, 180–186. [Google Scholar] [CrossRef]
- Orsini, F.; D'Urzo, M.P.; Inan, G.; Serra, S.; Oh, D.H.; Mickelbart, M.V.; Consiglio, F.; Li, X.; Jeong, J.C.; Yun, D.J.; et al. A comparative study of salt tolerance parameters in 11 wild relatives of Arabidopsis thaliana. J. Exp. Bot. 2010, 61, 3787–3798. [Google Scholar] [CrossRef]
- Cicevan, R.; Al Hassan, M.; Sestras, A.F.; Prohens, J.; Vicente, O.; Sestras, R.E.; Boscaiu, M. Screening for drought tolerance in cultivars of the ornamental genus Tagetes (Asteraceae). PeerJ 2016, 4, e2133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todea, I.M.; González-Orenga, S.; Plazas, M.; Sestras, A.F.; Prohens, T.J.; Vicente, O.; Sestras, R.E. Screening for salt and water stress tolerance in fir (Abies alba) populations. Not. Bot. Horti Agrobot. Cluj Napoca 2019, 47, 1063–1072. [Google Scholar] [CrossRef] [Green Version]
- Kazal, V.; Bhushan, B.; Kumari Narval, R.; Kumar, S.; Pal, A. Evaluation of salt tolerance in different isabgol (Plantago ovata Forsk.) genotypes under differential salt stress imposition. Indian J. Agric. Biochem. 2017, 30, 41–49. [Google Scholar] [CrossRef]
- Khaliq, R.; Zahoor, M.; Zafar, Z.U.; Athar, H.R. Growth responses of Plantago ovata L. to varying levels of NaCl. Iran. J. Plant Physiol. 2011, 1, 157–167. [Google Scholar]
- Golkar, P.; Amooshahi, F.; Arzani, A. The effects of salt stress on physio-biochemical traits, total phenolic and mucilage content of Plantago ovata Forsk under in vitro conditions. J. Appl. Bot. Food Qual. 2017, 90, 224–231. [Google Scholar]
- Neumann, P.M. Inhibition of root growth by salinity stress: Toxicity or an adaptive biophysical response. In Structure and Function of Roots; Baluska, F., Ciamporova, M., Gasparikova, O., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1995; pp. 299–304. [Google Scholar]
- Chelli-Chaabouni, A.; Hammami, M.; Gouia, M.; Gargouri, K.; Gargouri, R.; Drira, N. Effect of Salt Stress on Pistacia atlantica Rootstock Seedlings in Nursery Conditions. In XIV GREMPA Meeting on Pistachios and Almonds; Zakynthinos, G., Ed.; CIHEAM/FAO/AUA/TEI Kalamatas/NAGREF: Zaragoza, Spain, 2010; pp. 135–140. [Google Scholar]
- Dubey, S.; Bhargava, A.; Fuentes, F.; Shukla, S.; Srivastava, S. Effect of salinity stress on yield and quality parameters in flax (Linum usitatissimum L.). Not. Bot. Horti Agrobot. Cluj Napoca 2020, 48, 954–966. [Google Scholar] [CrossRef]
- Rahneshan, Z.; Nasibi, F.; Moghadam, A.A. Effects of salinity stress on some growth, physiological, biochemical parameters and nutrients in two pistachio (Pistacia vera L.) rootstocks. J. Plant Interact. 2018, 13, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Stepien, P.; Johnson, G.N. Contrasting responses of photosynthesis to salt-stress in glycophyte Arabidopsis and halophyte Thellungiella: Role of the plastid terminal oxidase as an alternative electron sink. Plant Physiol. 2009, 149, 1154–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozminska, A.; Al Hassan, M.; Hanus-Fajerska, E.; Naranjo, M.A.; Bosciau, M.; Vicente, O. Comparative analysis of water deficit and salt tolerance mechanisms in Silene. South Afr. J. Bot. 2018, 117, 193–206. [Google Scholar] [CrossRef]
- Ashraf, M. Some important physiological selection criteria for salt tolerance in plants. Flora 2004, 199, 361–376. [Google Scholar] [CrossRef]
