
Gentianella lutescens subsp. carpatica J. Holub.: Shoot Propagation In Vitro and Effect of Sucrose and Elicitors on Xanthones Production

 , , and
, , and 
Abstract
:1. Introduction
2. Results and Discussion
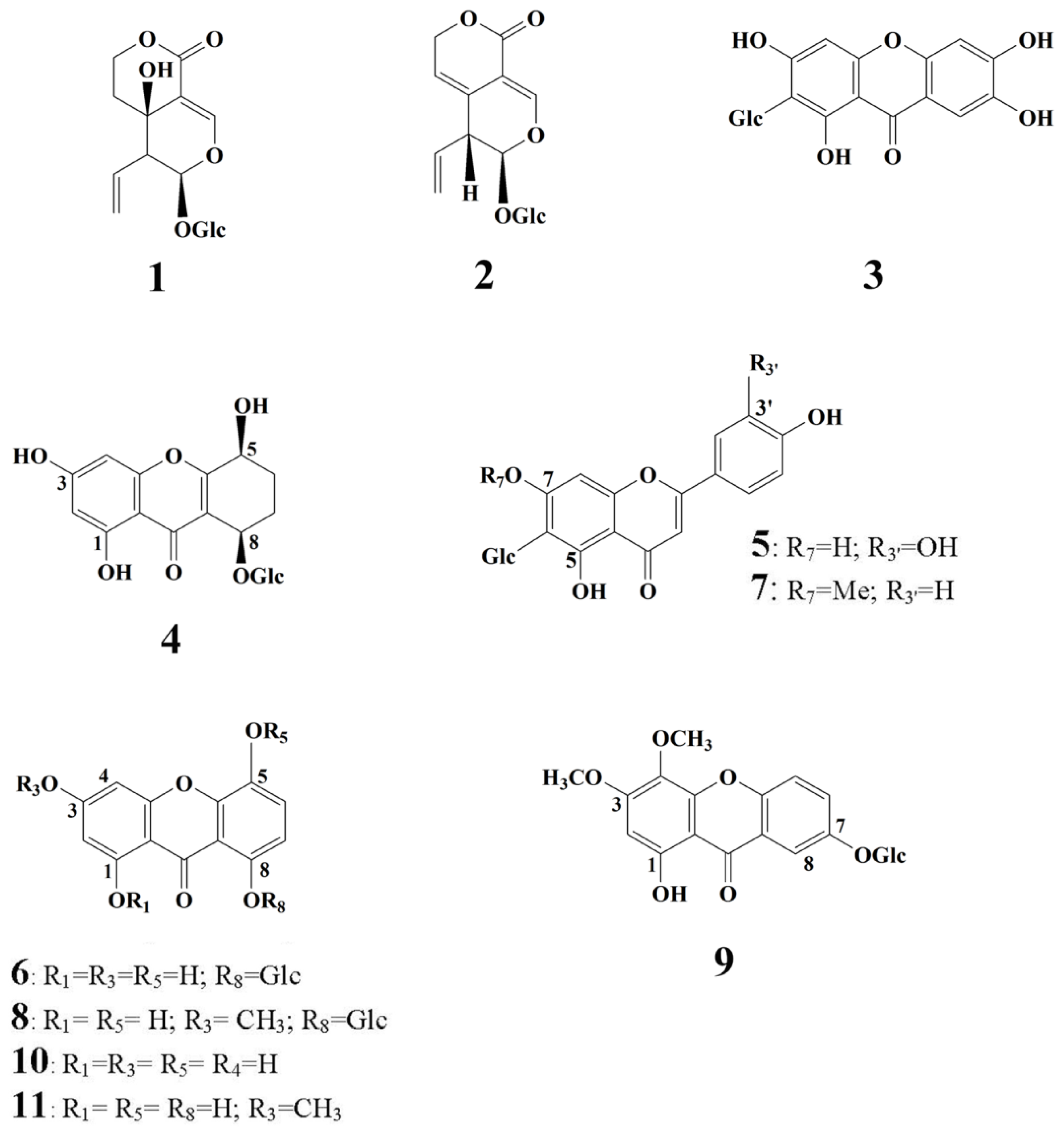
2.1. HPLC-DAD Analysis of Secondary Metabolites of Wild Grown G. lutescens Plants
2.2. In Vitro Shoot Propagation of G. lutescens
2.3. HPLC-DAD Analysis of Secondary Metabolites of G. lutescens Cultured In Vitro
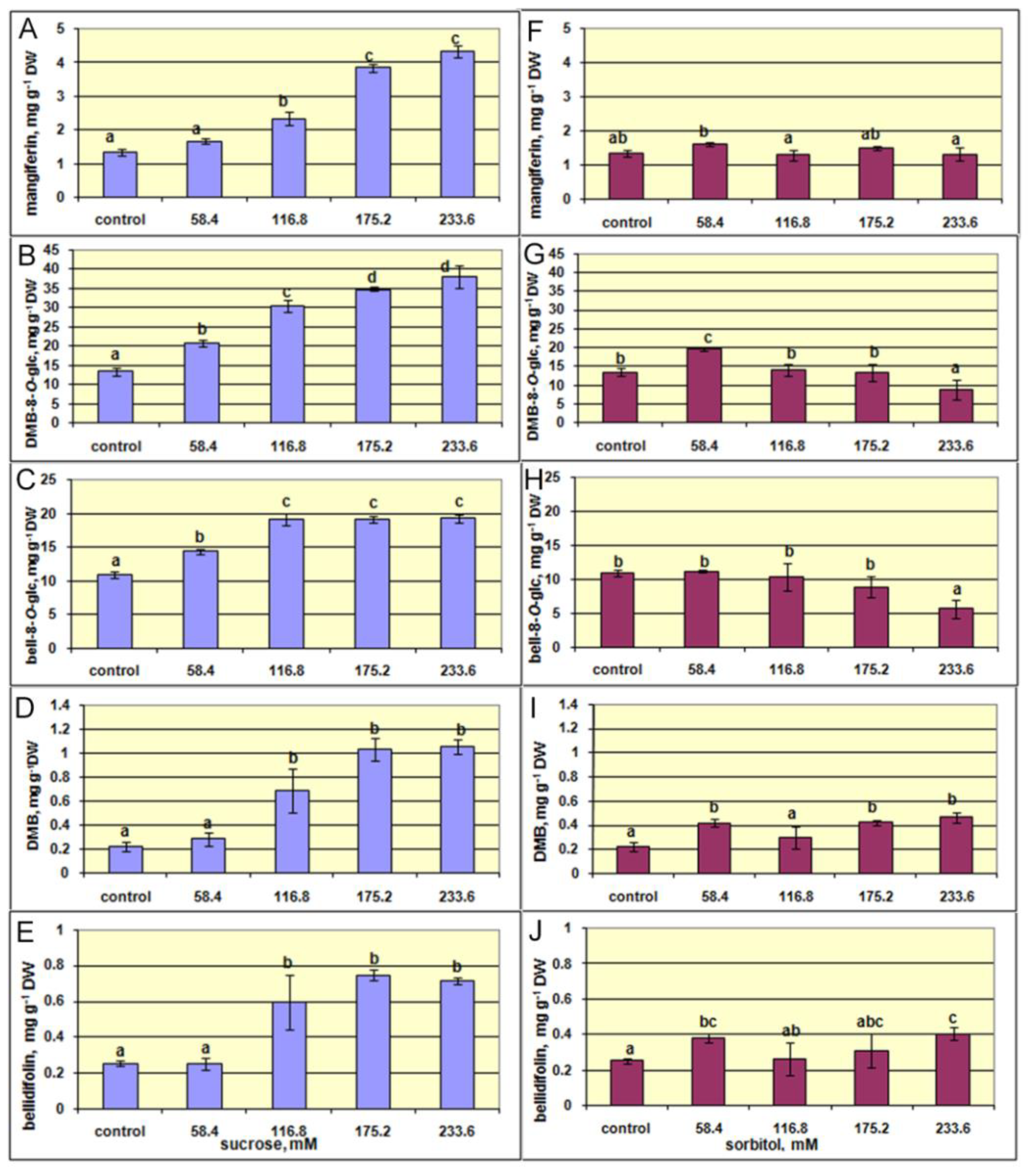
2.4. The Effect of Sucrose and Sorbitol on Shoot Growth and Xanthone Production
2.5. The Effect of Elicitors JA, MeJA and SA on Shoot Growth and Xanthone Production
3. Materials and Methods
3.1. Plant Material
3.2. In Vitro Seed Germination and Shoot Culture Initiation
3.3. Increase in Sucrose and Sorbitol Level
3.4. Elicitor Preparation and Application
3.5. Xanthone Extraction and Conditions for HPLC-DAD Analysis
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- von Hagen, K.B.; Kadereit, J.W. The phylogeny of Gentianella (Gentianaceae) and its colonization of the southern hemisphere as revealed by nuclear and chloroplast DNA sequence variation. Org. Divers. Evol. 2001, 1, 61–79. [Google Scholar] [CrossRef] [Green Version]
- Stevanović, V.; Jakovljević, K. Distribution, ecology, and some taxonomical notes of the genera Gentiana L. and Gentianella Moench (Gentianaceae) in the Balkans. In The Gentianaceae—Volume 1: Characterization and Ecology; Rybczyński, J., Davey, M., Mikula, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 169–200. [Google Scholar]
- Jovanović-Dunjić, R. Rod Gentianella Moench. In Flora SR Srbije; Josifović, M., Ed.; SANU: Beograd, Serbia, 1973; Volume 5, pp. 426–433. [Google Scholar]
- Lacaille-Dubois, M.; Galle, K.; Wagner, H. Secoiridoids and xanthones from Gentianella nitida. Planta Med. 1996, 62, 365–367. [Google Scholar] [CrossRef]
- Nadinic, E.; Gorzalczany, S.; Rojo, A.; van Baren, C.; Debenedetti, S.; Acevedo, C. Topical antiinflammatory activity of Gentianella achalensis. Fitoterapia 1999, 70, 166–171. [Google Scholar] [CrossRef]
- Lu, S.; Tanaka, N.; Kawazoe, K.; Murakami, K.; Damdinjav, D.; Dorjbal, E.; Kashiwada, Y. Tetrahydroxanthones from Mongolian medicinal plant Gentianella amarella ssp. acuta. J. Natur. Med. 2016, 70, 780–788. [Google Scholar] [CrossRef]
- Hostettmann-Kaldas, M.; Jacot-Guillarmod, A. Contribution to the phytochemistry of the genus Gentiana. Part XXIII. Xanthones and flavone C-glucosides of the genus Gentiana (subgenus Gentianella). Phytochemistry 1978, 17, 2083–2086. [Google Scholar] [CrossRef]
- Tovilovic-Kovacevic, G.; Zogovic, N.; Krstic-Milosevic, D. Secondary metabolites from endangered Gentiana, Gentianella, Centaurium, and Swertia species (Gentianaceae): Promising natural biotherapeutics. In Biodiversity and Biomedicine; Ozturk, M., Egamberdieva, D., Pešić, M., Eds.; Elsevier-Academic Press: London, UK, 2020; pp. 335–385. [Google Scholar]
