
Sepal Identity of the Pappus and Floral Organ Development in the Common Dandelion (Taraxacum officinale; Asteraceae)

 , , and
, , and
Abstract
:1. Introduction
1.1. Evolution of Dispersal Capacity
1.2. Pappus Development in the Asteraceae
1.3. The ABC(D)E-Model of Floral Organ Development
1.4. Dandelion
1.5. Research Question and Aim
2. Results
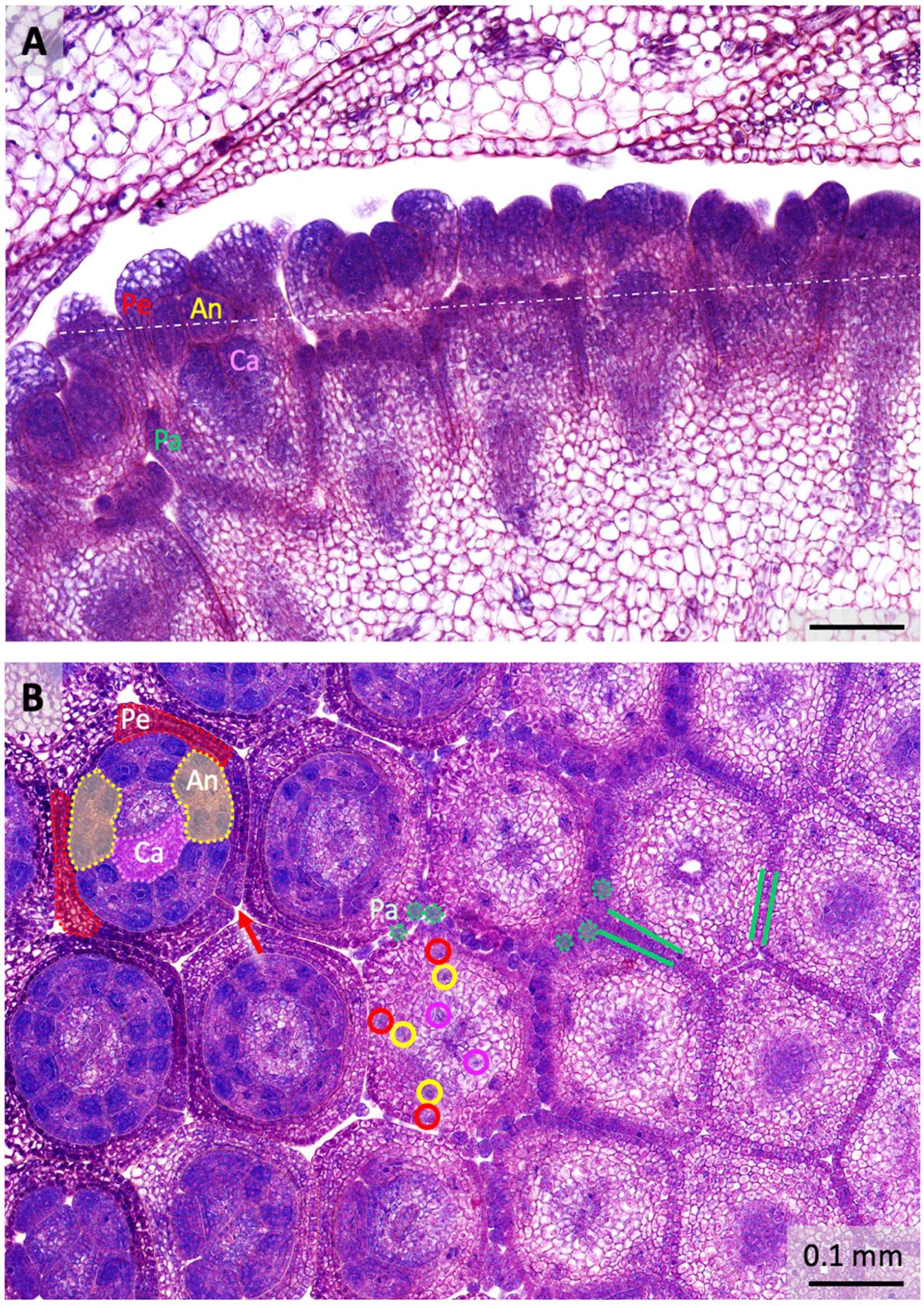
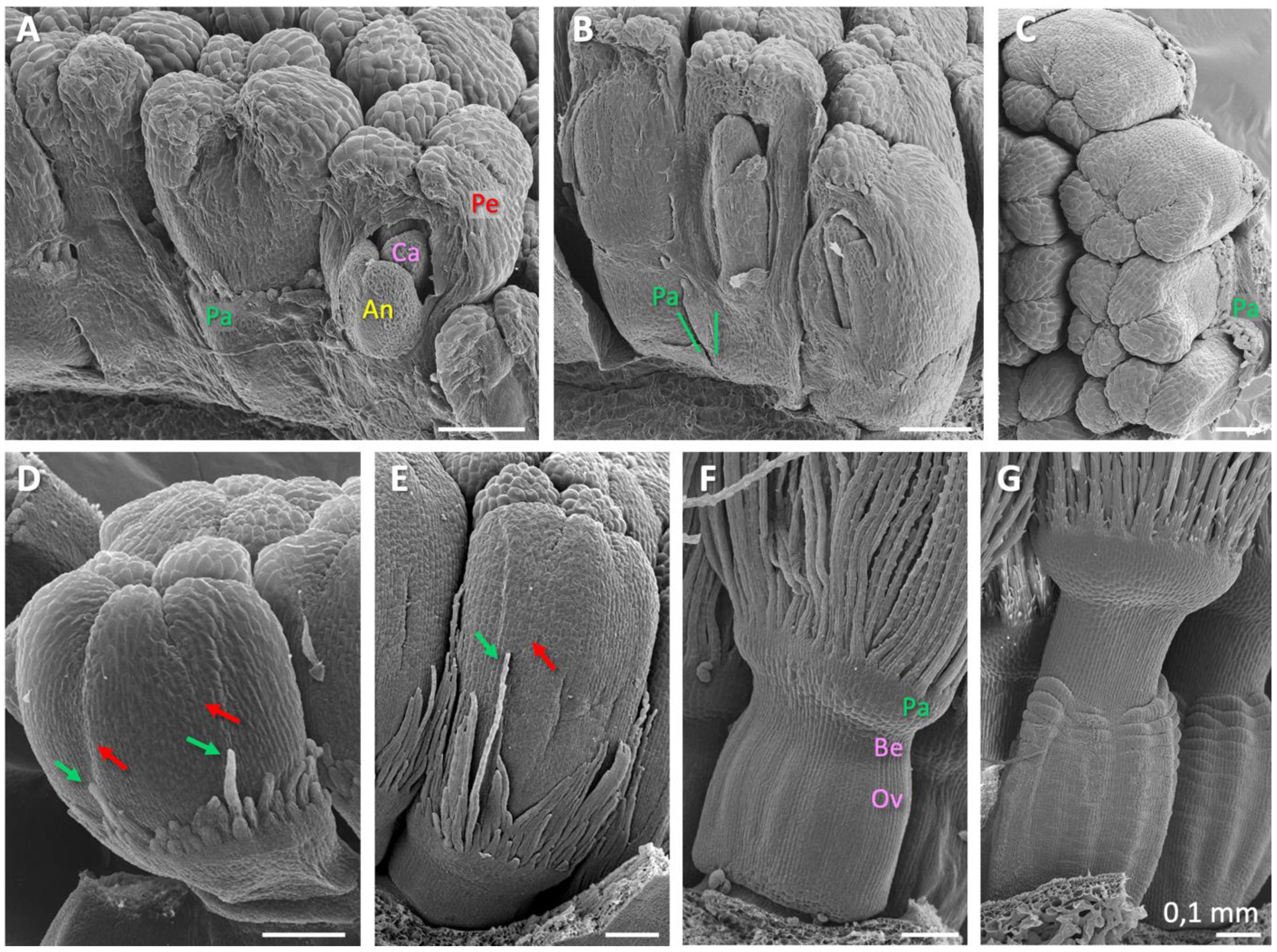
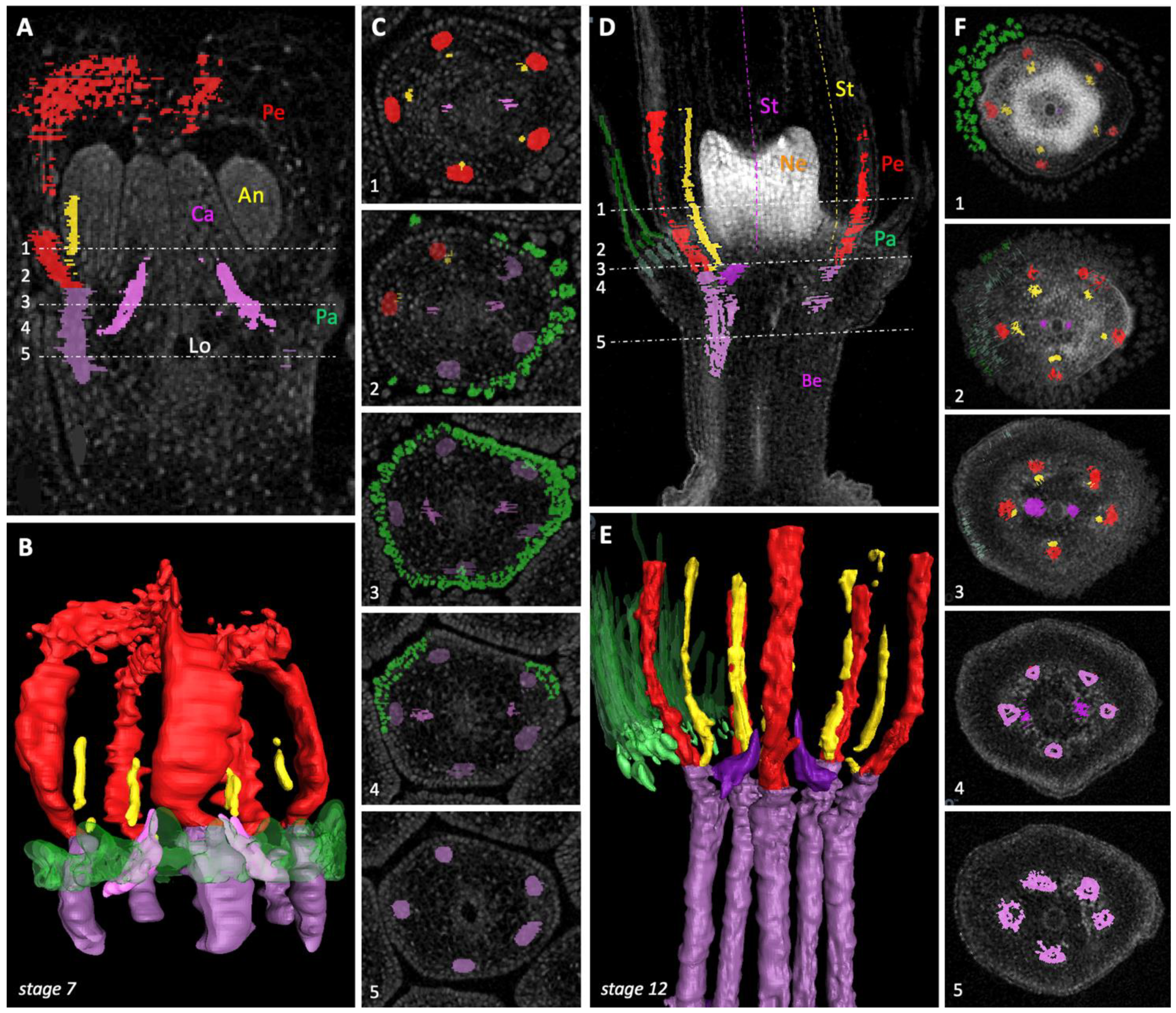
2.1. Floral Organ Ontogeny and Vasculature
2.2. Floral Organ Gene Expression
3. Discussion
3.1. Floral Organogenesis and Stage Definition
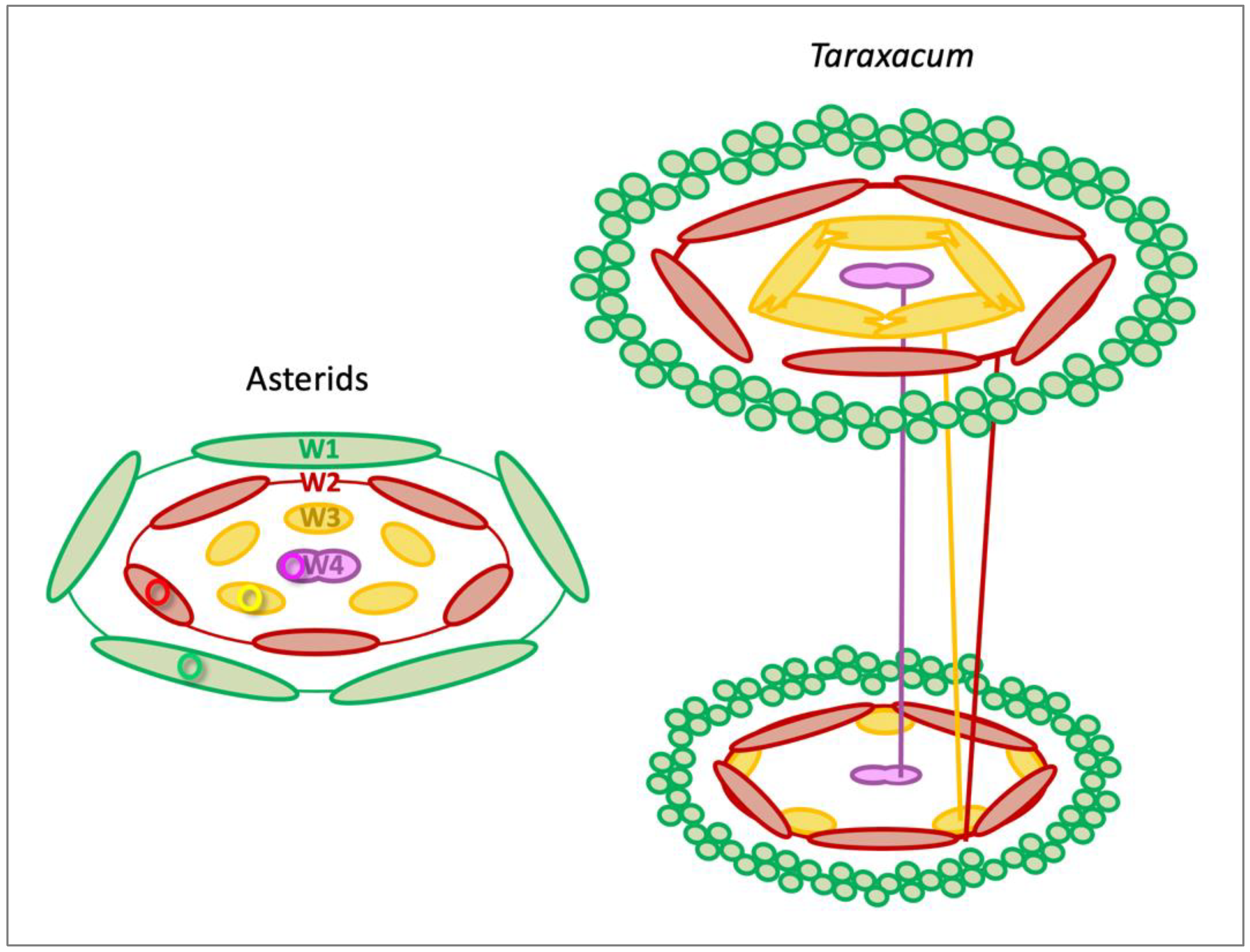
3.2. Sepaloid Origin of the Pappus
3.3. Development of the Beak and its Connection to the Pappus
3.4. Floral Vasculature
3.5. Evolution of Dispersal Capacity
4. Materials and Methods
4.1. Plant Material and Sampling of Buds for Micromorphological Analysis
4.2. Histology
4.3. Scanning Electron Microscopy (SEM)
4.4. Micro Computed Tomography (μCT) Scanning
4.5. Primer Design and Verification
4.6. DNA Isolation, RNA Isolation, Samples Used, and cDNA Preparation
4.7. qPCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cummins, C.; Seale, M.; Macente, A.; Certini, D.; Mastropaolo, E.; Viola, I.M.; Nakayama, N. A separated vortex ring underlies the flight of the dandelion. Nature 2018, 562, 414–418. [Google Scholar] [CrossRef]
