
Influence of Nano Silicon and Nano Selenium on Root Characters, Growth, Ion Selectivity, Yield, and Yield Components of Rice (Oryza sativa L.) under Salinity Conditions


 and
and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Treatments, Plant Materials, and Soil Analysis
2.2. Measurements
- Root characteristics: At heading, plant samples involving root were carefully taken to determine root length (cm), root volume (cm3), and root thickness (mm). Root length was determined as the length of the root from the base of the plant to the tip of the main axis of primary root. Volume of the root system per plant was determined in cubic centimeters using a standard column. To measure root thickness, the average diameter (mm) of the tip portion (about 1 cm from the tip) of three random secondary roots at the middle position of the plant root was estimated.
- Growth and physiological characters: Leaf relative water content (RWC %) was calculated based on the described methods by Yamasaki and Dillenburg [41].
- 3.
- Ion selectivity: At heading, leaf samples were taken into the laboratory to analyze the Na+ and K+ contents as mg g−1 dry weight as well as to measure the Na+/K+ ratio according to Jackson [42].
- 4.
- Yield and its components: At harvest, number of panicles hill−1, panicle length (cm), number of unfilled grains panicle−1, number of filled grains panicle−1, 1000-grain weight (g), and grain weight/ panicle (g) were estimated from each plot. For grain yield, six inner rows from each plot were harvested, dried, threshed, and the grain, straw, and biological yields were determined. Then, yields/ha were calculated at 14% moisture content. Harvest index % was estimated as follows:
2.3. Statistical Analysis
3. Results
3.1. Root Characters
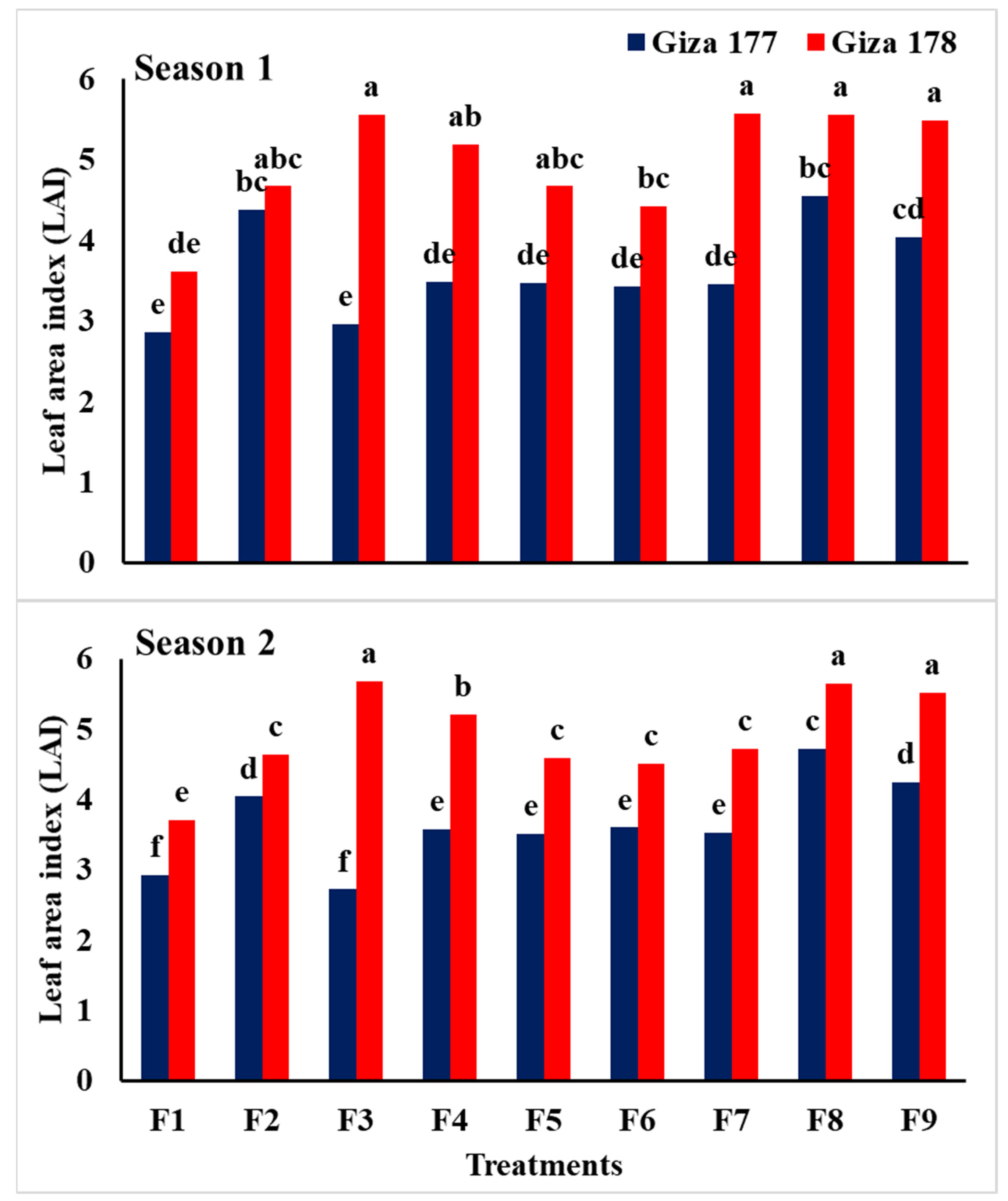
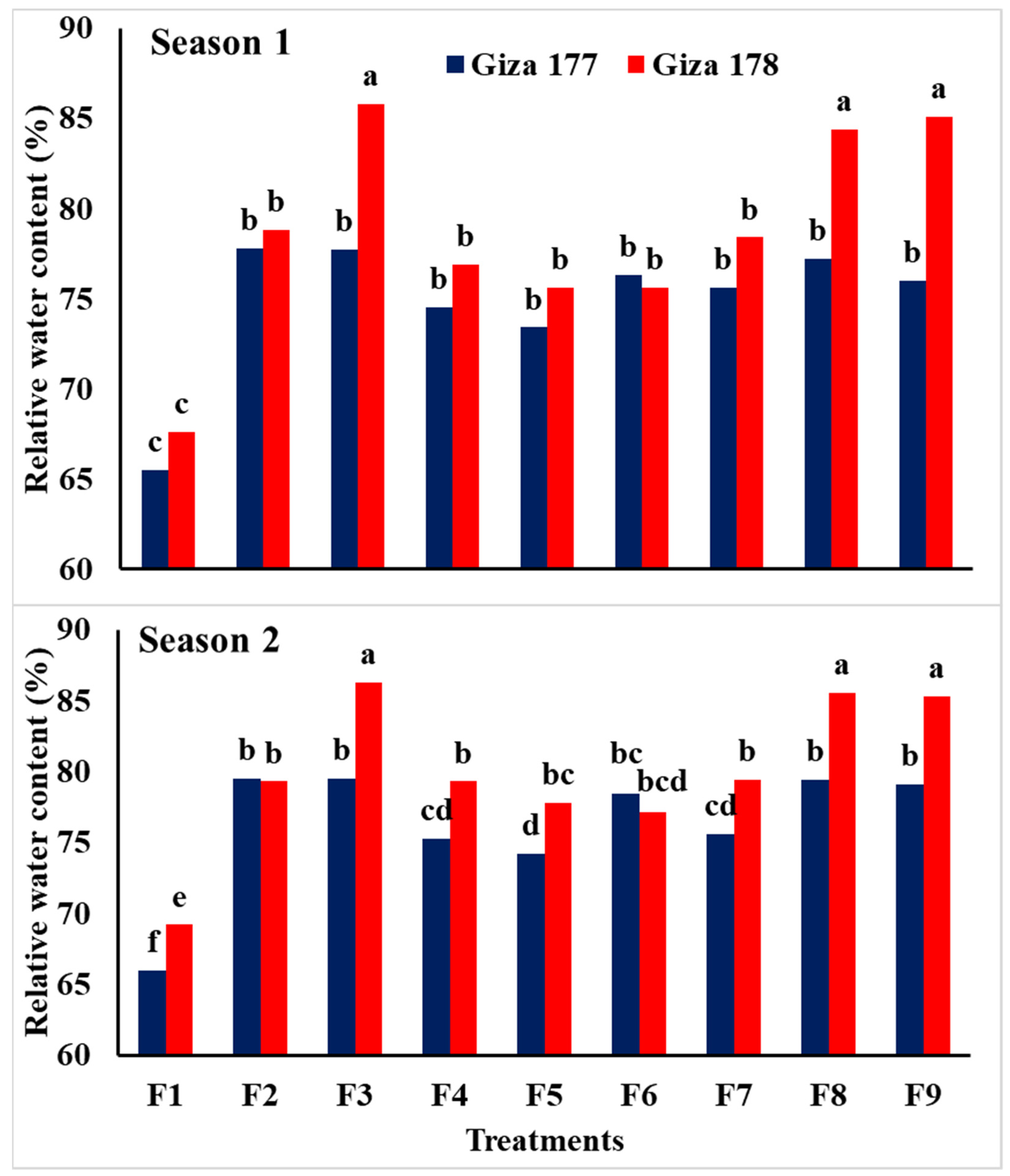
3.2. Growth and Physiological Traits
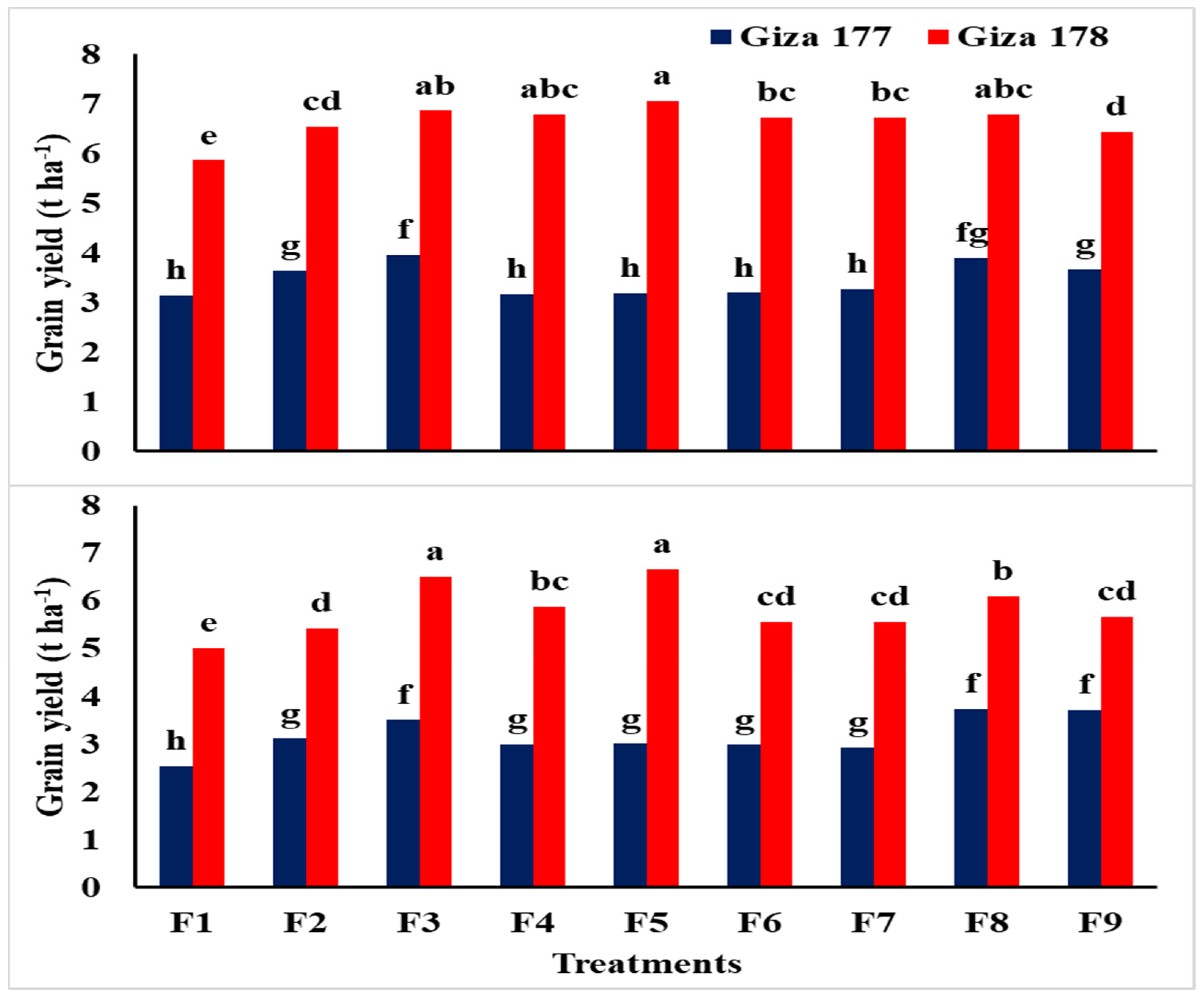
3.3. Yield and Yield Components
3.4. Ion Selectivity
4. Discussion
4.1. Root Characters
4.2. Growth and Physiological Traits
4.3. Yield and Yield Components
4.4. Ion Selectivity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Taha, R.S.; Seleiman, M.F.; Alhammad, B.A.; Alkahtani, J.; Alwahibi, M.S.; Mahdi, A.H.A. Activated Yeast Extract Enhances Growth, Anatomical Structure, and Productivity of Lupinus termis L. Plants under Actual Salinity Conditions. Agronomy 2021, 11, 74. [Google Scholar] [CrossRef]
- Zheng, L.; Shannon, M.C.; Lesch, S.M. Timing of salinity stress affecting rice growth and yield components. Agri. Water Manag. 2001, 48, 191–206. [Google Scholar] [CrossRef]
- Zayed, B.A.; Okasha, A.M.; Rashwan, E. Impact of different rates of phosphoric acid foliar applicationing on rice growth and yield traits under normal and saline soils conditions. J. Agri. Life Sci. 2019, 2, 56–66. [Google Scholar]
- Maas, E.V.; Hoffman, G.J. Crop salt tolerance-current assessment. J. Irrig. Drain. Div. 1986, 103, 115–134. [Google Scholar] [CrossRef]
- Heenan, D.P.; Lewin, L.G.; McCaffery, D.W. Salinity tolerance in rice varieties at different growth stages. Aust. J. Exp. Agric. 1988, 28, 343–349. [Google Scholar] [CrossRef]
- Lutts, S.; Kinet, J.M.; Bouharmont, J. Changes in plant response to NaCl during development of rice (Oryza sativus L.) varieties differing in salinity resistance. J. Exp. Bot. 1995, 46, 1843–1852. [Google Scholar] [CrossRef]
- Khan, M.S.A.; Hamid, A.; Karim, M.A. Effect of sodium chloride on germination and seedling characters of different types of rice (Oryza sativa L.). J. Agron. Crop Sci. 1997, 179, 163–169. [Google Scholar] [CrossRef]
- Mariko, M.; Fournier, J.M.; Benlloch, M. Strategies underlying salt tolerance in halophytes are present in Cynara cardunculus. Plant Sci. 1992, 168, 653–659. [Google Scholar]
- Baldmin, K.C.; Gallagher, J.L. Morphological and physiological responses to increased salinity in marsh and dune ecotypes of Sporobolus virginicus (L.) Kunth. Oecologia 1996, 87, 330–335. [Google Scholar]
- Grieve, C.M.; Suarez, D.L. Purslane (Portulaca oleracea L.): A halophytic crop for drainage water reuse systems. Plant Soil 1997, 192, 277–283. [Google Scholar] [CrossRef]
