
Biosynthetic Pathway of Proanthocyanidins in Major Cash Crops


Abstract
:1. Introduction
2. Biosynthetic Pathways of PAs and Anthocyanins
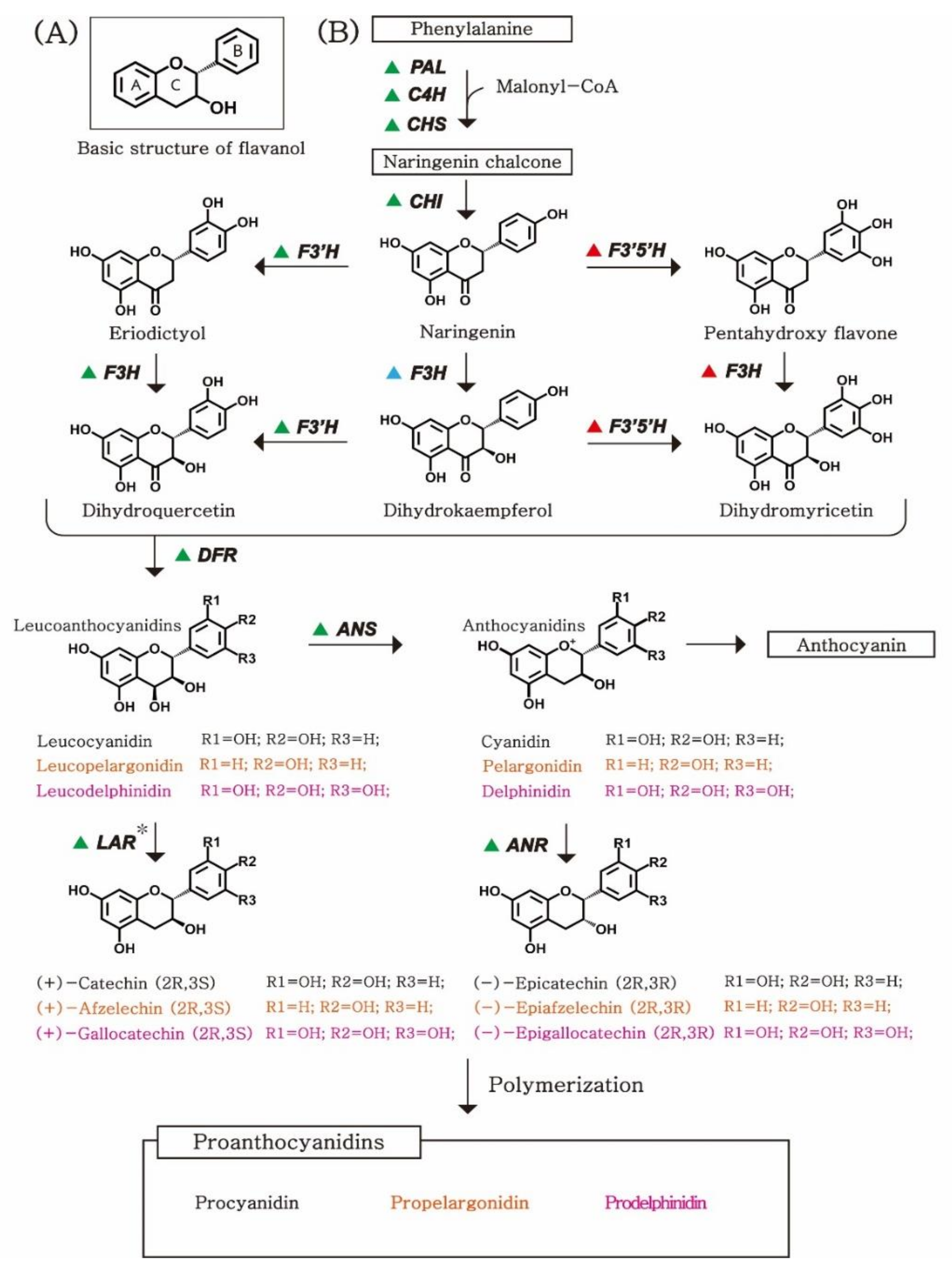
2.1. Chemical Structure of PAs
2.2. Biosynthetic Pathway of PAs
2.3. Regulatory Mechanisms of Flavonoids in Model Plants
2.4. Major Cash Crops with High PA Contents
2.5. Identification of Orthologous Genes Involved in PA Biosynthesis in Major Cash Crops
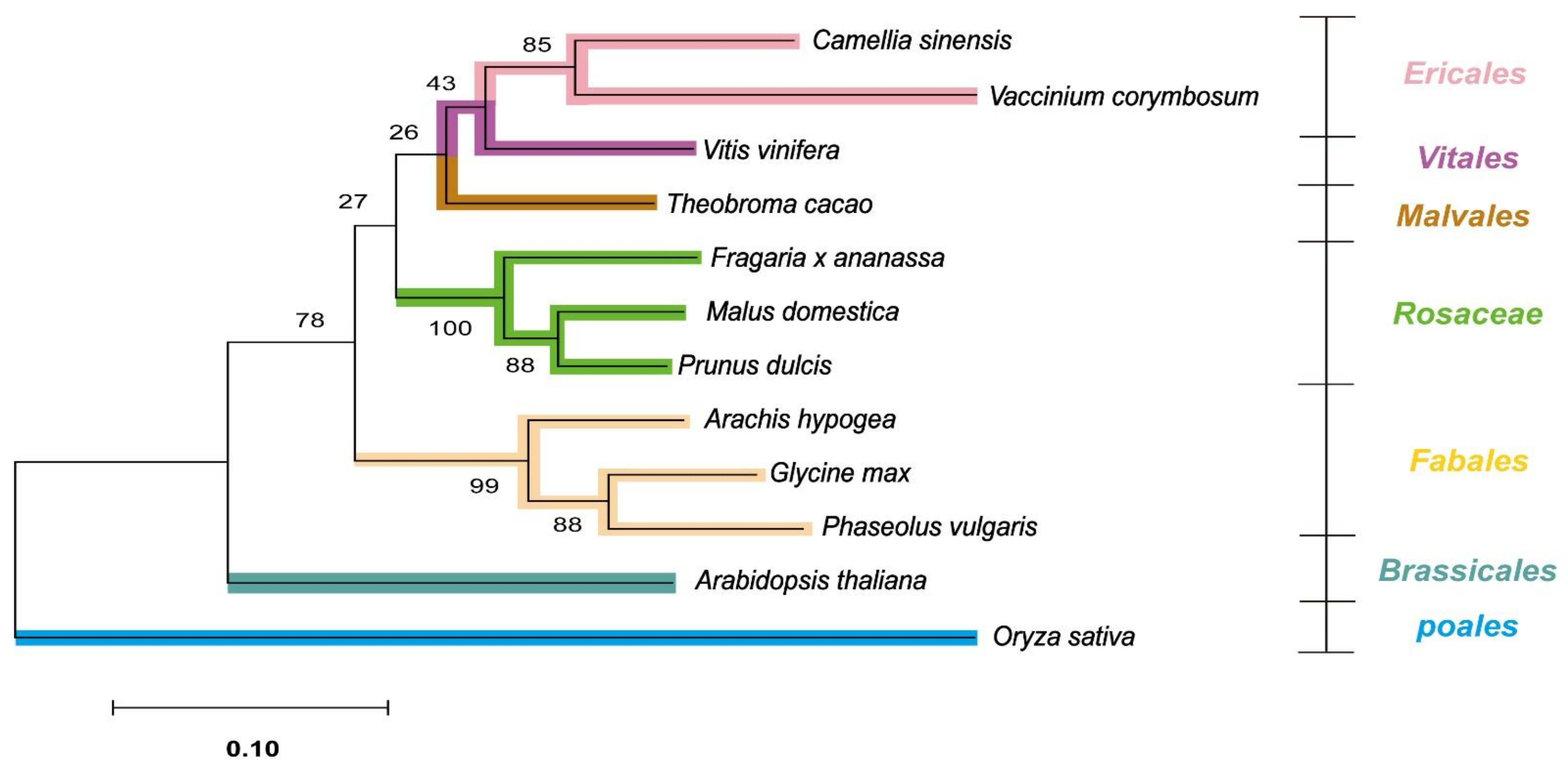
2.6. Phylogenetic Analysis of Orthologs
3. Regulatory Mechanisms for the Biosynthesis of PAs and Anthocyanins
3.1. Competitions between Parallel Pathways in the Flavonoid Pathway
3.2. Species-Specific Pathways for the Biosynthesis of Catechin and Epicatechin
4. Conclusions
5. Materials and Methods
5.1. Sequence Analysis
5.2. Phylogenetic Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lepiniec, L.; Debeaujon, I.; Routaboul, J.-M.; Baudry, A.; Pourcel, L.; Nesi, N.; Caboche, M. Genetics and Biochemistry of Seed Flavonoids. Annu. Rev. Plant. Biol. 2006, 57, 405–430. [Google Scholar] [CrossRef]
- Hellström, J.K.; Törrönen, A.R.; Mattila, P.H. Proanthocyanidins in Common Food Products of Plant Origin. J. Agric. Food Chem. 2009, 57, 7899–7906. [Google Scholar] [CrossRef]
- Dixon, R.A.; Xie, D.; Sharma, S.B. Proanthocyanidins—A Final Frontier in Flavonoid Research? New Phytol. 2005, 165, 9–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, D.-Y.; Dixon, R.A. Proanthocyanidin Biosynthesis–Still More Questions than Answers? Phytochemistry 2005, 66, 2127–2144. [Google Scholar] [CrossRef] [Green Version]
- An, J.-P.; Li, R.; Qu, F.-J.; You, C.-X.; Wang, X.-F.; Hao, Y.-J. R2R3-MYB Transcription Factor MdMYB23 Is Involved in the Cold Tolerance and Proanthocyanidin Accumulation in Apple. Plant J. 2018, 96, 562–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malisch, C.S.; Salminen, J.-P.; Kölliker, R.; Engström, M.T.; Suter, D.; Studer, B.; Lüscher, A. Drought Effects on Proanthocyanidins in Sainfoin (Onobrychis Viciifolia Scop.) Are Dependent on the Plant’s Ontogenetic Stage. J. Agric. Food Chem. 2016, 64, 9307–9316. [Google Scholar] [CrossRef]
- Mellway, R.D.; Tran, L.T.; Prouse, M.B.; Campbell, M.M.; Constabel, C.P. The Wound-, Pathogen-, and Ultraviolet B-Responsive MYB134 Gene Encodes an R2R3 MYB Transcription Factor That Regulates Proanthocyanidin Synthesis in Poplar. Plant Physiol. 2009, 150, 924–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, D.; Huang, T.; Tian, B.; Zhan, J. Advances in Biosynthesis and Biological Functions of Proanthocyanidins in Horticultural Plants. Foods 2020, 9, 1774. [Google Scholar] [CrossRef]