- Bont, Z.; Pfander, M.; Robert, C.A.M.; Huber, M.; Poelman, E.H.; Raaijmakers, C.E.; Erb, M. Adapted dandelions trade dispersal for germination upon root herbivore attack. Proc. Biol. Sci. 2020, 287, 20192930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheldon, J.C.; Burrows, F.M. The dispersal effectiveness of the achene–pappus units of selected compositae in steady winds with convection. New Phytol. 1972, 72, 665–675. [Google Scholar] [CrossRef]
- Ledda, P.G.; Siconolfi, L.; Viola, F.; Camarri, S.; Gallaire, F. Flow dynamics of a dandelion pappus: A linear stability approach. Phys. Rev. Fluids 2019, 4, 071901. [Google Scholar] [CrossRef]
- Nathan, R.; Muller-Landau, H.C. Spatial patterns of seed dispersal, their determinants and consequences for recruitment. Trends Ecol. Evol. 2000, 15, 278–285. [Google Scholar] [CrossRef]
- Vijverberg, K.; Mes, T.H.; Bachmann, K. Chloroplast DNA evidence for the evolution of Microseris (Asteraceae) in Australia and New Zealand after long-distance dispersal from western North America. Am. J. Bot. 1999, 86, 1448–1463. [Google Scholar] [CrossRef]
- Rieseberg, L.H.; Kim, S.C.; Randell, R.A.; Whitney, K.D.; Gross, B.L.; Lexer, C.; Clay, K. Hybridization and the colonization of novel habitats by annual sunflowers. Genetica 2007, 129, 149–165. [Google Scholar] [CrossRef]