- Tian, L.-Y.; Bai, X.; Chen, X.-H.; Fang, J.-B.; Liu, S.-H.; Chen, J.-C. Anti-diabetic effect of methylswertianin and bellidifolin from Swertia punicea Hemsl. and its potential mechanism. Phytomedicine 2010, 17, 533–539. [Google Scholar] [CrossRef]
- Zhao, Z.-Y.; Gao, Y.-Y.; Gao, L.; Zhang, M.; Wang, H.; Zhang, C.-H. Protective effects of bellidifolin in hypoxia-induced in pheochromocytoma cells (PC12) and underlying mechanisms. J. Toxicol. Envir. Health Part A 2017, 80, 1187–1192. [Google Scholar] [CrossRef]
- Cruz, M.I.; Cidade, H.; Pinto, M. Dual/multitargeted xanthone derivatives for Alzheimer’s disease: Where do we stand? Future Med. Chem. 2017, 9, 1611–1630. [Google Scholar] [CrossRef] [PubMed]
- Tutin, T.G. Gentianella Moench. In Flora Europea; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1972; Volume 3, pp. 63–67. [Google Scholar]
- Kirschnerová, L.; Kirschner, J. Changes in the distribution of Gentianella lutescens subsp. lutescens and subsp. carpatica in the Czech Republic. Zprávy Èes. Bot. Spoleè. Praha 2003, 38, 205–216. [Google Scholar]
- IUCN. IUCN Red List Categories and Criteria: Version 3.1, 2nd ed.; IUCN: Gland, Switzerland; Cambridge, UK, 2012; 32p. [Google Scholar]
- Sharma, V.; Bhardwaj, U.; Sharma, S.; Sharma, S. Medicinal Plants: Need for Sustainable Exploitation (With Special reference to Himachal Pradesh). J. Pharm. Res. 2012, 5, 4313–4317. [Google Scholar]
- Dias, M.I.; Sousa, M.J.; Alves, R.C.; Ferreira, I.C.F.R. Exploring plant tissue culture to improve the production of phenolic compounds: A review. Ind. Crops Prod. 2016, 82, 9–22. [Google Scholar] [CrossRef] [Green Version]
- Chandran, H.; Meena, M.; Barupal, T.; Sharma, K. Plant tissue culture as a perpetual source for production of industrially important bioactive compounds. Biotechn. Rep. 2020, 26, e00450. [Google Scholar] [CrossRef]
- Vinterhalter, B.; Janković, T.; Šavikin, K.; Nikolić, R.; Vinterhalter, D. Propagation and xanthone content of Gentianella austriaca shoot cultures. Plant. Cell Tiss. Organ. Cult. 2008, 94, 329–335. [Google Scholar] [CrossRef]
- Janković, T.; Vinterhalter, B.; Krstić-Milošević, D.; Nikolić, R.; Vinterhalter, D.; Milosavljević, S. Xanthone compounds in shoot cultures of Gentianella bulgarica. Acta Physiol. Plant. 2011, 33, 1515–1520. [Google Scholar] [CrossRef]
- Krstić-Milošević, D.; Vinterhalter, B.; Janković, T.; Vinterhalter, D. Biotechnology and Phytochemistry of Gentianella Species from the Central Regions of the Balkan Peninsula. In The Gentianaceae—Biotechnology and Applications; Rybczyński, J., Davey, M., Mikuła, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; Volume 2, pp. 93–112. [Google Scholar]