- Ullah, C.; Unsicker, S.B.; Fellenberg, C.; Constabel, C.P.; Schmidt, A.; Gershenzon, J.; Hammerbacher, A. Flavan-3-Ols Are an Effective Chemical Defense against Rust Infection. Plant Physiol. 2017, 175, 1560–1578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, L.; Wang, L.; Han, Z.; Jiang, Y.; Zhao, L.; Liu, H.; Yang, L.; Luo, K. Molecular Cloning and Characterization of PtrLAR3, a Gene Encoding Leucoanthocyanidin Reductase from Populus Trichocarpa, and Its Constitutive Expression Enhances Fungal Resistance in Transgenic Plants. J. Exp. Bot. 2012, 63, 2513–2524. [Google Scholar] [CrossRef] [PubMed]
- Santos-Buelga, C.; Scalbert, A. Proanthocyanidins and Tannin-like Compounds–Nature, Occurrence, Dietary Intake and Effects on Nutrition and Health. J. Sci. Food Agric. 2000, 80, 1094–1117. [Google Scholar] [CrossRef]
- Engelhardt, U.H.; Lakenbrink, C.; Pokorny, O. Proanthocyanidins, Bisflavanols, and Hydrolyzable Tannins in Green and Black Teas. In Nutraceutical Beverages; (ACS Symposium Series); American Chemical Society: Washington, DC, USA, 2003; Volume 871, pp. 254–264. ISBN 978-0-8412-3823-7. [Google Scholar]
- Schroeter, H.; Heiss, C.; Balzer, J.; Kleinbongard, P.; Keen, C.L.; Hollenberg, N.K.; Sies, H.; Kwik-Uribe, C.; Schmitz, H.H.; Kelm, M. (–)-Epicatechin Mediates Beneficial Effects of Flavanol-Rich Cocoa on Vascular Function in Humans. Proc. Natl. Acad. Sci. USA 2006, 103, 1024–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prior, R.L.; Gu, L. Occurrence and Biological Significance of Proanthocyanidins in the American Diet. Phytochemistry 2005, 66, 2264–2280. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Yang, B.; Wang, J.; Liu, Y.; Yu, L.; Jiang, Y. Immunomodulatory and Anticancer Activities of Flavonoids Extracted from Litchi (Litchi Chinensis Sonn.) Pericarp. Int. Immunopharmacol. 2007, 7, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Rauf, A.; Imran, M.; Abu-Izneid, T.; Patel, S.; Pan, X.; Naz, S.; Sanches Silva, A.; Saeed, F.; Rasul Suleria, H.A. Proanthocyanidins: A Comprehensive Review. Biomed. Pharmacother. 2019, 116, 108999. [Google Scholar] [CrossRef]