- Snell, R.S.; Beckman, N.G.; Fricke, E.; Loiselle, B.A.; Carvalho, C.S.; Jones, L.R.; Lichti, N.I.; Lustenhouwer, N.; Schreiber, S.J.; Strickland, C.; et al. Consequences of intraspecific variation in seed dispersal for plant demography, communities, evolution and global change. AoB Plants 2019, 11, plz016. [Google Scholar] [CrossRef] [Green Version]
- United Nations’ Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services (IPBES). Global Assessment Report on Biodiversity and Ecosystem Services. 2019. Available online: ipbes.net/global-assessment (accessed on 8 July 2021).
- Johnson, M.T.J.; Munshi-South, J. Evolution of life in urban environments. Science 2017, 358, eaam8327. [Google Scholar] [CrossRef]
- Cheptou, P.-O.; Carrue, O.; Rouifed, S.; Cantarel, A. Rapid evolution of seed dispersal in an urban environment in the weed Crepis sancta. Proc. Natl. Acad. Sci. USA 2008, 105, 3796–3799. [Google Scholar] [CrossRef] [Green Version]
- Palumbi, S.R. Humans as the World’s greatest evolutionary force. Science 2001, 293, 1786–1790. [Google Scholar] [CrossRef] [PubMed]
- Sih, A.; Ferrari, M.C.O.; Harris, D.J. Evolution and behavioural responses to human-induced rapid environmental change. Evol. Appl. 2011, 4, 367–387. [Google Scholar] [CrossRef]
- Jana, B.K.; Mukherjee, S.K. Pappus structure in the family compositae—A Short communication. Int. J. Sci. Res. 2014, 3, 29–30. [Google Scholar] [CrossRef]