- Lutts, S.; Kinet, J.M.; Bouharmont, J. NaCl-induced senescence in leaves of rice (Oryza sativa L.) varieties, differing in salinity resistance. Ann. Bot. 1996, 78, 389–398. [Google Scholar] [CrossRef]
- Taha, R.S.; Seleiman, M.F.; Alotaibi, M.; Alhammad, B.A.; Rady, M.M.; Mahdi, A.H.A. Exogenous potassium treatments elevate salt tolerance and performances of Glycine max L. by boosting antioxidant defense system under actual saline field conditions. Agronomy 2020, 10, 1741. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Semida, W.M.; Rady, M.M.; Mohamed, G.F.; Hemida, K.A.; Alhammad, B.A.; Hassan, M.M.; Shami, A. Sequential Application of Antioxidants Rectifies Ion Imbalance and Strengthens Antioxidant Systems in Salt-Stressed Cucumber. Plants 2020, 9, 1783. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, R.; Mohammakhani, A.; Roohi, V.; Armand, N. Effects of salt stress and silicon nutrition on cholorophyll content, yield and yield components in fennel (Foeniculum vulgar Mill.). Intl. J. Agric. Crop Sci. 2012, 4, 1591–1595. [Google Scholar]
- Seleiman, M.F. Use of plant nutrients in improving abiotic stress tolerance in wheat. In Wheat Production in Changing Environments; Hasanuzzaman, M., Nahar, K., Hossain, M.A., Eds.; Springer: Singapore, 2019; pp. 481–495. [Google Scholar]
- Taha, R.S.; Seleiman, M.F.; Shami, A.; Alhammad, B.A.; Mahdi, A.H.A. Integrated Application of Selenium and Silicon Enhances Growth and Anatomical Structure, Antioxidant Defense System and Yield of Wheat Grown in Salt-Stressed Soil. Plants 2021, 10, 1040. [Google Scholar] [CrossRef] [PubMed]
- Al-Aghabary, K.; Zhu, Z.; Shi, Q.H. Infuence of silicon supply on chlorophyll content, chlorophyll fluorescence, and antioxidative enzyme activities in tomato plants under salt stress. J. Plant Nutr. 2004, 27, 2101–2115. [Google Scholar] [CrossRef]
- Liang, Y.C.; Zhang, W.H.; Chen, Q.; Ding, R.X. Effects of silicon on tonoplast Hþ–ATPase and Hþ–PPase activity, fatty acid composition and fluidity in roots of salt stressed barley (Hordeum vulgare L.). Environ Exp. Bot. 2005, 53, 29–37. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Abdelaal, M.S. Effect of organic, inorganic and bio-fertilization on growth, yield and quality traits of some chickpea (Cicer arietinum L.) varieties. Egypt. J. Agron. 2018, 40, 105–117. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Almutairi, K.F.; Alotaibi, M.; Shami, A.; Alhammad, B.A.; Battaglia, M.L. Nano-Fertilization as an Emerging Fertilization Technique: Why Can Modern Agriculture Benefit from Its Use? Plants 2021, 10, 2. [Google Scholar] [CrossRef]
- Zahedi, S.M.; Moharrami, F.; Sarikhani, S.; Padervand, M. Selenium and silica nanostructure-based recovery of strawberry plants subjected to drought stress. Sci. Rep. 2020, 10, 1–18. [Google Scholar] [CrossRef]
- Alkharabsheh, H.M.; Seleiman, M.F.; Battaglia, M.L.; Shami, A.; Jalal, R.S.; Alhammad, B.A.; Almutairi, K.F.; Al-Saif, A.M. Biochar and Its Broad Impacts in Soil Quality and Fertility, Nutrient Leaching and Crop Productivity: A Review. Agronomy 2021, 11, 993. [Google Scholar] [CrossRef]
- Cheng, G.; Chang, T.; Qin, Q.; Huang, H.; Zhu, Y. Mechanical Properties of Silicon Carbide Nanowires: Effect of Size Dependent Defect Density. Nano Let. 2014, 14, 754–758. [Google Scholar] [CrossRef] [PubMed]
- Torney, F.; Trewyn, B.G.; Lin, V.S.Y.; Wang, K. Mesoporous silica nanoparticles deliver DNA and chemicals into plants. Nature Nano Tech. 2007, 2, 295–300. [Google Scholar] [CrossRef]
- Bao-Shan, L.; Chun-hui, L.; Li-jun, F.; Shu-chun, Q.; Min, Y. Effects of TMS (nanostructured silicon dioxide) on growth of Changbai larch seedlings. J. Forest Res. 2004, 15, 138–140. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Alotaibi, M.A.; Alhammad, B.A.; Alharbi, B.M.; Refay, Y.; Shimaa, A.B. Effects of ZnO Nanoparticles and Biochar of Rice Straw and Cow Manure on Characteristics of Contaminated Soil and Sunflower Productivity, Oil Quality, and Heavy Metals Uptake. Agronomy 2020, 10, 790. [Google Scholar] [CrossRef]