- Lin, G.-M.; Lin, H.-Y.; Hsu, C.-Y.; Chang, S.-T. Structural Characterization and Bioactivity of Proanthocyanidins from Indigenous Cinnamon (Cinnamomum Osmophloeum). J. Sci. Food Agric. 2016, 96, 4749–4759. [Google Scholar] [CrossRef]
- Ha, J.; Kim, M.; Kim, M.Y.; Lee, T.; Yoon, M.Y.; Lee, J.; Lee, Y.-H.; Kang, Y.-G.; Park, J.S.; Lee, J.H.; et al. Transcriptomic Variation in Proanthocyanidin Biosynthesis Pathway Genes in Soybean (Glycine Spp.). J. Sci. Food Agric. 2018, 98, 2138–2146. [Google Scholar] [CrossRef]
- Rodríguez-Pérez, C.; García-Villanova, B.; Guerra-Hernández, E.; Verardo, V. Grape Seeds Proanthocyanidins: An Overview of In Vivo Bioactivity in Animal Models. Nutrients 2019, 11, 2435. [Google Scholar] [CrossRef] [Green Version]
- Buzzini, P.; Turchetti, B.; Ieri, F.; Goretti, M.; Branda, E.; Mulinacci, N.; Romani, A. Catechins and Proanthocyanidins: Naturally Occurring O-Heterocycleswith Antimicrobial Activity. In Bioactive Heterocycles IV; Khan, M.T.H., Ed.; Topics in Heterocyclic Chemistry; Springer: Berlin/Heidelber, Germany, 2007; pp. 239–263. ISBN 978-3-540-73404-8. [Google Scholar]
- Gu, L.; Kelm, M.A.; Hammerstone, J.F.; Beecher, G.; Holden, J.; Haytowitz, D.; Prior, R.L. Screening of Foods Containing Proanthocyanidins and Their Structural Characterization Using LC-MS/MS and Thiolytic Degradation. J. Agric. Food Chem. 2003, 51, 7513–7521. [Google Scholar] [CrossRef]
- Gu, L.; Kelm, M.A.; Hammerstone, J.F.; Beecher, G.; Holden, J.; Haytowitz, D.; Gebhardt, S.; Prior, R.L. Concentrations of Proanthocyanidins in Common Foods and Estimations of Normal Consumption. J. Nutr. 2004, 134, 613–617. [Google Scholar] [CrossRef]
- Kimura, H.; Ogawa, S.; Akihiro, T.; Yokota, K. Structural Analysis of A-Type or B-Type Highly Polymeric Proanthocyanidins by Thiolytic Degradation and the Implication in Their Inhibitory Effects on Pancreatic Lipase. J. Chromatogr. A 2011, 1218, 7704–7712. [Google Scholar] [CrossRef]
- Forrest, G.I.; Bendall, D.S. The Distribution of Polyphenols in the Tea Plant (Camellia Sinensis L.). Biochem. J. 1969, 113, 741–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huo, Q.; Kong, X.; Yang, X.; Wang, Y.; Ma, L.; Luo, M.; Xu, D. Molecular Modification of Proanthocyanidins. Bioengineered 2016, 7, 274–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alianza SIDALC. Available online: http://www.sidalc.net/cgi-bin/wxis.exe/?IsisScript=COLPOS.xis&method=post&formato=2&cantidad=1&expresion=mfn=039519 (accessed on 23 March 2021).