- Huo, L.-Y.; Zheng, X.-J. Studies on tissue culture of Gentianella albifpra. Chin. Pharm. J. 2002, 37, 415–418. [Google Scholar]
- Solorzano Acosta, A.; Ruiz Reyez, G.; Venegas, E. Preliminary identification secondry metabolites produced in induced by callus “Corpus way” Gentianella bicolor (Wedd.) Fabris exJ.s. Pringle. Rev. Sci. 2014, 6, 137–142. [Google Scholar]
- Moraes, R.M.; Cerdeira, A.L.; Lourenço, M.V. Using micropropagation to develop medicinal plants into crops. Molecules 2021, 26, 1752. [Google Scholar] [CrossRef] [PubMed]
- Franklin, G.; Conceicao, L.F.R.; Kombrink, E.; Dias, A.C.P. Xanthone biosynthesis in Hypericum perforatum cells provides antioxidant and antimicrobial protection upon biotic stress. Phytochemistry 2009, 70, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Conceição, L.F.R.; Ferreres, F.; Tavares, R.M.; Dias, A.C.P. Induction of Hypericum perforatum L. cells by Colletotrichum gloeosporioides elicitation. Phytochemistry 2006, 67, 140–155. [Google Scholar] [CrossRef]
- Wang, J.; Qian, J.; Yao, L.; Lu, Y. Enhanced production of flavonoids by methyl jasmonate elicitation in cell suspension culture of Hypericum perforatum. Bioresour. Bioprocess. 2015, 2, 5–13. [Google Scholar] [CrossRef] [Green Version]
- Gadzovska, S.; Maury, S.; Delaunay, A.; Spasenoski, M.; Hagege, D.; Courtois, D.; Joseph, C. The influence of salicylic acid elicitation of shoots, callus, and cell suspension cultures on production of naphtodianthrones and phenylpropanoids in Hypericum perforatum L. Plant. Cell Tiss. Org. Cult. 2013, 113, 25–39. [Google Scholar] [CrossRef]
- Krstić-Milošević, D.; Janković, T.; Vinterhalter, B.; Menković, N.; Aljančić, I.; Vinterhalter, D. Influence of carbohydrate source on xanthone content in root cultures of Gentiana dinarica Beck. Plant. Growth Regul. 2013, 71, 147–155. [Google Scholar] [CrossRef]
- Sahitya, U.L.; Krishna, M.S.R.; Deepthi, R.S. Biochemical and physiological changes induced by water stress in hot pepper (Capsicum annum L.) genotypes. Plant. Cell Biotech. Mol. Biol. 2018, 19, 179–195. [Google Scholar]
- Jensen, S.R.; Schripsema, J. Chemotaxonomy and pharmacology of Gentianaceae. In Gentianaceae: Systematics and Natural History; Struwe, L., Albert, V.A., Eds.; Cambridge University Press: Cambridge, UK, 2002; p. 573. [Google Scholar]
- Janković, T.; Krstić, D.; Aljančić, I.; Šavikin-Fodulović, K.; Menković, N.; Vajs, V.; Milosavljević, S. Xanthones and C-glucosides from the aerial parts of four species of Gentianella from Serbia and Montenegro. Biol. Biochem. Syst. Ecol. 2005, 33, 729–735. [Google Scholar] [CrossRef]
- Murthy, H.N.; Dandin, V.S.; Paek, K.-Y. Tools for biotechnological production of useful phytochemicals from adventitious root cultures. Phytochem. Rev. 2016, 15, 129–145. [Google Scholar] [CrossRef]