- Suriyaprabha, R.; Karunakaran, G.; Yuvakkumar, R.; Rajendran, V.; Kannan, N. Silica nanoparticles for increased silica availability in maize (Zea mays L.) seeds under hydroponic conditions. Curr. Nano Sci. 2012, 8, 1–7. [Google Scholar] [CrossRef]
- Chauhan, R.; Awasthi, S.; Srivastava, S.; Dwivedi, S.; Pilon-Smits, E.A.H.; Dhankher, O.P.; Tripathi, R.D. Understanding selenium metabolism in plants and its role as a beneficial element. Crit. Rev. Environ. Sci. Technol. 2019, 49, 1937–1958. [Google Scholar] [CrossRef]
- Szarka, V.; Jokai, Z.; El-Ramady, H.; Abdalla, N.; Kaszás, L.; Domokos-Szabolcsy, E. Biofortification of Stevia rebaudiana (Bert.) plant with selenium. Environ. Biodivers. Soil Secur. 2020, 4, 19–31. [Google Scholar] [CrossRef]
- Reis, A.R.; Boleta, E.H.M.; Alves, C.Z.; Cotrim, M.F.; Barbosa, J.Z.; Silva, V.M.; Porto, R.L.; Lanza, M.G.D.; Lavres, J.; Gomes, M.H.F.; et al. Selenium toxicity in upland field-grown rice: Seed physiology responses and nutrient distribution using the μ-XRF technique. Ecotoxicol. Environ. Saf. 2020, 190, 110–147. [Google Scholar] [CrossRef]
- El-Ramady, H.; Alshaal, T.; Elhawat, N.; El-Nahrawy, E.; Omara, A.; El-Nahrawy, S.; Elsakhawy, T.; Azza Ghazi, A.; Abdalla, N.; Fári, M. Biological aspects of selenium and silicon nanoparticles in the terrestrial environments. In Phytoremediation; Springer: Cham, Switzerland, 2018; pp. 235–264. [Google Scholar] [CrossRef]
- Hussein, H.A.A.; Darwesh, O.M.; Mekki, B.B. Environmentally friendly nano- selenium to improve antioxidant system and growth of groundnut cultivars under sandy soil conditions. Biocatal. Agric. Biotechnol. 2019, 18, 101080. [Google Scholar] [CrossRef]
- Zahedi, S.M.; Hosseini, M.S.; Daneshvar Hakimi Meybodi, N.; Teixeira da Silva, J.A. Foliar application of selenium and nano-selenium affects pomegranate (Punica granatum cv. Malase Saveh) fruit yield and quality. South Afr. J. Bot. 2019, 124, 350–358. [Google Scholar] [CrossRef]
- Zahedi, S.M.; Abdelrahman, M.; Hosseini, M.S.; Hoveizeh, N.F.; Tran, L.-S.P. Alleviation of the effect of salinity on growth and yield of strawberry by foliar spray of selenium-nanoparticles. Environ. Pollut. 2019, 253, 246–258. [Google Scholar] [CrossRef]
- Soleymanzadeh, R.; Iranbakhsh, A.; Habibi, G.; Ardebili, Z.O. Selenium nanoparticle protected strawberry against salt stress through modifications in salicylic acid, ion homeostasis, antioxidant machinery, and photosynthesis performance. Acta Biol. Crac. Bot. Ser. Bot. 2020, 62, 33–42. [Google Scholar]
- Elsheery, N.I.; Helaly, M.N.; Omar, S.A.; John, S.V.S.; Zabochnicka-Swiątek, M.; Kalaji, H.M.; Rastogi, A. Physiological and molecular mechanisms of salinity tolerance in grafted cucumber. South Afr. J. Bot. 2020, 130, 90–102. [Google Scholar] [CrossRef]
- Naderi, M.R.; Danesh-Sharaki, A. Nanofertilizers and their role in sustainable agriculture. Int. J. Agric. Crop. Sci. 2013, 5, 2229–2232. [Google Scholar]
- Liu, R.; Lal, R. Potentials of engineered nanoparticles as fertilizers for increasing agronomic productions. A review. Sci. Total. Environ. 2015, 514, 131–139. [Google Scholar] [CrossRef]
- Eszenyi, P.; Sztrik, A.; Babka, B.; Prokisch, J. Elemental, nano-sized (100–500 nm) selenium production by probiotic lactic acid bacteria. Int. J. Biosci. Biochem. Bioinform. 2011, 1, 148–152. [Google Scholar] [CrossRef] [Green Version]
- Black, C.A.; Evans, D.D.; Ensminger, L.E.; Clark, F.E. Methods of Soil Analysis. Part 2—Chemical and Microbiological Properties; John Wiley & Sons: Hoboken, NJ, USA, 1965. [Google Scholar]
- Yamasaki, S.; Dillenburg, L.C. Measurements of Leaf Relative Water Content in Araucaria angustifolia. Braz. J. Plant Physiol. 1999, 11, 69–75. [Google Scholar]
- Jackson, M.L.A. Simplified assay for milled rice analysis. J. Cereal Sci. 1971, 16, 334–340. [Google Scholar]
- Gomes, K.A.; Gomes, A.A. Statistical Procedures for Agricultural Research, 2nd ed.; John Wily and Sons: New York, NY, USA, 1984; p. 680. [Google Scholar]
- Duncan, B.D. Multiple ranges and multiple F-tests. Biometrics 1995, 11, 42. [Google Scholar] [CrossRef]