- Foo, L.Y.; Porter, L.J. The Phytochemistry of Proanthocyanidin Polymers. Phytochemistry 1980, 19, 1747–1754. [Google Scholar] [CrossRef]
- Ferreira, D.; Brandt, E.V.; Coetzee, J.; Malan, E. Condensed Tannins. In Fortschritte der Chemie Organischer Naturstoffe/Progress in the Chemistry of Organic Natural Products; Ayer, W.A., Brandt, E.V., Coetzee, J., Daniewski, W.M., Ferreira, D., Malan, E., Trifonov, L.S., Vidari, G., Eds.; Springer: Vienna, Austria, 1999; pp. 21–67. ISBN 978-3-7091-6366-5. [Google Scholar]
- Porter, L.J. Flavans and proanthocyanidins. In The Flavonoids: Advances in Research since 1980; Harborne, J.B., Ed.; Springer: Boston, MA, USA, 1988; pp. 21–62. ISBN 978-1-4899-2913-6. [Google Scholar]
- Ferreira, D.; Li, X.-C. Oligomeric Proanthocyanidins: Naturally Occurring O-Heterocycles. Nat. Prod. Rep. 2000, 17, 193–212. [Google Scholar] [CrossRef]
- Alioto, T.; Alexiou, K.G.; Bardil, A.; Barteri, F.; Castanera, R.; Cruz, F.; Dhingra, A.; Duval, H.; Fernández I Martí, Á.; Frias, L.; et al. Transposons Played a Major Role in the Diversification between the Closely Related Almond and Peach Genomes: Results from the Almond Genome Sequence. Plant J. Cell Mol. Biol. 2020, 101, 455–472. [Google Scholar] [CrossRef] [Green Version]
- Daccord, N.; Celton, J.-M.; Linsmith, G.; Becker, C.; Choisne, N.; Schijlen, E.; van de Geest, H.; Bianco, L.; Micheletti, D.; Velasco, R.; et al. High-Quality de Novo Assembly of the Apple Genome and Methylome Dynamics of Early Fruit Development. Nat. Genet. 2017, 49, 1099–1106. [Google Scholar] [CrossRef]
- Colle, M.; Leisner, C.P.; Wai, C.M.; Ou, S.; Bird, K.A.; Wang, J.; Wisecaver, J.H.; Yocca, A.E.; Alger, E.I.; Tang, H.; et al. Haplotype-Phased Genome and Evolution of Phytonutrient Pathways of Tetraploid Blueberry. GigaScience 2019, 8, giz012. [Google Scholar] [CrossRef] [Green Version]
- Kan, L.; Nie, S.; Hu, J.; Liu, Z.; Xie, M. Antioxidant Activities and Anthocyanins Composition of Seed Coats from Twenty-Six Kidney Bean Cultivars. J. Funct. Foods 2016, 26, 622–631. [Google Scholar] [CrossRef]
- Schmutz, J.; McClean, P.E.; Mamidi, S.; Wu, G.A.; Cannon, S.B.; Grimwood, J.; Jenkins, J.; Shu, S.; Song, Q.; Chavarro, C.; et al. A Reference Genome for Common Bean and Genome-Wide Analysis of Dual Domestications. Nat. Genet. 2014, 46, 707–713. [Google Scholar] [CrossRef] [Green Version]
- Jaillon, O.; Aury, J.-M.; Noel, B.; Policriti, A.; Clepet, C.; Casagrande, A.; Choisne, N.; Aubourg, S.; Vitulo, N.; Jubin, C.; et al. The Grapevine Genome Sequence Suggests Ancestral Hexaploidization in Major Angiosperm Phyla. Nature 2007, 449, 463–467. [Google Scholar] [CrossRef]
- Zhuang, W.; Chen, H.; Yang, M.; Wang, J.; Pandey, M.K.; Zhang, C.; Chang, W.-C.; Zhang, L.; Zhang, X.; Tang, R.; et al. The Genome of Cultivated Peanut Provides Insight into Legume Karyotypes, Polyploid Evolution and Crop Domestication. Nat. Genet. 2019, 51, 865–876. [Google Scholar] [CrossRef]
- Lee, C.C.; Dudonné, S.; Dubé, P.; Desjardins, Y.; Kim, J.H.; Kim, J.S.; Kim, J.-E.; Park, J.H.Y.; Lee, K.W.; Lee, C.Y. Comprehensive Phenolic Composition Analysis and Evaluation of Yak-Kong Soybean (Glycine Max) for the Prevention of Atherosclerosis. Food Chem. 2017, 234, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.; Thelen, J.J.; Cheng, J.; et al. Genome Sequence of the Palaeopolyploid Soybean. Nature 2010, 463, 178–183. [Google Scholar] [CrossRef] [Green Version]
- Edger, P.P.; Poorten, T.J.; VanBuren, R.; Hardigan, M.A.; Colle, M.; McKain, M.R.; Smith, R.D.; Teresi, S.J.; Nelson, A.D.L.; Wai, C.M.; et al. Origin and Evolution of the Octoploid Strawberry Genome. Nat. Genet. 2019, 51, 541–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, E.; Li, F.; Tong, W.; Yang, H.; Wang, S.; Zhao, J.; Liu, C.; Gao, L.; Tai, Y.; She, G.; et al. The Tea Plant Reference Genome and Improved Gene Annotation Using Long-Read and Paired-End Sequencing Data. Sci. Data 2019, 6, 122. [Google Scholar] [CrossRef] [Green Version]
- Bell-Lelong, D.A.; Cusumano, J.C.; Meyer, K.; Chapple, C. Cinnamate-4-Hydroxylase Expression in Arabidopsis (Regulation in Response to Development and the Environment). Plant Physiol. 1997, 113, 729–738. [Google Scholar] [CrossRef] [Green Version]
- Cochrane, F.C.; Davin, L.B.; Lewis, N.G. The Arabidopsis Phenylalanine Ammonia Lyase Gene Family: Kinetic Characterization of the Four PAL Isoforms. Phytochemistry 2004, 65, 1557–1564. [Google Scholar] [CrossRef]