- Bakhtiar, Z.; Mirjalili, M.H.; Sonboli, A.; Moridi Farimani, M.; Ayyari, M. In vitro propagation, genetic and phytochemical assessment of Thymus persicus—A medicinally important source of pentacyclic triterpenoids. Biologia 2014, 69, 594–603. [Google Scholar] [CrossRef]
- Sirikonda, A.; Jogam, P.; Ellendula, R.; Kudikala, H.; Mood, K.; Allini, V.R. In vitro micropropagation and genetic fidelity assesment in Flemingia macrophylla (Willd.) Merr: An ethnomedicinal plant. Vegetos 2020, 33, 286–295. [Google Scholar] [CrossRef]
- Devi, T.R.; Dasgupta, M.; Sahoo, M.R.; Kole, P.C.; Prakash, N. High efficient de novo rootto-shoot organogenesis in Citrus jambhiri Lush.: Gene expression, genetic stability and virus indexing. PLoS ONE 2021, 16, e0246971. [Google Scholar] [CrossRef] [PubMed]
- Gu, R.H.; Hong, L.Y.; Long, C.L. The ways of producing secondary metabolites via plant cell culture. Plant. Physiol. J. 2016, 49, 869–881. [Google Scholar]
- Dević, M.; Momčilović, I.; Kristić, D.; Maksimović, V.; Konjević, R. In vitro multiplication of willow gentian (Gentiana asclepiadea L.) and the production of gentiopicrin and mangiferin. Phyton 2006, 46, 45–54. [Google Scholar]
- Rolland, F.; Moore, B.; Sheen, J. Sugar sensing and signaling in plants. Plant. Cell 2002, 14, 185–205. [Google Scholar] [CrossRef] [Green Version]
- De Paiva Neto, V.B.; Otoni, W.C. Carbon sources and their osmotic potential in plant tissue culture: Does it matter? Sci. Hortic. 2003, 97, 193–202. [Google Scholar] [CrossRef]
- Grattapaglia, D.; Machado, M.A. Micropropagação. In Cultura de Tecidos e Transformação Genética de Plantas; Torres, A.C., Caldas, L.S., Buso, J.A., Eds.; Embrapa-SPI/Embrapa-CNPH: Brasília, Brazil, 1998; Volume 1, pp. 43–76. [Google Scholar]
- Hazubska-Przybył, T.; Kalemba, E.; Ratajczak, E.; Bojarczuk, K. Effects of abscisic acid and an osmoticum on the maturation, starch accumulation and germination of Picea spp. somatic embryos. Acta Phy. Plant. 2016, 38, 59. [Google Scholar] [CrossRef] [Green Version]
- Siwach, P.; Grover, K.; Gill, A.R. The influence of plant growth regulators, explant nature and sucrose concentration on in vitro callus growth of Thevetia peruviana Schum. Asian J. Biotechnol. 2011, 3, 280–292. [Google Scholar] [CrossRef] [Green Version]
- Takeno, K. Stress-induced flowering: The third category of flowering response. J. Exp. Bot. 2016, 67, 4925–4934. [Google Scholar] [CrossRef] [Green Version]
- Al-Khateeb, A.A. Regulation of in vitro bud formation of date palm (Phoenix dactylifera L.) cv. Khanezi by different carbon sources. Bioresour. Technol. 2008, 99, 6550–6555. [Google Scholar] [CrossRef]
- Mohamed, M.A.H.; Alsadon, A.A. Influence of ventilation and sucrose on growth and leaf anatomy of micropropagated potato plantlets. Sci. Hort. 2010, 123, 295–300. [Google Scholar] [CrossRef]