- Abbas, T.; Rizwan, M.; Ali, S.; Adrees, M.; Mahmood, A.; Zia-ur-Rehman, M.; Ibrahim, M.; Arshad, M.; Qayyum, M.F. Biochar application increased the growth and yield and reduced cadmium in drought stressed wheat grown in an aged contaminated soil. Ecotoxicol. Environ. Saf. 2018, 148, 825–833. [Google Scholar] [CrossRef]
- Gholami, R.; Zahedi, S.M. Identifying superior drought-tolerant olive genotypes and their biochemical and some physiological responses to various irrigation levels. J. Plant Nutr. 2019, 42, 2057–2069. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought Stress Impacts on Plants and Different Approaches to Alleviate Its Adverse Effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef] [PubMed]
- Seleiman, M.F.; Hafez, E.M. Optimizing Inputs Management for Sustainable Agricultural Development. In Mitigating Environmental Stresses for Agricultural Sustainability in Egypt; Springer Water; Springer: Cham, Switzerland, 2021; pp. 487–507. [Google Scholar]
- Rahman, M.S.; Miyake, H.; Taheoka, Y. Effect of sodium chloride salinity on seed germination and early seedling growth of rice (Oryza sativa L.). Pak. J. Biol. Sci. 2001, 4, 351–355. [Google Scholar] [CrossRef]
- Rasool, S.; Hameed, A.; Azooz, M.M.; Muneeb-u-Rehman; Siddiqi, T.O.; Ahmad, P. Salt stress: Causes, types and responses of plants. In Ecophysiology and Responses of Plants under Salt Stress; Springer: New York, NY, USA, 2013; pp. 1–24. [Google Scholar]
- Seleiman, M.F.; Kheir, A.M. Maize productivity, heavy metals uptake and their availability in contaminated clay and sandy alkaline soils as affected by inorganic and organic amendments. Chemosphere 2018, 204, 514–522. [Google Scholar] [CrossRef]
- Kalteh, M.; Alipour, Z.T.; Ashraf, S.; Aliabadi, M.M.; Nosratabadi, A.F. Effect of silica nanoparticles on basil (Ocimum basilicum) under salinity stress. J. Chem. Health Risks 2018, 4, 49–55. [Google Scholar] [CrossRef]
- Gong, H.; Chen, K.; Chen, G.; Wang, S.; Zhang, C. Effects of silicon on growth of wheat under drought. J. Plant Nutr. 2003, 26, 1055–1063. [Google Scholar] [CrossRef]
- Ma, J.F.; Tamai, K.; Yamaji, N.; Mitani, N.; Konishi, S.; Katsuhara, M. A silicon transporter in rice. Nature 2006, 440, 688–691. [Google Scholar] [CrossRef] [PubMed]
- Shahzade, M. Growth-Related Changes in Subcellularion and Protein Patterns in Maiz and Field Bean Leaves under Salt Stress; MSc Institute of Plant Nutrition and Soil Science, University of Kiel: Kiel, Germany, 2012. [Google Scholar]
- Parveen, N.; Ashraf, M. Role of silicon in mitigating the adverse effects ofsalt stress on growth and photosynthetic attributes of two maize (Zea mays L.) cultivars grown hydroponically. Pak. J. Bot. 2010, 42, 1675–1684. [Google Scholar]
- Haghighi, M.; Afifipour, Z.; Mozafarian, M. The effect of N-Si on tomato seed germination under salinity levels. J. Biol. Environ. Sci. 2012, 6, 87–90. [Google Scholar]
- Zayed, B.A.; Rania, K.; Hadifa, A.A.; Amira, M.O. Some anti-oxidants, Phsysio-morphological, and Yield of Varying Rice Varieties Affected by Salinity Levels. J. Plant Prod. Mansoura Univ. 2017, 8, 747–754. [Google Scholar] [CrossRef] [Green Version]
- Zahedi, S.M.; Karimi, M.; da Silva, J.A.T. The use of nanotechnology to increase quality and yield of fruit crops. J. Sci. Food Agric. 2019, 100, 25–31. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Fujita, M. Silicon and selenium: Two vital trace elements that confer abiotic stress tolerance to plants. In Emerging Technologies and Management of Crop Stress Tolerance; Elsevier: Amsterdam, The Netherlands, 2014; pp. 377–422. [Google Scholar]
- Yaghubi, K.; Vafaee, Y.; Ghaderi, N.; Javadi, T. Potassium silicate improves salinity resistant and affects fruit quality in two strawberry cultivars grown under salt stress. Commun. Soil. Sci. Plant Anal. 2019, 50, 1439–1451. [Google Scholar] [CrossRef]
- Al-Ashkar, I.; Alderfasi, A.; El-Hendawy, S.; Al-Suhaibani, N.; El-Kafafi, S.; Seleiman, M.F. Detecting Salt Tolerance in Doubled Haploid Wheat Lines. Agronomy 2019, 9, 211. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Zou, C.; Wang, L.; Zhang, F. Silicon decreases transpiration rate and conductance from stomata of maize plants. J. Plant. Nutr. 2006, 29, 1637–1647. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–668. [Google Scholar] [CrossRef] [Green Version]