- Feinbaum, R.L.; Ausubel, F.M. Transcriptional Regulation of the Arabidopsis Thaliana Chalcone Synthase Gene. Mol. Cell. Biol. 1988, 8, 1985–1992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirley, B.W.; Hanley, S.; Goodman, H.M. Effects of Ionizing Radiation on a Plant Genome: Analysis of Two Arabidopsis Transparent Testa Mutations. Plant Cell 1992, 4, 333–347. [Google Scholar] [CrossRef] [PubMed]
- Seitz, C.; Ameres, S.; Forkmann, G. Identification of the Molecular Basis for the Functional Difference between Flavonoid 3′-Hydroxylase and Flavonoid 3′,5′-Hydroxylase. FEBS Lett. 2007, 581, 3429–3434. [Google Scholar] [CrossRef] [PubMed]
- Charrier, B.; Coronado, C.; Kondorosi, A.; Ratet, P. Molecular Characterization and Expression of Alfalfa (Medicago Sativa L.) Flavanone-3-Hydroxylase and Dihydroflavonol-4-Reductase Encoding Genes. Plant Mol. Biol. 1995, 29, 773–786. [Google Scholar] [CrossRef]
- Fischer, T.C.; Halbwirth, H.; Meisel, B.; Stich, K.; Forkmann, G. Molecular Cloning, Substrate Specificity of the Functionally Expressed Dihydroflavonol 4-Reductases from Malus Domestica and Pyrus Communis Cultivars and the Consequences for Flavonoid Metabolism. Arch. Biochem. Biophys. 2003, 412, 223–230. [Google Scholar] [CrossRef]
- Saito, K.; Kobayashi, M.; Gong, Z.; Tanaka, Y.; Yamazaki, M. Direct Evidence for Anthocyanidin Synthase as a 2-Oxoglutarate-Dependent Oxygenase: Molecular Cloning and Functional Expression of CDNA from a Red Forma of Perilla Frutescens. Plant J. Cell Mol. Biol. 1999, 17, 181–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanner, G.J.; Francki, K.T.; Abrahams, S.; Watson, J.M.; Larkin, P.J.; Ashton, A.R. Proanthocyanidin Biosynthesis in Plants. Purification of Legume Leucoanthocyanidin Reductase and Molecular Cloning of Its CDNA. J. Biol. Chem. 2003, 278, 31647–31656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, D.-Y.; Sharma, S.B.; Paiva, N.L.; Ferreira, D.; Dixon, R.A. Role of Anthocyanidin Reductase, Encoded by BANYULS in Plant Flavonoid Biosynthesis. Science 2003, 299, 396–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolling, B.W. Almond Polyphenols: Methods of Analysis, Contribution to Food Quality, and Health Promotion. Compr. Rev. Food Sci. Food Saf. 2017, 16, 346–368. [Google Scholar] [CrossRef] [Green Version]
- Jeong, S.T.; Goto-Yamamoto, N.; Hashizume, K.; Esaka, M. Expression of the Flavonoid 3′-Hydroxylase and Flavonoid 3′,5′-Hydroxylase Genes and Flavonoid Composition in Grape (Vitis Vinifera). Plant Sci. 2006, 170, 61–69. [Google Scholar] [CrossRef]
- Liu, M.; Tian, H.; Wu, J.-H.; Cang, R.-R.; Wang, R.-X.; Qi, X.-H.; Xu, Q.; Chen, X.-H. Relationship between Gene Expression and the Accumulation of Catechin during Spring and Autumn in Tea Plants (Camellia Sinensis L.). Hortic. Res. 2015, 2, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.-Q.; Gan, R.-Y.; Ge, Y.-Y.; Zhang, D.; Corke, H. Polyphenols in Common Beans (Phaseolus Vulgaris L.): Chemistry, Analysis, and Factors Affecting Composition. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1518–1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshioka, Y.; Li, X.; Zhang, T.; Mitani, T.; Yasuda, M.; Nanba, F.; Toda, T.; Yamashita, Y.; Ashida, H. Black Soybean Seed Coat Polyphenols Prevent AAPH-Induced Oxidative DNA-Damage in HepG2 Cells. J. Clin. Biochem. Nutr. 2017, 60, 108–114. [Google Scholar] [CrossRef] [Green Version]
- Abrahams, S.; Lee, E.; Walker, A.R.; Tanner, G.J.; Larkin, P.J.; Ashton, A.R. The Arabidopsis TDS4 Gene Encodes Leucoanthocyanidin Dioxygenase (LDOX) and Is Essential for Proanthocyanidin Synthesis and Vacuole Development. Plant J. 2003, 35, 624–636. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, S.; Matsuda, F.; Tohge, T.; Yonekura-Sakakibara, K.; Yamazaki, M.; Saito, K.; Narumi, I. Metabolic Profiling and Cytological Analysis of Proanthocyanidins in Immature Seeds of Arabidopsis Thaliana Flavonoid Accumulation Mutants. Plant J. 2010, 62, 549–559. [Google Scholar] [CrossRef]
- Routaboul, J.-M.; Kerhoas, L.; Debeaujon, I.; Pourcel, L.; Caboche, M.; Einhorn, J.; Lepiniec, L. Flavonoid Diversity and Biosynthesis in Seed of Arabidopsis Thaliana. Planta 2006, 224, 96–107. [Google Scholar] [CrossRef] [PubMed]
- Aaby, K.; Ekeberg, D.; Skrede, G. Characterization of Phenolic Compounds in Strawberry (Fragaria × Ananassa) Fruits by Different HPLC Detectors and Contribution of Individual Compounds to Total Antioxidant Capacity. J. Agric. Food Chem. 2007, 55, 4395–4406. [Google Scholar] [CrossRef]
- Riihinen, K.; Jaakola, L.; Kärenlampi, S.; Hohtola, A. Organ-Specific Distribution of Phenolic Compounds in Bilberry (Vaccinium Myrtillus) and “northblue” Blueberry (Vaccinium Corymbosum × V. Angustifolium). Food Chem. 2008, 110, 156–160. [Google Scholar] [CrossRef] [PubMed]