- Panero, J.L.; Crozier, B.S. Macroevolutionary dynamics in the early diversification of Asteraceae. Mol. Phylogenet. Evol. 2016, 99, 116–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marzinek, J.; Cavalari De-Paula, O.; Oliveira, D.M.T. Cypsela or achene? Refining terminology by considering anatomical and historical factors. Braz. J. Bot. 2008, 31, 549–553. [Google Scholar] [CrossRef] [Green Version]
- Stuessy, T.F.; Garver, D. The defensive role of pappus in heads of Compositae. In Compositae: Biology & Utilization, Proceedings of International Compositae Conference Kew 1994; Caligari, P.D.S., Hind, D.J.N., Eds.; Royal Botanic Gardens: Kew, UK, 1996; Volume 2, pp. 81–91. [Google Scholar]
- Hale, A.N.; Imfeld, S.M.; Hart, C.E.; Gribbins, K.M.; Yoder, J.A.; Collier, M.H. Reduced seed germination after pappus removal in the North American dandelion (Taraxacum officinale; Asteraceae). Weed Sci. 2010, 58, 420–425. [Google Scholar] [CrossRef]
- Mandel, J.R.; Dikow, R.B.; Siniscalchi, C.M.; Thapa, R.; Watson, L.E.; Funk, V.A. A fully resolved backbone phylogeny reveals numerous dispersals and explosive diversifications throughout the history of Asteraceae. Proc. Natl. Acad. Sci. USA 2019, 116, 14083–14088. [Google Scholar] [CrossRef] [Green Version]
- The Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Stevens, P.F. Angiosperm Phylogeny Website, Version 14. July 2017. 2001 Onwards (and More or Less Continuously Updated Since). Available online: mobot.org/MOBOT/research/APweb (accessed on 8 July 2021).