- Kardoni, F.; Mosavi, S.J.S.; Parande, S.; Torbaghan, M.E. Effect of salinity stress and silicon application on yield and component yield of Faba bean (Vicia faba). Inter. J. Agric. Crop Sci. 2013, 6, 814–818. [Google Scholar]
- Tahir, M.A.; Rammatullah, A.; Ashraf, T.M.; Kanwal, S.; Maqsood, M.A. Beneficial effects of silicon in wheat (Triticum aestivum) under salinity stress. Pak. J. Bot. 2006, 38, 715–1722. [Google Scholar]
- Miyake, Y.; Takahashi, E. Effect of silicon on the growth and fruit production of strawberry plants in a solution culture. Soil Sci. Plant Nurt. 1986, 32, 321–326. [Google Scholar] [CrossRef] [Green Version]
- Meena, V.D.; Dotaniya, M.L.; Coumar, V.; Rajendiran, S.; Kundu, S.; Subba Rao, A. A Case for Silicon Fertilization to Improve Crop Yields in Tropical Soils. Biol Sci. 2014, 84, 505–518. [Google Scholar] [CrossRef] [Green Version]
- Seleiman, M.F.; Refay, Y.; Al-Suhaibani, N.; Al-Ashkar, I.; El-Hendawy, S.; Hafez, E.M. Integrative Effects of Rice-Straw Biochar and Silicon on Oil and Seed Quality, Yield and Physiological Traits of Helianthus annuus L. Grown under Water Deficit Stress. Agronomy 2019, 9, 637. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Wei, G.; Li, J.; Qian, Q.; Yu, J. Silicon alleviates salt stress and increases antioxidant enzymes activity in leaves of salt-stressed cucumber (Cucumis sativus L.). Plant Sci. 2004, 167, 527–533. [Google Scholar] [CrossRef]
- Moussa, H.R. Influence of exogenous application of silicon on physiological response of salt-stressed maize (Zea mays L.). Inter. J. Agric. Biol. 2006, 8, 293–297. [Google Scholar]
- Romero-Arnada, M.R.; Jourado, O.; Cuartero, J. Silicon alleviates the deleterious salt effects on tomato plant growth by improving plant water status. J. Plant Phy. 2006, 163, 847–855. [Google Scholar] [CrossRef]
- Tuna, A.L.; Kaya, C.; Higgs, D.; Amador, B.M.; Aydemir, S.; Girgin, A.R. Silicon improves salinity tolerance in wheat plants. Environ. Exp. Bot. 2008, 62, 10–16. [Google Scholar] [CrossRef]
- Liang, Y.; Sun, W.; Zhu, Y.G.; Christie, P. Mechanisms of silicon mediated alleviation of abiotic stresses in higher plants: A review. Environ. Poll. 2007, 147, 422–428. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef] [Green Version]
- Kobraee, S.; Shamsi, K.; Rasekhi, B. Soybean production under water deficit conditions. Ann. Biol. Res. 2011, 2, 423–434. [Google Scholar]
- Ahmad, A.; Afzal, M.; Ahmad, A.U.H.; Tahir, M. Effect of foliar application of silicon on yield and quality of rice (Oryza sativa L.). Cercet. Agron. Mold. 2013, 3, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Parande, S.; Zamani, G.R.; Zahan, M.H.S.; Ghaderi, M.G. Effects of silicon application on the yield and component of yield in the common bean (Phaseolus vulgaris) under salinity stress. Inter. J. Agron. Plant Prod. 2013, 4, 1574–1579. [Google Scholar]
- Zhang, Y.; Fang, J.; Wu, X.; Dong, L. Na+/K+ balance and transport regulatory mechanisms in weedy and cultivated rice (Oryza sativa L.) under salt stress. BMC Plant Biol. 2018, 18, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Savvas, D.; Giotis, D.; Chatzieustratiou, E.; Bakea, M.; Patakioutas, G. Silicon supply in soilless cultivations of zucchini alleviates stress induced by salinity and powdery mildew infections. Environ. Exp. Bot. 2009, 65, 11–17. [Google Scholar] [CrossRef]
- EL-Habet, H.B. Role of silica in mitigation of Cd, Pb, and Cr toxicities in rice under irrigation with drainage water in the Egypt Nile delta. Irrig. Drain 2020, 70, 52–69. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Parameters | pH | EC (dS m−1) | OM (%) | Soluble Cations (meq L−1) | Soluble Anions (meq L−1) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Seasons | Ca++ | Mg++ | K+ | Na+ | HCO3− | Cl− | SO42− | ||||
| 2018 | 8.20 | 7.20 | 162 | 10.8 | 8.4 | 0.50 | 64 | 9.34 | 68.5 | 6.33 | |
| 2019 | 8.26 | 7.00 | 1.60 | 6.4 | 6.2 | 0.70 | 66 | 8.64 | 64.6 | 5.33 | |