- Debeaujon, I.; Léon-Kloosterziel, K.M.; Koornneef, M. Influence of the Testa on Seed Dormancy, Germination, and Longevity in Arabidopsis. Plant Physiol. 2000, 122, 403–414. [Google Scholar] [CrossRef] [Green Version]
- Qi, T.; Song, S.; Ren, Q.; Wu, D.; Huang, H.; Chen, Y.; Fan, M.; Peng, W.; Ren, C.; Xie, D. The Jasmonate-ZIM-Domain Proteins Interact with the WD-Repeat/BHLH/MYB Complexes to Regulate Jasmonate-Mediated Anthocyanin Accumulation and Trichome Initiation in Arabidopsis Thaliana. Plant Cell 2011, 23, 1795–1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catalá, R.; Medina, J.; Salinas, J. Integration of Low Temperature and Light Signaling during Cold Acclimation Response in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 16475–16480. [Google Scholar] [CrossRef] [Green Version]
- Cominelli, E.; Gusmaroli, G.; Allegra, D.; Galbiati, M.; Wade, H.K.; Jenkins, G.I.; Tonelli, C. Expression Analysis of Anthocyanin Regulatory Genes in Response to Different Light Qualities in Arabidopsis Thaliana. J. Plant Physiol. 2008, 165, 886–894. [Google Scholar] [CrossRef]
- Li, P.; Li, Y.-J.; Zhang, F.-J.; Zhang, G.-Z.; Jiang, X.-Y.; Yu, H.-M.; Hou, B.-K. The Arabidopsis UDP-Glycosyltransferases UGT79B2 and UGT79B3, Contribute to Cold, Salt and Drought Stress Tolerance via Modulating Anthocyanin Accumulation. Plant J. 2017, 89, 85–103. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Grain, D.; Bobet, S.; Gourrierec, J.L.; Thévenin, J.; Kelemen, Z.; Lepiniec, L.; Dubos, C. Complexity and Robustness of the Flavonoid Transcriptional Regulatory Network Revealed by Comprehensive Analyses of MYB–BHLH–WDR Complexes and Their Targets in Arabidopsis Seed. New Phytol. 2014, 202, 132–144. [Google Scholar] [CrossRef]
- Chen, M.; Penfield, S. Feedback Regulation of COOLAIR Expression Controls Seed Dormancy and Flowering Time. Science 2018, 360, 1014–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, A.; Zhao, M.; Leavitt, J.M.; Lloyd, A.M. Regulation of the Anthocyanin Biosynthetic Pathway by the TTG1/BHLH/Myb Transcriptional Complex in Arabidopsis Seedlings. Plant J. 2008, 53, 814–827. [Google Scholar] [CrossRef] [PubMed]
- Routaboul, J.-M.; Dubos, C.; Beck, G.; Marquis, C.; Bidzinski, P.; Loudet, O.; Lepiniec, L. Metabolite Profiling and Quantitative Genetics of Natural Variation for Flavonoids in Arabidopsis. J. Exp. Bot. 2012, 63, 3749–3764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stracke, R.; Ishihara, H.; Huep, G.; Barsch, A.; Mehrtens, F.; Niehaus, K.; Weisshaar, B. Differential Regulation of Closely Related R2R3-MYB Transcription Factors Controls Flavonol Accumulation in Different Parts of the Arabidopsis Thaliana Seedling. Plant J. 2007, 50, 660–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, L.; Pang, Y.; Dixon, R.A. Biosynthesis and Genetic Engineering of Proanthocyanidins and (Iso)Flavonoids. Phytochem. Rev. 2008, 7, 445–465. [Google Scholar] [CrossRef]
- Bogs, J.; Downey, M.O.; Harvey, J.S.; Ashton, A.R.; Tanner, G.J.; Robinson, S.P. Proanthocyanidin Synthesis and Expression of Genes Encoding Leucoanthocyanidin Reductase and Anthocyanidin Reductase in Developing Grape Berries and Grapevine Leaves. Plant Physiol. 2005, 139, 652–663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogs, J.; Jaffé, F.W.; Takos, A.M.; Walker, A.R.; Robinson, S.P. The Grapevine Transcription Factor VvMYBPA1 Regulates Proanthocyanidin Synthesis during Fruit Development. Plant Physiol. 2007, 143, 1347–1361. [Google Scholar] [CrossRef] [Green Version]
- Todd, J.J.; Vodkin, L.O. Pigmented Soybean (Glycine Max) Seed Coats Accumulate Proanthocyanidins during Development. Plant Physiol. 1993, 102, 663–670. [Google Scholar] [CrossRef] [Green Version]
- Tuteja, J.H.; Zabala, G.; Varala, K.; Hudson, M.; Vodkin, L.O. Endogenous, Tissue-Specific Short Interfering RNAs Silence the Chalcone Synthase Gene Family in Glycine Max Seed Coats. Plant Cell 2009, 21, 3063–3077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zabala, G.; Vodkin, L. Cloning of the Pleiotropic T Locus in Soybean and Two Recessive Alleles That Differentially Affect Structure and Expression of the Encoded Flavonoid 3′ Hydroxylase. Genetics 2003, 163, 295–309. [Google Scholar] [CrossRef] [PubMed]
- Zabala, G.; Vodkin, L.O. The Wp Mutation of Glycine Max Carries a Gene-Fragment-Rich Transposon of the CACTA Superfamily. Plant Cell 2005, 17, 2619–2632. [Google Scholar] [CrossRef] [Green Version]
- Zabala, G.; Vodkin, L.O. A Rearrangement Resulting in Small Tandem Repeats in the F3′ 5′ H Gene of White Flower Genotypes Is Associated with the Soybean W1 Locus. Crop Sci. 2007, 47, S-113. [Google Scholar] [CrossRef]