- Martins, J.P.R.; Verdoodt, V.; Pasqual, M.; De Proft, M. Impacts of photoautotrophic and photomixotrophic conditions on in vitro propagated Billbergia zebrina (Bromeliaceae). Plant. Cell Tiss. Org. Cult. 2015, 123, 121–132. [Google Scholar] [CrossRef]
- Sivanesan, L.; Hwang, S.L.; Jeong, B.R. Influence of plant growth regulators on axillary shoot multiplication and iron source on growth of Scrophularia takesimensis Nakal—A rare endemic medicinal plant. Afr. J. Biotechnol. 2008, 7, 4484–4490. [Google Scholar]
- Babb, V.M.; Haigler, C.H. Sucrose phosphate synthase activity rises in correlation with high-rate cellulose synthesis in three heterotrophic systems. Plant. Physiol. 2001, 127, 1234–1242. [Google Scholar] [CrossRef]
- Gurel, S.; Gulsen, Y. The effects of different sucrose, agar and pH levels on in vitro shoot production of almond (Amygdalus communis L.). Turkish J. Bot. 1998, 22, 363–373. [Google Scholar]
- Sotiropoulos, T.E.; Molassiotis, A.N.; Mouhtaridou, G.I.; Papadakis, I.; Dimassi, K.N.; Therios, I.N.; Diamantidis, G. Sucrose and sorbitol effects on shoot growth and proliferation in vitro, nutritional status and peroxidase and catalaze isoenzymes of M 9 and MM 106 apple (Malus domestica Borkh.) rootstocks. Eur. J. Hortic. Sci. 2006, 71, 114–119. [Google Scholar]
- Ahmad, T.; Abbasi, N.A.; Hafiz, I.A.; Ali, A. Comparison of sucrose and sorbitol as main carbon energy sources in micropropagation of peach rootstock GF-677. Pak. J. Bot. 2007, 39, 1269–1275. [Google Scholar]
- Salerno, G.L.; Curatti, L. Origin of sucrose metabolism in higher plants: When, how and why? Trends Plant. Sci. 2003, 8, 63–69. [Google Scholar] [CrossRef]
- Janković, T.; Krstić, D.; Šavikin-Fodulović, K.; Menković, N.; Grubišić, D. Xanthone compounds of Centaurium erythraea grown in nature and cultured in vitro. Pharm. Pharmacol. Lett. 2000, 10, 23–25. [Google Scholar]
- Cui, X.-H.; Murthy, H.N.; Wu, C.-H.; Peak, K.-Y. Sucrose-induced osmotic stress affects biomass, metabolite, and antioxidant levels in root suspension cultures of Hypericum perforatum L. Plant. Cell Tiss. Org. Cult. 2010, 103, 7–14. [Google Scholar] [CrossRef]
- Zhao, J.; Davis, L.C.; Verpoorte, R. Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol. Adv. 2005, 23, 283–333. [Google Scholar] [CrossRef]
- Namedo, A.G. Plant cell elicitation for production of secondary metabolites: A review. Pharmacogn. Rev. 2007, 1, 69–79. [Google Scholar]
- Wungsintaweekul, J.; Choo-malee, J.; Charoonratana, T.; Niwat, N. Methyl jasmonate and yeast extract stimulate mitragynine production in Mitragyna speciosa (Roxb.) Korth shoot culture. Biotechnol. Lett. 2012, 34, 1945–1950. [Google Scholar] [CrossRef]