- Elomaa, P.; Zhao, Y.; Zhang, T. Flower heads in Asteraceae—Recruitment of conserved developmental regulators to control the flower-like inflorescence architecture. Hortic. Res. 2018, 5. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Cieslak, M.; Owens, A.; Wang, F.; Broholm, S.K.; Teeri, T.H.; Elomaa, P.; Prusinkiewicz, P. Phyllotactic patterning of gerbera flower heads. Proc. Natl. Acad. Sci. USA 2021, 118, e2016304118. [Google Scholar] [CrossRef]
- Anderberg, A.A.; Baldwin, B.G.; Bayer, R.G.; Breitwieser, J.; Jeffrey, C.; Dillon, M.O.; Eldenäs, P.; Funk, V.; Garcia-Jacas, N.; Hind, D.J.N.; et al. Compositae. In Flowering Plants, Eudicots: Asterales. the Families and Genera of Vascular Plants; Kubitzki, K., Kadereit, J.W., Jeffrey, C., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; Volume VIII. [Google Scholar]
- Smyth, D.R. Evolution and genetic control of the floral ground plan. New Phytol. 2018, 220, 70–86. [Google Scholar] [CrossRef] [Green Version]
- Carlquist, S. Tribal interrelationships and phylogeny of the asteraceae. Aliso 1973, 8, 465–492. [Google Scholar] [CrossRef] [Green Version]
- Sattler, R.; Rutishauser, E. The fundamental relevance of morphology and morphogenesis to plant research. Ann. Bot. 1997, 80, 571–582. [Google Scholar] [CrossRef] [Green Version]
- Small, J. The origin and development of the Compositae. V. The pappus. New Phytol. Reprint 1919, 11, 98–123. [Google Scholar]
- Sattler, R. Organogenesis of Flowers. A Photographic Text-Atlas; University of Toronto Press: Toronto, ON, Canada; Buffalo, NY, USA, 1973. [Google Scholar]
- Ramiah, N.; Sayeeduddin, M. Homology of the pappus in the light of trichome distribution. Curr. Sci. 1958, 27, 402–404. [Google Scholar]
- Coen, E.S.; Meyerowitz, E.M. The war of the whorls: Genetic interactions controlling flower development. Nature 1991, 353, 31–37. [Google Scholar] [CrossRef]
- Thomson, B.; Wellmer, F. Molecular regulation of flower development. Curr. Top. Dev. Biol. 2019, 131, 185–210. [Google Scholar] [CrossRef]
- Smaczniak, C.; Immink, R.G.; Angenent, G.C.; Kaufmann, K. Developmental and evolutionary diversity of plant MADS-domain factors: Insights from recent studies. Development 2012, 139, 3081–3098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theißen, G.; Melzer, R.; Rümpler, F. MADS-domain transcription factors and the floral quartet model of flower development: Linking plant development and evolution. Development 2016, 143, 3259–3271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rijpkema, A.S.; Vandenbussche, M.; Koes, R.; Heijmans, K.; Gerats, T. Variations on a theme: Changes in the floral ABCs in angiosperms. Semin. Cell Dev. Biol. 2010, 21, 100–107. [Google Scholar] [CrossRef]
- Mandel, M.A.; Bowman, J.L.; Kempin, S.A.; Ma, H.; Meyerowitz, E.M.; Yanofsky, M.F. Manipulation of flower structure in transgenic tobacco. Cell 1992, 71, 133–143. [Google Scholar] [CrossRef]
- Jofuku, K.D.; den Boer, B.G.; Van Montagu, M.; Okamuro, J.K. Control of Arabidopsis flower and seed development by the homeotic gene APETALA2. Plant Cell 1994, 6, 1211–1225. [Google Scholar] [CrossRef] [Green Version]
- Huijser, P.; Klein, J.; Lönnig, W.E.; Meijer, H.; Saedler, H.; Sommer, H. Bracteomania, an inflorescence anomaly, is caused by the loss of function of the MADS-box gene squamosa in Antirrhinum majus. EMBO J. 1992, 11, 1239–1249. [Google Scholar] [CrossRef]
- Keck, E.; McSteen, P.; Carpenter, R.; Coen, E. Separation of genetic functions controlling organ identity in flowers. EMBO J. 2003, 22, 1058–1066. [Google Scholar] [CrossRef]
- Cartolano, M.; Castillo, R.; Efremova, N.; Kuckenberg, M.; Zethof, J.; Gerats, T.; Schwarz-Sommer, Z.; Vandenbussche, M. A conserved microRNA module exerts homeotic control over Petunia hybrida and Antirrhinum majus floral organ identity. Nat. Genet. 2007, 39, 901–905. [Google Scholar] [CrossRef] [Green Version]
- Morel, P.; Heijmans, K.; Ament, K.; Chopy, M.; Trehin, C.; Chambrier, P.; Rodrigues Bento, S.; Bimbo, A.; Vandenbussche, M. The floral C-lineage genes trigger nectary development in Petunia and Arabidopsis. Plant Cell 2018, 30, 2020–2037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tröbner, W.; Ramirez, L.; Motte, P.; Hue, I.; Huijser, P.; Lönnig, W.E.; Saedler, H.; Sommer, H.; Schwarz-Sommer, Z. GLOBOSA: A homeotic gene which interacts with DEFICIENS in the control of Antirrhinum floral organogenesis. EMBO J. 1992, 11, 4693–4704. [Google Scholar] [CrossRef] [PubMed]