| Traits | Root Thickness (mm) | Root Volume (cm3) | Root Length (cm) | ||||
|---|---|---|---|---|---|---|---|
| Seasons | S1 | S2 | S1 | S2 | S1 | S2 | |
| Rice Varieties | |||||||
| Giza 177 | 0.747 b | 0.763 b | 92.02 b | 93.5 b | 18.1 b | 19.0 b | |
| Giza 178 | 0.798 a | 0.810 a | 130.0 a | 129.6 a | 25.5 a | 26.2 a | |
| F test | ** | ** | ** | ** | ** | ** | |
| NPs-Si and NPs-Se Treatments | |||||||
| Control | 0.426 c | 0.486 e | 74.9 f | 75.8 f | 16.6 c | 17.5 c | |
| Soaking-NPs-Si | 0.78 b | 0.821 b,c | 113.3 c,d | 107.6 d | 22.1 b | 22.5 b | |
| Soaking-NPs-Se | 0.866 a | 0.923 a | 148.3 a | 147.8 a | 26.1 a | 27.3 a | |
| Foliar-NPs-Si at MT | 0.778 b | 0.740 d | 93.30 e | 94.80 e | 20.5 b | 21.3 b | |
| Foliar-NPs-Se at MT | 0.781 b | 0.743 d | 104.1 d | 106.8 d | 20.8 b | 21.6 b | |
| Foliar-NPs-Si at PI | 0.843 a,b | 0.793 c,d | 105.0 d | 106.1 d | 21.3 b | 22.0 b | |
| Foliar-NPs-Se at PI | 0.793 b | 0.855 b,c | 111.6 c,d | 128.1 b | 21.8 b | 22.6 b | |
| Foliar-NPs-Si at BT | 0.846 a,b | 0.876 a,b | 130.0 b | 119.1 c | 23.8 a,b | 24.6 b | |
| Foliar-NPs-Se at BT | 0.84 a,b | 0.845 b,c | 118.3 c | 117.6 c | 23.5 a,b | 24.1 b | |
| F test | ** | ** | ** | ** | ** | ** | |
| Interaction | ** | ** | ** | ** | ns | ns | |
| Traits | Root Thickness (mm) | Root Volume (cm3) | |||
|---|---|---|---|---|---|
| Treatments | Giza 177 | Giza 178 | Giza 177 | Giza 178 | |
| Season 2018 | |||||
| Control | 0.29 d | 0.56 c | 56.60 g | 93.30 f | |
| Soaking-NPs-Si | 0.79 a,b | 0.76 b | 86.60 f | 140.00 a,b | |
| Soaking-NPs-Se | 0.83 a,b | 0.90 a | 143.30 a,b | 153.30 a | |
| Foliar-NPs-Si at MT | 0.75 b | 0.80 a,b | 80.10 f | 106.60 e | |
| Foliar-NPs-Se at MT | 0.75 b | 0.81 a,b | 81.60 f | 126.60 c,d | |
| Foliar-NPs-Si at PI | 0.84 a,b | 0.84 a,b | 83.30 f | 126.60 c,d | |
| Foliar-NPs-Se at PI | 0.79 a,b | 0.79 a,b | 86.60 f | 136.60 b,c | |
| Foliar-NPs-Si at BT | 0.84 a,b | 0.85 a,b | 116.60 d | 143.30 a,b | |
| Foliar-NPs-Se at BT | 0.83 a,b | 0.85 a,b | 93.30 f | 143.30 a,b | |
| Season 2019 | |||||
| Control | 0.336 f | 0.636 e | 57.30 j | 94.30 g | |
| Soaking-NPs-Si | 0.82 b,c,d | 0.82 b,c,d | 87.30 h | 128.00 d | |
| Soaking-NPs-Se | 0.89 a,b | 0.95 a,b | 141.30 b,c | 154.30 a | |
| Foliar- NPs-Si at MT | 0.76 c,d | 0.71 d | 81.60 i | 108.00 f | |
| Foliar- NPs-Se at MT | 0.75 c,d | 0.73 c,d | 85.60 h,i | 128.00 d | |
| Foliar- NPs-Si at PI | 0.79 b,c,d | 0.79 b,c,d | 85.00 h,i | 127.30 d | |
| Foliar- NPs-Se at PI | 0.80 b,c,d | 0.90 a,b | 118.00 e | 138.30 c | |
| Foliar- NPs-Si at BT | 0.85 a,b,c | 0.90 a,b | 94.00 g | 144.30 b | |
| Foliar- NPs-Se at BT | 0.83 a,b,c | 0.84 a,b,c | 91.30 g | 144.00 b | |
| Traits | Relative Water Content (%) | Leaf Area Index (LAI) | Chlorophyll Content (SPAD) | ||||
|---|---|---|---|---|---|---|---|
| Seasons | |||||||
| Seasons | S1 | S2 | S1 | S2 | S1 | S2 | |
| Rice varieties | |||||||
| Giza 177 | 74.9 b | 76.3 b | 3.62 b | 3.65 b | 41.16 b | 41.9 b | |
| Giza 178 | 78.7 a | 79.9 a | 4.86 a | 4.91 a | 42.77 a | 42.3 a | |
| F test | ** | ** | ** | ** | ** | ** | |
| NPs-Si and NPs-Se treatments | |||||||
| Control | 66.5 c | 67.6 d | 3.23 d | 3.32 e | 38.6 c | 38.6 d | |
| Soaking-NPs-Si | 78.3 a,b | 79.4 b | 4.52 b,c | 4.33 c | 42.2 b | 42.3 b,c | |
| Soaking-NPs-Se | 81.7 a | 82.9 a | 4.26 b,c | 4.20 c,d | 44.1 a | 43.7 a | |
| Foliar-NPs-Si at MT | 75.7 b | 77.3 b,c | 4.33 b,c | 4.39 c | 42.15 b | 42.5 a,b,c | |
| Foliar-NPs-Se at MT | 74.5 b | 76.0 c | 4.07 c | 4.04 d | 41.9 b | 41.7 c | |
| Foliar-NPs-Si at PI | 75.9 b | 77.7 b,c | 3.93 c | 4.06 d | 41.5 b | 42.2 b,c | |
| Foliar-NPs-Se at PI | 77.0 b | 77.5 b,c | 4.01 c | 4.12 d | 41.6 b | 42.1 b,c | |
| Foliar-NPs-Si at BT | 80.8 a | 82.4 a | 5.06 a | 5.18 a | 42.6 b | 43.4 a,b | |
| Foliar-NPs-Se at BT | 80.6 a | 82.2 a | 4.76 a,b | 4.87 b | 42.7 b | 42.6 a,b,c | |
| F test | ** | ** | ** | ** | * | ** | |
| Interaction | ** | ** | ** | ** | ns | ns | |
| Traits | Dry Matter (g Hill−1) | Plant Height (cm) | |||
|---|---|---|---|---|---|
| Seasons | S1 | S2 | S1 | S2 | |
| Rice varieties | |||||