- Ziaratnia, S.; Kunert, K.; Lall, N. Elicitation of 7-methyljuglone in Drosera capensis. S. Afr. J. Bot. 2009, 75, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Yadav, S.; Srivastava, A.; Shrivastava, N. Methyl jasmonate mediates upregulation of bacoside A production in shoot cultures of Bacopa monnieri. Biotechnol. Lett. 2013, 35, 1121–1125. [Google Scholar] [CrossRef]
- Munish, S.; Ashok, A.; Rajinder, G.; Sharada, M. Enhanced bacoside production in shoot cultures of Bacopa monnieri under the influence of abiotic elicitors. Nat. Prod. Res. 2015, 29, 745–749. [Google Scholar]
- Sivanandhan, G.; Rajesh, M.; Arun, M.; Jeyaraj, M.; Kapil Dev, G.; Arjunan, A.; Manickavasagam, M.; Muthuselvam, M.; Selvaraj, N.; Ganapathi, A. Effect of culture conditions, cytokinins, methyl jasmonate and salicylic acid on the biomass accumulation and production of withanolides in multiple shoot culture of Withania somnifera (L.) Dunal using liquid culture. Acta Physiol. Plant. 2013, 35, 715–728. [Google Scholar] [CrossRef]
- Kovácik, J.; Grúz, J.; Backor, M.; Strnad, M.; Repcák, M. Salicylic acid induced changes to growth and phenolic metabolism in Matricaria chamomilla plants. Plant. Cell Rep. 2009, 28, 135–143. [Google Scholar] [CrossRef]
- Khojasteh, A.; Mirjalili, M.H.; Palazon, J.; Eibl, R.; Cusido, R.M. Methyl jasmonate enhanced production of rosmarinic acid in cell cultures of Satureja khuzistanica in a bioreactor. Eng. Life Sci. 2016, 16, 740–749. [Google Scholar] [CrossRef]
- Lucioli, S.; Di Bari, C.; Nota, P.; Frattarelli, A.; Forni, C.; Caboni, E. Methyl jasmonate promotes anthocyanins’ production in Prunus salicina × Prunus persica in vitro shoot cultures. Plant. Biosyst. 2017, 151, 788–791. [Google Scholar] [CrossRef]
- Krstić-Milošević, D.; Janković, T.; Uzelac, B.; Vinterhalter, D.; Vinterhalter, B. Effect of elicitors on xanthone accumulation and biomass production in hairy root cultures of Gentiana dinarica. Plant. Cell Tiss. Org. Cult. 2017, 130, 631–664. [Google Scholar] [CrossRef]
- Chodisetti, B.; Rao, K.; Gandi, S.; Giri, A. Gymnemic acid enhancement in the suspension cultures of Gymnema sylvestre by using the signaling molecules-methyl jasmonate and salicylic acid. In Vitro Cell Dev. Biol. Plant 2015, 51, 88–92. [Google Scholar] [CrossRef]
- Piatczak, E.; Kuzma, L.; Wysokinska, H. The influence of methyl jasmonate and salicylic acid on secondary metabolite production in Rehmannia glutinosa libosch hairy root culture. Acta Biol. Cracov. Bot. 2016, 58, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Pavlik, M.; Vacek, J.; Klejdus, B.; Kuban, V. Hypericin and hyperforin production in St. John’s worth in vitro culture: Influence of succharose, polyethylene glycol, methyl jasmonate and Agrobacterium tumefaciens. J. Agric. Food Chem. 2007, 55, 6147–6153. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.-Y.; Yang, F.; Piao, X.-C.; Jin, M.-Y.; Tian, W.; Gao, Y.; Lian, M.-L. Promising strategy to efficiently improve the kinsenoside and polysaccharide production of rhizome cultures of Anoectochilus roxburghii (Wall.) Lindl. Ind. Crops Prod. 2018, 125, 269–275. [Google Scholar] [CrossRef]