- Vandenbussche, M.; Zethof, J.; Royaert, S.; Weterings, K.; Gerats, T. The duplicated B-class heterodimer model: Whorl-specific effects and complex genetic interactions in Petunia hybrida flower development. Plant Cell 2004, 16, 741–754. [Google Scholar] [CrossRef] [Green Version]
- Rijpkema, A.S.; Royaert, S.; Zethof, J.; van der Weerden, G.; Gerats, T.; Vandenbussche, M. Analysis of the Petunia TM6 MADS box gene reveals functional divergence within the DEF/AP3 lineage. Plant Cell 2006, 18, 1819–1832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teeri, T.H.; Uimari, A.; Kotilainen, M.; Laitinen, R.; Help, H.; Elomaa, P.; Albert, V.A. Reproductive meristem fates in Gerbera. J. Exp. Bot. 2006, 57, 3445–3455. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.; Kotilainen, M.; Pölllanen, E.; Mehto, M.; Elomaa, P.; Helariutta, Y.; Albert, V.A.; Teeri, T.H. Organ identity genes and modified patterns of flower development in Gerbera hybrida (Asteraceae). Plant J. 1999, 17, 51–62. [Google Scholar] [CrossRef]
- Broholm, S.K.; Pöllänen, E.; Ruokolainen, S.; Tähtiharju, S.; Kotilainen, M.; Albert, V.A.; Elomaa, P.; Teeri, T.H. Functional characterization of B class MADS-box transcription factors in Gerbera hybrida. J. Exp. Bot. 2010, 61, 75–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; Zhao, Y.; Juntheikki, I.; Mouhu, K.; Broholm, S.K.; Rijpkema, A.S.; Kins, L.; Lan, T.; Albert, V.A.; Teeri, T.H.; et al. Dissecting functions of SEPALLATA-like MADS box genes in patterning of the pseudanthial inflorescence of Gerbera hybrida. New Phytol. 2017, 216, 939–954. [Google Scholar] [CrossRef] [Green Version]
- Ruokolainen, S.; Ng, Y.P.; Broholm, S.K.; Albert, V.A.; Elomaa, P.; Teeri, T.H. Characterization of SQUAMOSA-like genes in Gerbera hybrida, including one involved in reproductive transition. BMC Plant Biol. 2010, 10, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badouin, H.; Gouzy, J.; Grassa, C.J.; Murat, F.; Staton, S.E.; Cottret, L.; Langlade, N.B. The sunflower genome provides insights into oil metabolism, flowering and Asterid evolution. Nature 2017, 546, 148–152. [Google Scholar] [CrossRef]
- Shchennikova, A.V.; Shulga, O.A.; Skryabin, K.G. Diversification of the homeotic AP3 clade MADS-Box genes in Asteraceae species Chrysanthemum morifolium L. and Helianthus annuus L. Biochem. Biophys. Mol. Biol. 2019, 483, 348–354. [Google Scholar] [CrossRef]
- Reyes-Chin-Wo, S.; Wang, Z.; Yang, X.; Kozik, A.; Arikit, S.; Song, C.; Xia, L.; Froenicke, L.; Lavelle, D.O.; Truco, M.J.; et al. Genome assembly with in vitro proximity ligation data and whole-genome triplication in lettuce. Nat. Commun. 2017, 8, 1–11. [Google Scholar] [CrossRef]
- Ning, K.; Han, Y.; Chen, Z.; Luo, C.; Wang, S.; Zhang, W.; Li, L.; Zhang, X.; Fan, S.; Wang, Q. Genome-wide analysis of MADS-box family genes during flower development in lettuce. Plant Cell Environ. 2019, 42, 1868–1881. [Google Scholar] [CrossRef]
- Plants of the World Online (POWO). Royal Botanic Gardens Kew. 2019. Available online: plantsoftheworldonline.org/?q=Taraxacum (accessed on 8 July 2021).
- Sterk, A.A.; Hommels, C.H.; Jenniskens, M.J.P.J.; Neuteboom, J.H.; Den Nijs, J.C.M.; Oosterveld, P.; Segal, S. Paardebloemen, Planten Zonder Vader; Koninklijke Nederlandse Natuurhistorische Vereniging: Zeist, The Netherlands, 1987. [Google Scholar]
- Richards, A.J. The origin of Taraxacum agamospecies. Bot. J. Linn. Soc. 1973, 66, 189–211. [Google Scholar] [CrossRef]
- Van Dijk, P.; De Jong, H.; Vijverberg, K.; Biere, A. An apomixis-gene’s view on dandelions. In Lost Sex, the Evolutionary Biology of Parthenogenesis; Schön, I., Martens, K., Van Dijk, P., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 475–493. [Google Scholar]
- Kirschner, J.; Záveská-Drábková, L.; Štěpánek, J.; Uhlemann, I. Towards a better understanding of the Taraxacum evolution (Compositae-Cichorieae) on the basis of nrDNA of sexually reproducing species. Plant Syst. Evol. 2015, 301, 1135–1156. [Google Scholar] [CrossRef]
- Taraxacum Nederland, 2017–2021. Available online: taraxacumnederland.nl (accessed on 8 July 2021).