| Giza 177 | 28.9 b | 30.3 b | 85.0 b | 86.0 b | |
| Giza 178 | 38.5 a | 39.7 a | 87.6 a | 88.1 a | |
| F test | ** | ** | ** | ** | |
| NPs-Si and NPs-Se treatments | |||||
| Control | 27.4 c | 27.6 d | 83.6 c | 84.4 c | |
| Soaking-NPs-Si | 34.5 a,b | 35.3 b | 85.0 b,c | 85.7 b,c | |
| Soaking-NPs-Se | 39.7 a | 40.6 a | 86.2 a,b | 86.9 a,b | |
| Foliar-NPs-Si at MT | 33.1 a,b,c | 33.4 b,c | 86.6 a,b | 87.4 a,b | |
| Foliar-NPs-Se at MT | 31.6 b,c | 33.7 b,c | 86.7 a,b | 87.4 a,b | |
| Foliar-NPs-Si at PI | 31.1 b,c | 31.7 c | 86.6 a,b | 87.3 a,b | |
| Foliar-NPs-Se at PI | 33.1 a,b,c | 35.2 b | 87.8 a | 88.5 a | |
| Foliar-NPs-Si at BT | 37.3 a,b | 39.1 a | 87.9 a | 88.7 a | |
| Foliar-NPs-Se at BT | 35.6 a,b | 38.3 a | 86.5 a,b | 87.2 a,b | |
| F test | ** | ** | ** | ** | |
| Interaction | ns | ns | ns | ns | |
| Traits | Number of Panicles Hill−1 | Panicle Weight | Panicle Length | ||||
|---|---|---|---|---|---|---|---|
| Seasons | S1 | S2 | S1 | S2 | S1 | S2 | |
| Rice varieties | |||||||
| Giza 177 | 16.37 b | 15.6 b | 2.80 a | 2.70 | 19.9 b | 18.4 b | |
| Giza 178 | 21.8 a | 20.4 a | 2.64 b | 2.47 | 21.3 a | 21.4 a | |
| F test | ** | ** | ** | ns | ** | ** | |
| NPs-Si and NPs-Se treatments | |||||||
| Control | 17.00 c | 16.2 c | 2.41 d | 2.17 c | 19.61 | 19.3 | |
| Soaking-NPs-Si | 18.9 a,b | 18.06 b,c | 2.83 a,b,c | 2.54 b | 21.0 | 20.4 | |
| Soaking-NPs-Se | 20.4 a | 20.7 a | 2.99 a | 2.91 a | 21.3 | 20.4 | |
| Foliar-NPs-Si at MT | 18.5 a,b | 19.1 a,b | 2.69 b,c | 2.58 b | 20.6 | 19.8 | |
| Foliar-NPs-Se at MT | 18.5 a,b | 17.9 b,c | 2.73 b,c | 2.47 b | 20.6 | 19.1 | |
| Foliar-NPs-Si at PI | 18.6 a,b | 17.4 b,c | 2.50 d | 2.60 b | 20.0 | 20.4 | |
| Foliar-NPs-Se at PI | 18.9 a,b | 18.3 b | 2.65 c | 2.56 b | 20.6 | 20.5 | |
| Foliar-NPs-Si at BT | 20.5 a | 19.2 a,b | 2.86 a,b | 2.78 a | 21.1 | 19.7 | |
| Foliar-NPs-Se at BT | 19.8 a | 17.8 b,c | 2.82 a,b,c | 2.80 a | 21.0 | 20.0 | |
| F test | ** | ** | ** | ** | ns | ns | |
| Interaction | ns | ns | ns | ns | ns | ns | |
| Traits | Number of Filled Grains Panicle−1 | Number of Unfilled Grains Panicle−1 | 1000-Grain Weight (g) | ||||
|---|---|---|---|---|---|---|---|
| Seasons | |||||||
| Seasons | S1 | S2 | S1 | S2 | S1 | S2 | |
| Rice varieties | |||||||
| Giza 177 | 115.42 b | 93.01 b | 24.54 a | 23.0 a | 25.9 a | 25.7 a | |
| Giza 178 | 127.0 a | 116.1 a | 17.91 b | 15.2 b | 18.7 b | 19.23 b | |
| F test | ** | ** | ** | ** | ** | ** | |
| NPs-Si and NPs-Se treatments | |||||||
| Control | 103.1 e | 90.9 d | 34.2 a | 32.8 a | 19.50 e | 19.40 c | |
| Soaking-NPs-Si | 123.4 c | 100.0 c | 21.9 b | 20.8 b | 22.2 b,c,d | 22.30 b | |
| Soaking-NPs-Se | 127.85 b | 108.0 b | 16.8 d | 16.6 c,d | 23.20 a | 23.50 a,b | |
| Foliar-NPs-Si at MT | 118.6 d | 102.7 c | 21.8 b | 19.6 b,c | 22.0 c,d | 22.20 b | |
| Foliar-NPs-Se at MT | 124.1 b | 104.8 b,c | 19.1 c | 20.9 b | 22.6 b,c | 22.40 b | |
| Foliar-NPs-Si at PI | 116.0 d | 103.8 b,c | 21.4 b | 16.6 c,d | 22.2 b,c,d | 22.10 b | |
| Foliar-NPs-Se at PI | 117.7 d | 103.9 b,c | 19.7 b,c | 17.2 b,c,d | 22.1 c,d | 22.30 b | |
| Foliar-NPs-Si at BT | 133.3 a | 114.1 a | 15.8 d | 12.1 e | 23.9 a | 24.60 a | |
| Foliar-NPs-Se at BT | 130.7 a,b | 113.6 a | 20.0 b,c | 15.2 d | 23.3 a | 23.80 a | |
| F test | ** | ** | ** | ** | ** | * | |
| Interaction | ** | ns | ** | ** | ns | ns | |
| Traits | Number of Filled Grains Panicle −1 | Number of Unfilled Grains Panicle−1 | |||||
|---|---|---|---|---|---|---|---|
| Treatments | Giza 177 | Giza 178 | Giza 177 | Giza 178 | Giza 177 | Giza 178 | |
| S1 | S1 | S2 | |||||
| Control | 91.7 g | 114.5 f | 39.3 a | 29.1 b | 42.16 a | 23.4 b | |
| Soaking-NPs-Si | 118.8 e,f | 127.9 b,c | 19.5 e,f,g | 24.1 c | 23.3 b,c | 18.4 b,c,d | |
| Soaking-NPs-Se | 123.1 d,e | 132.4 ab | 16.35 g,h | 17.37 f,g,h | 18.7 b,c,d | 14.2 c,d | |
| Foliar-NPs-Si at MT | 111.6 f | 125.5 b,c,d | 23.47 c,d | 20.03 d,e,f | 23.5 b | 15.7 c,d | |
| Foliar-NPs-Se at MT | 111.6 f | 130.10 a,b | 25.67 c | 12.76 i | 20.9 b,c | 20.90 b,c | |
| Foliar-NPs-Si at PI | 113.3 f | 118.7 e,f | 24.10 c | 18.9 e,f,g | 20.7 b,c | 12.4 d,e | |