- Patil, J.G.; Ahire, M.L.; Nitnaware, K.M.; Panda, S.; Bhatt, V.P.; Kavi Kishor, P.B.; Nikam, T.D. In vitro propagation and production of cardiotonic glycosides in shoot cultures of Digitalis purpurea L. by elicitation and precursor feeding. Appl. Microbiol. Biotechnol. 2013, 97, 2379–2393. [Google Scholar] [CrossRef]
- Tanatorn, S.; Nuttha, T.; Polkit, S.; Aphichart, K. Effect of methyl jasmonate on isoflavonoid accumulation and antioxidant enzymes in Pueraria mirifica cell suspension culture. J. Med. Plant. Res. 2014, 8, 401–407. [Google Scholar] [CrossRef] [Green Version]
- Wiktorowska, E.; Dlugosz, M.; Janiszowska, W. Significant enhancement of oleanolic acid accumulation by biotic elicitors in cell suspension cultures of Calendula officinalis L. Enzym Microb. Technol. 2010, 46, 14–20. [Google Scholar] [CrossRef]
- Chong, T.M.; Abdullah, M.A.; Lai, O.M.; Nor’Aini, F.M.; Lajis, N.H. Effective elicitation factors in Morinda elliptica cell suspension culture. Process. Biochem. 2005, 40, 3397–3405. [Google Scholar] [CrossRef]
- Coste, A.; Vlase, L.; Halmagyi, A.; Deliu, C.; Coldea, G. Effects of plant growth regulators and elicitors on production of secondary metabolites in shoot cultures of Hypericum hirsutum and Hypericum maculatum. Plant. Cell Tiss. Org. Cult. 2011, 106, 279–288. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F.A. Revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Linsmaier, E.M.; Skoog, F. Organic growth factor requirements of tobacco tissue cultures. Physiol. Plant. 1965, 18, 100–127. [Google Scholar] [CrossRef]
- Krstić-Milošević, D.; Banjac, N.; Janković, T.; Eler, K.; Vinterhalter, B. Gentiana clusii Perr.&Song.: Enhanced production of secondary metabolites by in vitro propagation. Plant. Phyisol. Biochem. 2020, 154, 735–744. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Line (BA 0.2 mg L−1) | No. of Explants | Multiplication Index ± SE | Length of Main Shoot (mm) ± SE | |
| line 1 | 80 | 3.03 ± 0.23 ab | 14.49 ± 0.65 a | |
| line 2 | 77 | 3.31 ± 0.19 ab | 17.40 ± 0.92 ab | |
| line 3 | 80 | 3.53 ± 0.23 b | 23.05 ± 1.67 c | |
| line 4 | 75 | 2.76 ± 0.25 a | 17.19 ± 1.22 ab | |
| line 5 | 80 | 3.41 ± 0.26 ab | 19.39 ± 1.07 b | |
| ANOVA Source of Variation | Df | Mean Square | F-Ratio | p-Value |
| Multiplication index | 4 | 7.48476 | 1.73 | 0.1428 |
| Length of main shoot | 4 | 804.293 | 7.67 | 0.0000 |
| BA (mg L−1) | No. of Expl. | Multiplication Index ± SE | Length of Main Shoot (mm) ± SE | Vitrification | |
| No. | % | ||||
| 0 | 85 | 2.89 ± 0.16 a | 15.02 ± 0.66 | 1 | 0.8 |
| 0.05 | 100 | 3.16 ± 0.17 a | 15.85 ± 0.50 e | 1 | 1.0 |
| 0.1 | 128 | 3.20 ± 0.14 a | 13.66 ± 0.46 cd | 7 | 5.47 |