- Flexas, J.; Medrano, H. Drought inhibition of photosynthesis in C3 plants: Stomatal and non-stomatal limitations revised. Ann. Bot. 2002, 89, 183–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munné Bosch, S.; Allegre, I. Die and let live: Leaf senescence contributes to plant survival under drought stress. Funct. Plant Biol. 2004, 31, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef] [PubMed]
- Raven, J.A. Regulation of pH and generation of osmolarity in vascular plants: A cost-benefit analysis in relation to efficiency of use of energy, nitrogen and water. New Phytol. 1985, 101, 25–77. [Google Scholar] [CrossRef]
- Erdei, L.; Kuiper, P.J.C. The effect of salinity on growth, cation content, Na+-Uptake and translocation in salt-sensitive and salt-tolerant Plantago species. Physiol. Plant. 1979, 47, 95–99. [Google Scholar] [CrossRef]
- Königshofer, H. Changes in ion composition and hexitol content of different Plantago species under the influence of salt stress. Plant Soil 1983, 72, 289–296. [Google Scholar] [CrossRef]
- Sleimi, N.; Guerfali, S.; Bankaji, I. Biochemical indicators of salt stress in Plantago maritima: Implications for environmental stress assessment. Ecol. Indic. 2015, 48, 570–577. [Google Scholar] [CrossRef]
- Marschner, H. Ion uptake mechanisms of individual cells and roots: Short-distance transport. In Mineral Nutrition of Higher Plants, 2nd ed.; Academic Press: London, UK, 1995; pp. 6–78. [Google Scholar] [CrossRef]
- Greenway, H.; Munns, R. Mechanisms of salt tolerance in nonhalophytes. Ann. Rev. Plant Physiol. 1980, 31, 149–190. [Google Scholar] [CrossRef]
- Schiop, S.T.; Al Hassan, M.; Sestras, A.F.; Boscaiu, M.; Sestras, R.E.; Vicente, O. Identification of salt stress biomarkers in Romanian Carpathian populations of Picea abies (L.) Karst. PLoS ONE 2015, 10, e0135419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arteaga, S.; Al Hassan, M.; Chaminda Bandara, W.M.; Yabor, L.; Llinares, J.V.; Boscaiu, M.; Vicente, O. Screening for salt tolerance in four local varieties of Phaseolus lunatus from Spain. Agriculture 2018, 8, 201. [Google Scholar] [CrossRef] [Green Version]
- Borsai, O.; Al Hassan, M.; Negrușier, C.; Raigón, M.D.; Boscaiu, M.; Sestraș, R.E.; Vicente, O. Responses to salt stress in Portulaca: Insight into its tolerance mechanisms. Plants 2020, 9, 1660. [Google Scholar] [CrossRef] [PubMed]
- Shabala, S.; Cuin, T.A. Potassium transport and plant salt tolerance. Physiol. Plant. 2008, 133, 651–669. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, I.; Larher, F.; Stewart, G.R. Sorbitol, a compatible osmotic solute in Plantago maritima. New Phytol. 1979, 82, 671–678. [Google Scholar] [CrossRef]
- Tipirdamaz, R.; Gagneul, D.; Duhazé, C.; Aïnouche, A.; Monnier, C.; Özkum, D.; Larher, F. Clustering of halophytes from an inland salt marsh in Turkey according to their ability to accumulate sodium and nitrogenous osmolytes. Environ. Exp. Bot. 2006, 57, 139–153. [Google Scholar] [CrossRef]
- Pardo-Domènech, L.; Tifrea, A.; Grigore, M.N.; Boscaiu, M.; Vicente, O. Proline and glycine betaine accumulation in two succulent halophytes under natural and experimental conditions. Plant Biosyst 2016, 150, 904–915. [Google Scholar] [CrossRef]
- Kishor, P.B.K.; Sangam, S.; Amrutha, R.N.; Laxmi, P.S.; Naidu, K.R.; Rao, K.R.S.S.; Rao, S.; Reddy, K.J.; Theriappan, P.; Sreenivasulu, N. Regulation of proline biosynthesis, degradation, uptake and transport in higher plants: Its implications in plant growth and abiotic stress tolerance. Curr. Sci. 2005, 88, 424–438. [Google Scholar]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Iqbal, N.; Umar, S.; Khan, N.A.; Khan, M.I.R. A new perspective of phytohormones in salinity tolerance regulation of proline metabolism. Environ. Exp. Bot. 2014, 100, 34–42. [Google Scholar] [CrossRef]