| Foliar-NPs-Se at PI | 113.2 f | 122.2 c,d,e | 20.84 d,e,f | 18.8 e,f,g | 18.5 b,c,d | 16.0 c,d | |
| Foliar-NPs-Si at BT | 129.4 b | 136.6 a | 17.11 f,g,h | 14.30 h,i | 15.4 c,d | 8.76 e | |
| Foliar-NPs-Se at BT | 126.1 b,c | 135.1 a | 18.46 f,g | 21.90 c,d,e | 15.4 c,d | 15.0 c,d | |
| Traits | Grain Yield (t ha−1) | Biological Yield (t ha−1) | Harvest Index (%) | ||||
|---|---|---|---|---|---|---|---|
| Seasons | S1 | S2 | S1 | S2 | S1 | S2 | |
| Rice varieties | |||||||
| Giza 177 | 3.45 b | 3.16 b | 8.06 b | 10.1 b | 0.428 b | 0.310 b | |
| Giza 178 | 6.64 a | 5.81 a | 13.5 a | 12.6 a | 0.488 a | 0.456 a | |
| F test | ** | ** | ** | ** | ** | ** | |
| NPs-Si and NPs-Se treatments | |||||||
| Control | 4.49 c | 3.78 d | 9.94 d | 9.95 d | 0.445 b | 0.370 b | |
| Soaking-NPs-Si | 5.08 b | 4.26 c | 10.6 b,c | 10.7 c | 0.477 a | 0.388 a,b | |
| Soaking-NPs-Se | 5.41 a | 5.00 a | 11.5 a | 11.7 a,b | 0.464 a,b | 0.416 a | |
| Foliar-NPs-Si at MT | 4.97 b | 4.43 c | 10.4 c | 11.6 a,b | 0.464 a,b | 0.373 b | |
| Foliar-NPs-Se at MT | 5.12 b | 4.83 a,b | 11.0 b | 11.9 a | 0.452 a,b | 0.390 a,b | |
| Foliar-NPs-Si at PI | 4.96 b | 4.26 c | 10.6 b,c | 11.5 a,b | 0.455 a,b | 0.363 b | |
| Foliar-NPs-Se at PI | 4.99 b | 4.24 c | 10.7 b,c | 11.2 b | 0.456 a,b | 0.370 b | |
| Foliar-NPs-Si at BT | 5.34 a | 4.91 a | 11.4 a | 11.9 a | 0.460 a,b | 0.406 a | |
| Foliar-NPs-Se at BT | 5.04 b | 4.68 b | 10.9 b | 11.6 a,b | 0.453 a,b | 0.398 a,b | |
| F test | ** | ** | ** | ** | ** | ** | |
| Interaction | ** | ** | ns | ns | ns | ns | |
| Traits | Na | K+ | Na+/K+ ratio | ||||
|---|---|---|---|---|---|---|---|
| Seasons | S1 | S2 | S1 | S2 | S1 | S2 | |
| Rice varieties | |||||||
| Giza 177 | 4.13 a | 4.32 a | 2.41 b | 2.42 b | 1.71 a | 1.79 a | |
| Giza 178 | 2.37 b | 2.14 b | 4.11 a | 4.38 a | 0.58 b | 0.49 b | |
| F test | ** | ** | ** | ** | ** | ** | |
| NPs-Si and NPs-Se treatments | |||||||
| Control | 4.56 a | 4.33 a | 2.31 d | 2.42 d | 1.97 a | 1.79 a | |
| Soaking-NPs-Si | 3.07 c | 3.00 c | 3.31 b | 3.70 b | 0.92 c | 0.85 c | |
| Soaking-NPs-Se | 3.00 c | 3.05 c | 3.35 b | 3.62 b | 0.90 c | 0.84 c | |
| Foliar-NPs-Si at MT | 3.56 b | 3.67 b | 3.15 b | 3.15 b | 1.13 b | 1.17 b | |
| Foliar-NPs-Se at MT | 3.63 b | 3.72 b | 3.00 b | 3.21 b | 1.21 b | 1.16 b | |
| Foliar-NPs-Si at PI | 3.34 b | 3.56 b | 3.14 b | 3.16 b | 1.06 b | 1.13 b | |
| Foliar-NPs-Se at PI | 3.45 b | 3.52 b | 3.16 b | 3.07 b | 1.09 b | 1.15 b | |
| Foliar-NPs-Si at BT | 2.15 d | 2.11 d | 4.36 a | 4.25 a | 0.49 d | 0.50 d | |
| Foliar-NPs-Se at BT | 2.33 d | 2.25 d | 4.00 a | 4.10 a | 0.58 d | 0.55 d | |
| F test | ** | ** | ** | ** | ** | ** | |
| Interaction | ns | ns | ns | ns | ns | ns | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Badawy, S.A.; Zayed, B.A.; Bassiouni, S.M.A.; Mahdi, A.H.A.; Majrashi, A.; Ali, E.F.; Seleiman, M.F. Influence of Nano Silicon and Nano Selenium on Root Characters, Growth, Ion Selectivity, Yield, and Yield Components of Rice (Oryza sativa L.) under Salinity Conditions. Plants 2021, 10, 1657. https://doi.org/10.3390/plants10081657
Badawy SA, Zayed BA, Bassiouni SMA, Mahdi AHA, Majrashi A, Ali EF, Seleiman MF. Influence of Nano Silicon and Nano Selenium on Root Characters, Growth, Ion Selectivity, Yield, and Yield Components of Rice (Oryza sativa L.) under Salinity Conditions. Plants. 2021; 10(8):1657. https://doi.org/10.3390/plants10081657
Chicago/Turabian StyleBadawy, Shimaa A., Bassiouni A. Zayed, Sherif M. A. Bassiouni, Ayman H. A. Mahdi, Ali Majrashi, Esmat F. Ali, and Mahmoud F. Seleiman. 2021. "Influence of Nano Silicon and Nano Selenium on Root Characters, Growth, Ion Selectivity, Yield, and Yield Components of Rice (Oryza sativa L.) under Salinity Conditions" Plants 10, no. 8: 1657. https://doi.org/10.3390/plants10081657
APA StyleBadawy, S. A., Zayed, B. A., Bassiouni, S. M. A., Mahdi, A. H. A., Majrashi, A., Ali, E. F., & Seleiman, M. F. (2021). Influence of Nano Silicon and Nano Selenium on Root Characters, Growth, Ion Selectivity, Yield, and Yield Components of Rice (Oryza sativa L.) under Salinity Conditions. Plants, 10(8), 1657. https://doi.org/10.3390/plants10081657