| 0.2 | 90 | 3.92 ± 0.25 b | 15.48 ± 0.64 cd | 10 | 7.75 |
| 0.5 | 129 | 3.22 ± 0.17 a | 13.09 ± 0.48 bc | 24 | 18.6 |
| 1.0 | 128 | 2.88 ± 0.15 a | 12.09 ± 0.38 b | 20 | 15.6 |
| 2.0 | 80 | 2.94 ± 0.20 a | 10.46 ± 0.47 a | 14 | 17.5 |
| ANOVA Source of Variation | Df | Mean Square | F-Ratio | p-Value | |
| Multiplication index | 6 | 12.2028 | 3.72 | 0.0012 | |
| Length of main shoot | 6 | 351.866 | 12.96 | 0.0000 | |
| IBA (mg L−1) | Auxin Treatment + PGR-Free Liquid Medium = Duration of Experiment (Day) | Explants No. | Shoot with Root Primordia (%) | Rooting (%) | Roots Per Rooted Explant ± SE | Length of the Longest Root (mm) ± SE |
|---|---|---|---|---|---|---|
| 0.2 | 50 + 55 = 105 | 40 | 12.5 | 17.5 | 2.57 ± 0.84 a | 7.29 ± 1.49 a |
| 0.5 | 50 + 55 = 105 | 52 | 13.5 | 7.7 | 2.25 ± 0.48 a | 8.75 ± 2.56 a |
| 1.0 | 50 + 55 = 105 | 50 | 16.0 | 14.0 | 3.43 ± 0.95 a | 5.86 ± 2.41 a |
| 2.0 | 50 + 55 = 105 | 50 | 6.0 | 6.0 | 2.33 ± 0.33 a | 10.51 ± 7.51 a |
| 1.0 | 14 + 66 = 80 | 34 | 8.8 | 0 | - | - |
| 1.0 | 21 + 59 = 80 | 37 | 8.1 | 3.3 | 2.0 ± 0 | 2.0 ± 0 |
| 1.0 | 28 + 52 = 80 | 45 | 13.3 | 0 | - | - |
| 2.0 | 14 + 66 = 80 | 30 | 16.7 | 6.7 | 4.5 ± 0.5 a | 14.0 ± 2.0 a |
| 2.0 | 21 + 59 = 80 | 30 | 16.7 | 13.3 | 5.25 ± 1.1 a | 13.75 ± 4.27 a |
| 2.0 | 28 + 52 = 80 | 33 | 6.1 | 0 | - | - |
| 5.0 | 1 + 61 = 62 | 30 | 13.3 | 30.0 | 2.89 ± 0.5 a | 12.4 ± 2.02 a |
| 5.0 | 2 + 61 = 63 | 30 | 23.3 | 33.3 | 5.4 ± 1.1 b | 11.2 ± 2.6 a |
| 5.0 | 4 + 61 = 65 | 30 | 16.7 | 33.3 | 3.5 ± 0.6 ab | 7.7 ± 1.23 a |
| 5.0 | 6 + 59 = 65 | 30 | 10.0 | 16.7 | 2.4 ± 1.2 a | 7.6 ± 1.21 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krstić-Milošević, D.; Banjac, N.; Janković, T.; Vinterhalter, D.; Vinterhalter, B. Gentianella lutescens subsp. carpatica J. Holub.: Shoot Propagation In Vitro and Effect of Sucrose and Elicitors on Xanthones Production. Plants 2021, 10, 1651. https://doi.org/10.3390/plants10081651
Krstić-Milošević D, Banjac N, Janković T, Vinterhalter D, Vinterhalter B. Gentianella lutescens subsp. carpatica J. Holub.: Shoot Propagation In Vitro and Effect of Sucrose and Elicitors on Xanthones Production. Plants. 2021; 10(8):1651. https://doi.org/10.3390/plants10081651
Chicago/Turabian StyleKrstić-Milošević, Dijana, Nevena Banjac, Teodora Janković, Dragan Vinterhalter, and Branka Vinterhalter. 2021. "Gentianella lutescens subsp. carpatica J. Holub.: Shoot Propagation In Vitro and Effect of Sucrose and Elicitors on Xanthones Production" Plants 10, no. 8: 1651. https://doi.org/10.3390/plants10081651
APA StyleKrstić-Milošević, D., Banjac, N., Janković, T., Vinterhalter, D., & Vinterhalter, B. (2021). Gentianella lutescens subsp. carpatica J. Holub.: Shoot Propagation In Vitro and Effect of Sucrose and Elicitors on Xanthones Production. Plants, 10(8), 1651. https://doi.org/10.3390/plants10081651