- Anderson, L. Floral anatomy of Chrysothamnus (Astereae, Compositae). SIDA Contrib. Bot. 1970, 3, 466–503. [Google Scholar]
- Sgamma, T.; Pape, J.; Massiah, A.; Jackson, S. Selection of reference genes for diurnal and developmental time-course real-time PCR expression analyses in lettuce. Plant Methods 2016, 12, 21. [Google Scholar] [CrossRef] [Green Version]
- Lin, T.; Xu, X. Genome analysis of Taraxacum kok-saghyz Rodin provides new insights into rubber biosynthesis. Nat. Sci. Rev. 2018, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Smyth, D.R.; Bowman, J.L.; Meyerowitz, E.M. Early flower development in Arabidopsis. Plant Cell 1990, 2, 755–767. [Google Scholar] [CrossRef] [Green Version]
- Helariutta, Y.; Elomaa, P.; Kotilainen, M.; Seppänen, P.; Teeri, T.H. Cloning of cDNA coding for dihydroflavonol-4-reductase (DFR) and characterization of dfr expression in the corollas of Gerbera hybrida var. Regina (Compositae). Plant Mol. Biol. 1993, 22, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Laitinen, R.A.; Broholm, S.; Albert, V.A.; Teeri, T.H.; Elomaa, P. Patterns of MADS-box gene expression mark flower-type development in Gerbera hybrida (Asteraceae). BMC Plant Biol. 2006, 6, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pu, Y.; Huang, H.; Wen, X.; Lu, C.; Zhang, B.; Gu, X.; Qi, S.; Fan, G.; Wang, W.; Dai, S. Comprehensive transcriptomic analysis provides new insights into the mechanism of ray floret morphogenesis in chrysanthemum. Genomics 2020, 21, 728. [Google Scholar] [CrossRef] [PubMed]
- Schuchovski, C.; Meulia, T.; Sant’Anna-Santos, B.F.; Fresnedo-Ramírez, J. Inflorescence development and floral organogenesis in Taraxacum kok-saghyz. J. Plants 2020, 9, 1258. [Google Scholar] [CrossRef]
- Harris, E. Inflorescence and floral ontogeny in Asteraceae: A synthesis of historical and current concepts. Bot. Rev. 1995, 61, 93–278. [Google Scholar] [CrossRef]
- Singh, B.; Agarwal, A. Floral anatomy of Actinomeris squarrosa and Bidens biternata (Asteraceae). Acta Bot. Hung. 2001, 43, 423–432. [Google Scholar] [CrossRef]
- Endress, P.K. The whole and the parts: Relationships between floral architecture and floral organ shape, and their repercussions on the interpretation of fragmentary floral fossils. Ann. Missouri Bot. Gard. 2008, 95, 101–120. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Shen, C.; Meng, P.; Tan, G.; Lv, L. Analysis and review of trichomes in plants. BMC Plant Biol. 2021, 21, 1–11. [Google Scholar] [CrossRef]
- De Craene, L.D. Meristic changes in flowering plants: How flowers play with numbers. Flora 2016, 221, 22–37. [Google Scholar] [CrossRef]
- Douglas, G.E. The inferior ovary. Bot. Rev. 1957, 23, 1–45. [Google Scholar] [CrossRef]
- Soltis, D.E.; Fishbein, M.; Kuzoff, R.K. Reevaluating the evolution of epigyny: Data from phylogenetics and floral ontogeny. Int. J. Plant Sci. 2003, 164, S251–S264. [Google Scholar] [CrossRef]
- Bachmann, K.; Chambers, K.L. Pappus part number in annual species of Microseris (Compositae, Cichoriaceae). Plant Syst. Evol. 1978, 129, 119–134. [Google Scholar] [CrossRef]
- Frangiote-Pallone, S.; De Souza, L.A. Pappus and cypsela ontogeny in Asteraceae: Structural considerations of the tribal category. Rev. Mex. Biodivers. 2014, 85, 62–77. [Google Scholar] [CrossRef] [Green Version]
- Eames, A.J. The vascular anatomy of the flower with refutation of the theory of carpel polymorphism. Am. J. Bot. 1931, 18, 147–188. [Google Scholar] [CrossRef]
- Gustafsson, M.H.G.; Bremer, K. Morphology and phylogenetic interrelationships of the Asteraceae, Calyceraceae, Campanulaceae, Goodeniaceae, and related families (Asterales). Am. J. Bot. 1995, 82, 250–265. [Google Scholar] [CrossRef]
- Verhoeven, K.J.F.; Biere, A. Geographic parthenogenesis and plant-enemy interactions in the common dandelion. BMC Evol. Biol. 2013, 13, 23. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Sun, T.; Xie, L.; Hayashi, T.; Kawabata, S.; Li, Y. Relationship between the velvet-like texture of flower petals and light reflection from epidermal cell surfaces. J. Plant Res. 2015, 128, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Yeung, E.C.; Chan, C.K.W. Glycol methacrylate: The art of embedding and serial sectioning. Botany 2015, 93, 1–8. [Google Scholar] [CrossRef]