- Gillman, J.D.; Tetlow, A.; Lee, J.-D.; Shannon, J.G.; Bilyeu, K. Loss-of-Function Mutations Affecting a Specific Glycine Max R2R3 MYB Transcription Factor Result in Brown Hilum and Brown Seed Coats. BMC Plant Biol. 2011, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Yang, K.; Jeong, N.; Moon, J.-K.; Lee, Y.-H.; Lee, S.-H.; Kim, H.M.; Hwang, C.H.; Back, K.; Palmer, R.G.; Jeong, S.-C. Genetic Analysis of Genes Controlling Natural Variation of Seed Coat and Flower Colors in Soybean. J. Hered. 2010, 101, 757–768. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Sekizaki, H.; Yang, Z.; Sawa, S.; Pan, J. Phenolics in the Seed Coat of Wild Soybean (Glycine Soja) and Their Significance for Seed Hardness and Seed Germination. J. Agric. Food Chem. 2010, 58, 10972–10978. [Google Scholar] [CrossRef] [PubMed]
- Ito, C.; Oki, T.; Yoshida, T.; Nanba, F.; Yamada, K.; Toda, T. Characterisation of Proanthocyanidins from Black Soybeans: Isolation and Characterisation of Proanthocyanidin Oligomers from Black Soybean Seed Coats. Food Chem. 2013, 141, 2507–2512. [Google Scholar] [CrossRef]
- Kovinich, N.; Saleem, A.; Arnason, J.T.; Miki, B. Combined Analysis of Transcriptome and Metabolite Data Reveals Extensive Differences between Black and Brown Nearly-Isogenic Soybean (Glycine Max) Seed Coats Enabling the Identification of Pigment Isogenes. BMC Genomics 2011, 12, 381. [Google Scholar] [CrossRef] [Green Version]
- Bursać, M.; Krstonošić, M.A.; Miladinović, J.; Malenčić, Đ.; Gvozdenović, L.; Cvejić, J.H. Isoflavone Composition, Total Phenolic Content and Antioxidant Capacity of Soybeans with Colored Seed Coat. Nat. Prod. Commun. 2017, 12, 1934578X1701200417. [Google Scholar] [CrossRef] [Green Version]
- Maloney, G.S.; DiNapoli, K.T.; Muday, G.K. The Anthocyanin Reduced Tomato Mutant Demonstrates the Role of Flavonols in Tomato Lateral Root and Root Hair Development. Plant Physiol. 2014, 166, 614–631. [Google Scholar] [CrossRef] [Green Version]
- Buer, C.S.; Imin, N.; Djordjevic, M.A. Flavonoids: New Roles for Old Molecules. J. Integr. Plant Biol. 2010, 52, 98–111. [Google Scholar] [CrossRef] [PubMed]
- Baudry, A.; Heim, M.A.; Dubreucq, B.; Caboche, M.; Weisshaar, B.; Lepiniec, L. TT2, TT8, and TTG1 Synergistically Specify the Expression of BANYULS and Proanthocyanidin Biosynthesis in Arabidopsis Thaliana. Plant J. 2004, 39, 366–380. [Google Scholar] [CrossRef] [PubMed]
- Feuillet, C.; Schachermayr, G.; Keller, B. Molecular Cloning of a New Receptor-like Kinase Gene Encoded at the Lr10 Disease Resistance Locus of Wheat. Plant J. 1997, 11, 45–52. [Google Scholar] [CrossRef]
- Tohge, T.; Nishiyama, Y.; Hirai, M.Y.; Yano, M.; Nakajima, J.; Awazuhara, M.; Inoue, E.; Takahashi, H.; Goodenowe, D.B.; Kitayama, M. Functional Genomics by Integrated Analysis of Metabolome and Transcriptome of Arabidopsis Plants Over-expressing an MYB Transcription Factor. Plant J. 2005, 42, 218–235. [Google Scholar] [CrossRef] [PubMed]
- Xie, D.-Y.; Sharma, S.B.; Wright, E.; Wang, Z.-Y.; Dixon, R.A. Metabolic Engineering of Proanthocyanidins through Co-Expression of Anthocyanidin Reductase and the PAP1 MYB Transcription Factor. Plant J. 2006, 45, 895–907. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Yu, H.; Tang, H.; Wang, X. Identification and Expression Analysis of Genes Involved in Anthocyanin and Proanthocyanidin Biosynthesis in the Fruit of Blackberry. Sci. Hortic. 2012, 141, 61–68. [Google Scholar] [CrossRef]
- De Pascual-Teresa, S.; Moreno, D.A.; García-Viguera, C. Flavanols and Anthocyanins in Cardiovascular Health: A Review of Current Evidence. Int. J. Mol. Sci. 2010, 11, 1679–1703. [Google Scholar] [CrossRef] [Green Version]
- Hammerstone, J.F.; Lazarus, S.A.; Schmitz, H.H. Procyanidin Content and Variation in Some Commonly Consumed Foods. J. Nutr. 2000, 130, 2086S–2092S. [Google Scholar] [CrossRef]
- Ferreira, D.; Nel, R.J.J.; Bekker, R. 3.19—Condensed Tannins. In Comprehensive Natural Products Chemistry; Barton, S.D., Nakanishi, K., Meth-Cohn, O., Eds.; Pergamon: Oxford, UK, 1999; ISBN 978-0-08-091283-7. [Google Scholar]
- Ferreira, D.; Marais, J.P.J.; Coleman, C.M.; Slade, D. 6.18—Proanthocyanidins: Chemistry and Biology. In Comprehensive Natural Products II; Liu, H.-W., Mander, L., Eds.; Elsevier: Oxford, UK, 2010; pp. 605–661. ISBN 978-0-08-045382-8. [Google Scholar]
- Dixon, R.A.; Sarnala, S. Proanthocyanidin Biosynthesis—A Matter of Protection. Plant Physiol. 2020, 184, 579–591. [Google Scholar] [CrossRef]
- Kobayashi, H.; Suzuki, S.; Tanzawa, F.; Takayanagi, T. Low Expression of Flavonoid 3′,5′-Hydroxylase (F3′,5′H) Associated with Cyanidin-Based Anthocyanins in Grape Leaf. Am. J. Enol. Vitic. 2009, 60, 362–367. [Google Scholar]