- Liang, X.; Zhang, L.; Natarajan, S.K.; Becker, D.F. Proline mechanisms of stress survival. Antioxid. Redox Signal. 2013, 19, 998–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karimi, N.; Modarresi, M.; Mollaei, S.E. Enhancement of compatible solute and secondary metabolites production in Plantago ovata Forsk. by salinity stress. J. Med. Plant Res. 2012, 6, 3495–3500. [Google Scholar]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plant: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef] [PubMed]
- Gil, R.; Boscaiu, M.; Lull, C.; Bautista, I.; Lid, N.A.; Vicente, O. Are soluble carbohydrates ecologically relevant for salt tolerance in halophytes? Funct. Plant. Biol. 2013, 40, 805–818. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–379. [Google Scholar] [CrossRef] [Green Version]
- Del Rio, D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [CrossRef]
- Sekmen, A.H.; Türkan, I.; Takio, S. Differential responses of antioxidative enzymes and lipid peroxidation to salt stress in salt-tolerant Plantago maritima and salt-sensitive Plantago media. Physiol. Plant. 2007, 131, 399–411. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Vicente, O.; Boscaiu, M. Flavonoids: Antioxidant compounds for plant defence... and for a healthy human diet. Not. Bot. Horti Agrobot. Cluj Napoca 2018, 46, 14–21. [Google Scholar] [CrossRef] [Green Version]
- Türkan, I.; Demiral, T.J.; Sekmen, A.H. The regulation of antioxidant enzymes in two Plantago species differing in salinity tolerance under combination of waterlogging and salinity. Funct. Plant Biol. 2012, 40484–40493. [Google Scholar] [CrossRef] [PubMed]
- Jurišić Grubešić, R.; Srečnik, G.; Kremer, D.; Vuković Rodríguez, J.; Nikolić, T.; Vladimir-Knežević, S. Simultaneous RP-HPLC-DAD separation and determination of flavonoids and phenolic acids in Plantago, L. species. Chem. Biodiver. 2013, 10, 1305–1316. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.J.; Gangadhar, K.N.; Vizetto-Duarte, C.; Wubshet, S.G.; Nyberg, N.T.; Barreira, L.; Varela, J.; Custódio, L. Maritime halophyte species from southern Portugal as sources of bioactive molecules. Mar. Drugs 2014, 12, 2228. [Google Scholar] [CrossRef] [Green Version]
- Pereira, C.G.; Custódio, L.; Rodrigues, M.J.; Neng, N.R.; Nogueira, J.M.F.; Carlier, J.; Costa, M.C.; Varela, J.; Barreira, L. Profiling of antioxidant potential and phytoconstituents of Plantago coronopus. Braz. J. Biol. 2017, 77, 3. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K.; Wellburn, A.R. Determination of total carotenoids and chlorophyll a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Weimberg, R. Solute adjustments in leaves of two species of wheat at two different stages of growth in response to salinity. Physiol. Plant. 1987, 70, 381–388. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Hodges, D.M.; Delong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Taulavuori, E.; Hellström, E.K.; Taulavuori, K.; Laine, K. Comparison of two methods used to analyse lipid peroxidation from Vaccinium myrtillus L. during snow removal, re-acclimation and cold acclimation. J. Exp. Bot. 2021, 52, 2375–2380. [Google Scholar] [CrossRef] [Green Version]
- Loreto, F.; Velikova, V. Isoprene produced by leaves protects the photosynthetic apparatus against ozone damage, quenches ozone products and reduces lipid peroxidation of cellular membranes. Plant Physiol. 2001, 127, 1781–1787. [Google Scholar] [CrossRef]