- D’Amico, F. A polychromatic staining method for epoxy embedded tissue: A new combination of methylene blue and basic fuchsin for light microscopy. Biotech. Histochem. 2005, 80, 207–210. [Google Scholar] [CrossRef]
- Etzold, H. Eine kontrastreiche, simultane Mehrfachfärbung für pflanzenanatomische Präparate: Fuchsin-Safranin-Astrablau. Mikrokosmos 1983, 72, 213–219. [Google Scholar]
- Pramanik, D.; Dorst, N.; Meesters, N.; Spaans, M.; Smets, E.; Welten, M.; Gravendeel, B. Evolution and development of three highly specialized floral structures of bee-pollinated Phalaenopsis species. Evodevo 2020, 11, 16. [Google Scholar] [CrossRef] [PubMed]
- Vijverberg, K.; Milanovic-Ivanovic, S.; Bakx-Schotman, T.; Van Dijk, P.J. Genetic fine-mapping of DIPLOSPOROUS in Taraxacum (dandelion; Asteraceae) indicates a duplicated DIP gene. BMC Plant Biol. 2010, 10, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogstad, S.H. Saturated NaCl-CTAB solution as a means of field preservation of leaves for DNA analyses. Taxon 1992, 41, 701–708. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage | Stem (cm) * | Flower (mm) * | Pappus (mm) * | Beak (mm) * | Ovary (mm) * | Description | Figures Number | Samples Figure 7 | |
|---|---|---|---|---|---|---|---|---|---|
| Floral organ initiation | 1 | <0.1 | <0.1 | <0.1 | flower primordia: domed (round) staged | ||||
| 2 | <0.1 | <0.1 | <0.1 | corolla primordia: ring/5 lobs | 1B | FS0 | |||
| 3 | <0.1 | 0.1–0.2 | <0.1 | stamen primordia: 5 lobs; pappus primordia ring | FS0 | ||||
| 4 | <0.1 | 0.2 | 0.01 | <0.1 | carpels initiated; organs elongating | 2A; S1A | |||
| 5 | <0.1 | 0.3 | 0.05 | <0.1 | petals closed at top; carpels lower than anthers; pappus longer at petal fusion zones | 2B, 4A,D; S1B | |||
| 6 | 0.1 | 0.4 | 0.1 | <0.1 | floral part elongating; carpels passed anthers | 4B,C | |||
| Floral maturation | 7 | 0.2 | 0.6 | 0.3 | <0.1 | 0.1 | ovary, beak and flower bottom pronouncing; pappus half corolla length; nectaries initiating | 1C, 4E, 5A–C; S5A, SV1 | |
| 8 | 0.5 | 2 | 1 | 0.05 | 0.2 | ovary, beak, flower bottom, nectaries defined; vertical lines on fruit coat; scales on pappus | 4F; S2 | ||
| 9 | 1–2 | 5 | 3 | 0.1 | 0.3 | upper floral part quickly elongating; pigmentation of petals initiated | 1D | ||
| 10 | 3–6 | 8 | 5 | 0.2 | 0.5 | upper floral part quickly elongating; horizontal lines on fruit coat | 4G | F1, S1 | |
| 11 | 7–12 | 10 | 7 | 0.4 | 1 | pigmentation of petals finalised; petals still closed at top; bud closed to loosen | 1E | F1, S1 | |
| 12 | 13–20 | 17 | 8 | 0.7 | 1.5 | anthesis: bud opens (D1); corolla’s opening; pollen mature; style enlarging; stigma closed | 1F, 5D–F; S3,4,5B, SV2 | F2, S2 | |
| Seed formation | 13 | 21–30 | 25 | 9 | 1.5 | 2 | also central florets open (D2,3–5); stigma lobes opening and receptive; ovary part elongating | 1G | F2, S2 |
| 14 | >25 | old | 10 | 3 | 2.5 | 3 DAP: flower organs blown; beak and ovary elongating | F3, S3 | ||
| 15 | >25 | blown | 10 | 6 | 3 | 7 DAP: flower organs gone; beak elongating; fruit browning | F7, S7 | ||
| 16 | >25 | blown | 11 | 10 | 3.5 | 12 DAP: mature brown seed; long beak; pappus opening | 1G-insert | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vijverberg, K.; Welten, M.; Kraaij, M.; van Heuven, B.J.; Smets, E.; Gravendeel, B. Sepal Identity of the Pappus and Floral Organ Development in the Common Dandelion (Taraxacum officinale; Asteraceae). Plants 2021, 10, 1682. https://doi.org/10.3390/plants10081682
Vijverberg K, Welten M, Kraaij M, van Heuven BJ, Smets E, Gravendeel B. Sepal Identity of the Pappus and Floral Organ Development in the Common Dandelion (Taraxacum officinale; Asteraceae). Plants. 2021; 10(8):1682. https://doi.org/10.3390/plants10081682
Chicago/Turabian StyleVijverberg, Kitty, Monique Welten, Marjan Kraaij, Bertie Joan van Heuven, Erik Smets, and Barbara Gravendeel. 2021. "Sepal Identity of the Pappus and Floral Organ Development in the Common Dandelion (Taraxacum officinale; Asteraceae)" Plants 10, no. 8: 1682. https://doi.org/10.3390/plants10081682
APA StyleVijverberg, K., Welten, M., Kraaij, M., van Heuven, B. J., Smets, E., & Gravendeel, B. (2021). Sepal Identity of the Pappus and Floral Organ Development in the Common Dandelion (Taraxacum officinale; Asteraceae). Plants, 10(8), 1682. https://doi.org/10.3390/plants10081682