- Dixon, R.A.; Liu, C.; Jun, J.H. Metabolic Engineering of Anthocyanins and Condensed Tannins in Plants. Curr. Opin. Biotechnol. 2013, 24, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castresana, J. Selection of Conserved Blocks from Multiple Alignments for Their Use in Phylogenetic Analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Species | Proanthocyanidins Content (mg/100 g) | Reference | Genome Database | Reference | Assembly Size (Mb) | Coverage (%) | Contig N50 (Kb) | Number of Genes Predicted |
|---|---|---|---|---|---|---|---|---|
| Almond (Prunus dulcis) | 184 | Prior & Gu, 2005 [14] | Prunus dulcis Lauranne Genome v1.0 (http://rosaceae.org/, accessed on 12 February 2021) | Alioto et al., 2020 [31] | 227 | 95 | 103 | 27,969 |
| Apple tree (Malus domestica) | 162 | Hellström et al., 2009 [2] | (iris.angers.inra.fr/gddh13/, accessed on 12 February 2021) | Daccord et al., 2017 [32] | 643 | 100 | 620 | 42,140 |
| Blueberry (Vaccinium corymbosum) | 255 | Prior & Gu, 2005 [14] | V_corymbosum v1.0 (http://gigadb.org/, accessed on 12 February 2021) | Colle et al., 2019 [33] | 1680 | 102 | 15 | 32,140 |
| Cacao bean (Theobroma cacao) | 1460 | Hellström et al., 2009 [2] | Cacao Matina1-6 Genome v2.1 (http://cacaogenomedb.org, accessed on 12 February 2021) | Publication in progress (http://cacaogenomedb.org) | 346 | 80 | 1080 | 27,379 |
| Common bean (Phaseolus vulgaris) | 1000 | Kan et al., 2016 [34] | Phaseolus vulgaris v2.1 (http://phytozome.jgi.doe.gov/, accessed on 12 February 2021) | Schmutz et al., 2014 [35] | 600 | 80 | 1900 | 27,433 |
| Grape (Vitis vinifera) | 3532 | Prior & Gu, 2005 [14] | Vitis vinifera v2.1 (http://phytozome.jgi.doe.gov/, accessed on 12 February 2021) | Jaillon et al., 2007 [36] | 487 | 102 | 566 | 26,346 |
| Peanut (Arachis hypogaea) | 186 | Hellström et al., 2009 [2] | (http://peanutgr.fafu.edu.cn/, accessed on 12 February 2021) | Zhuang et al., 2019 [37] | 2538 | 94 | 1509 | 83,709 |
| Soybean (Glycine max) | 300 | Lee et al., 2017 [38] | Glycine max Wm82.a4 (http://www.soybase.org, accessed on 12 February 2021) | Schmutz et al., 2010 [39] | 1150 | 95 | 1492 | 46,430 |
| Strawberry (Fragaria × ananassa) | 145 | Prior & Gu, 2005 [14] | (https://datadryad.org/, accessed on 12 February 2021) | Edger et al., 2019 [40] | 813 * | 99 | 79 | 108,087 |
| Tea tree (Camellia sinensis) | 189 | Engelhardt et al., 2003 [12] | (http://tpia.teaplant.org/, accessed on 12 February 2021) | Xia et al., 2019 [41] | 2890 | 95 | 67 | 53,512 |
| Species | a Number of Genes from KEGG Pathway | b Number of Genes Confirmed from Reference Database | c Number of Genes Newly Identified Using a | Number of Orthologous Genes Newly Identified Using b+c |
|---|---|---|---|---|
| Arachis hypogea | 6 * | 6 | 5 | 1 |
| Arabidopsis thaliana | 6 | 6 | - | - |
| Fragaria × ananassa | 4 * | 4 | 9 | 7 |
| Glycine max | 20 | 19 | - | - |
| Camelia sinensis | - | - | - | 19 |
| Malus domestica | 12 | 12 | - | - |
| Phaseolus vulgaris | 12 | 11 | - | - |
| Theobroma cacao | 9 | 8 | - | - |
| Vitis vinifera | 23 | 17 | - | - |
| Prunus dulcis | - | - | - | 4 |
| Vaccinium corymbosum | - | - | - | 74 |
| Total | 92 | 83 | 14 | 105 |
| Arachis hypogea | Arabidopsis thaliana | Fragaria × ananassa | Glycine max | Malus domestica | Phaseolus vulgaris | Theobroma cacao | Vitis vinifera | Camelia sinensis | Prunus dulcis | Vaccinium corymbosum | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| F3′H | 2 | 1 | 3 | 5 | 2 | 1 | 1 | 2 | 1 | 0 | 4 |
| F3H | 2 | 1 | 4 | 4 | 2 | 1 | 1 | 2 | 2 | 0 | 8 |
| F3′5′H | 0 | 0 | 0 | 1 | 0 | 2 | 1 | 7 | 5 | 0 | 37 |
| DFR | 2 | 1 | 4 | 3 | 2 | 3 | 1 | 3 | 3 | 1 | 6 |
| LAR | 2 | 0 | 4 | 2 | 2 | 1 | 2 | 2 | 3 | 1 | 12 |
| ANS | 2 | 2 | 3 | 2 | 2 | 1 | 1 | 1 | 3 | 1 | 3 |
| ANR | 2 | 1 | 2 | 2 | 2 | 2 | 1 | 0 | 2 | 1 | 4 |
| Total | 12 | 6 | 20 | 19 | 12 | 11 | 8 | 17 | 19 | 4 | 74 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, I.; Ha, J. Biosynthetic Pathway of Proanthocyanidins in Major Cash Crops. Plants 2021, 10, 1792. https://doi.org/10.3390/plants10091792
Lim I, Ha J. Biosynthetic Pathway of Proanthocyanidins in Major Cash Crops. Plants. 2021; 10(9):1792. https://doi.org/10.3390/plants10091792
Chicago/Turabian StyleLim, Insu, and Jungmin Ha. 2021. "Biosynthetic Pathway of Proanthocyanidins in Major Cash Crops" Plants 10, no. 9: 1792. https://doi.org/10.3390/plants10091792
APA StyleLim, I., & Ha, J. (2021). Biosynthetic Pathway of Proanthocyanidins in Major Cash Crops. Plants, 10(9), 1792. https://doi.org/10.3390/plants10091792