- Blainski, A.; Lopes, G.C.; Palazzodemello, J.C. Application and analysis of the Folin-Ciocalteu method for the determination of the total phenolic content from Limonium brasiliense L. Molecules 2013, 18, 6852–6865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Utz, H.F. Plabstat: A Computer Program for Statistical Analysis of Plant Breeding Experiments; Version 3A; Universitat Hohenheim: Stuttgart, Germany, 2001. [Google Scholar]
- Hammer, O.; Harper, D.A.T.; Rayan, P.D. Past: Palaeontogical statistics software package for education and data analysis. Palaeontol. Electron. 2011, 4, 9. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | S | T | S × T |
|---|---|---|---|
| ECs | 297.04 * | 129.78 * | 62.07 * |
| RL | 205.23 * | 34.43 * | 70.59 * |
| SL | 35.99 * | 53.34 * | 8.65 * |
| FW (%) | 10.29 * | 66.62 * | 1.56 |
| RWC | 91.76 * | 49.30 * | 17.78 * |
| LWC | 142.33 * | 87.91 * | 16.21 * |
| Chl a | 17.68 * | 45.54 * | 8.04 * |
| Chl b | 11.69 * | 29.51 * | 2.68 * |
| Caro | 12.59 * | 48.68 * | 3.30 * |
| Na+ R | 239.91 * | 26.67 * | 22.49 * |
| Na+ L | 235.82 * | 62.31 * | 49.70 * |
| Cl− R | 23.76 * | 10.69 * | 7.04 * |
| Cl− L | 77.36 * | 23.50 * | 29.56 * |
| K+ R | 40.97 * | 21.01 * | 3.45 * |
| K+ L | 63.60 * | 0.90 | 9.12 * |
| MDA | 55.67 * | 102.62 * | 29.84 * |
| H2O2 | 102.17 * | 11.32 * | 22.67 * |
| TSS | 7.75 * | 58.23 * | 20.62 * |
| Pro | 192.94 * | 63.45 * | 33.07 * |
| TPC | 40.68 * | 10.35 * | 24.94 * |
| TF | 93.51 * | 18.10 * | 51.53 * |
| Control | 200 mM | 400 mM | 600 mM | 800 mM NaCl | |
|---|---|---|---|---|---|
| P. crassifolia | 0.48 ± 0.02 a | 3.56 ± 0.18 b | 5.46 ± 0.22 c | 7.94 ± 0.50 d | 9.45 ± 0.30 e |
| P. coronopus | 0.45 ± 0.02 a | 1.20 ± 0.09 b | 5.66 ± 0.50 c | 7.60 ± 0.10 d | 9.57 ± 0.20 e |
| P. ovata | 0.45 ± 0.02 a | 1.51 ± 0.05 b | 5.63 ± 0.14 c | 8.51 ± 0.40 d | n.d. |
| P. afra | 0.49 ± 0.05 a | 2.15 ± 0.15 b | n.d. | n.d. | n.d. |
| Species | Subgenus | Location | Latitude | Longitude | Bioclimatic Zones |
|---|---|---|---|---|---|
| P. coronopus L. | Coronopus | Hergla/Sousse | 35°58′52.07″ | 10°31′38.14″ | Semi-arid inferior |
| P. crassifolia Forsk. | Coronopus | Djerba/Mednine | 33°49′52.96″ | 11°2′17.67″ | Arid inferior |
| P. ovata Forsk. | Psyllium | Tataouine | 32°55′8.63″ | 10°24′59.45″ | Saharian superior |
| P. afra L. | Psyllium | Bouargoub | 36°28′34.84″ | 10°36′46.35″ | Semi-arid superior |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ltaeif, H.B.; Sakhraoui, A.; González-Orenga, S.; Landa Faz, A.; Boscaiu, M.; Vicente, O.; Rouz, S. Responses to Salinity in Four Plantago Species from Tunisia. Plants 2021, 10, 1392. https://doi.org/10.3390/plants10071392
Ltaeif HB, Sakhraoui A, González-Orenga S, Landa Faz A, Boscaiu M, Vicente O, Rouz S. Responses to Salinity in Four Plantago Species from Tunisia. Plants. 2021; 10(7):1392. https://doi.org/10.3390/plants10071392
Chicago/Turabian StyleLtaeif, Hela Belhaj, Anis Sakhraoui, Sara González-Orenga, Anbu Landa Faz, Monica Boscaiu, Oscar Vicente, and Slim Rouz. 2021. "Responses to Salinity in Four Plantago Species from Tunisia" Plants 10, no. 7: 1392. https://doi.org/10.3390/plants10071392
APA StyleLtaeif, H. B., Sakhraoui, A., González-Orenga, S., Landa Faz, A., Boscaiu, M., Vicente, O., & Rouz, S. (2021). Responses to Salinity in Four Plantago Species from Tunisia. Plants, 10(7), 1392. https://doi.org/10.3390/plants10071392