
Deciphering the Physicochemical and Microscopical Changes in Ganoderma boninense-Infected Oil Palm Woodblocks under the Influence of Phenolic Compounds



Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Microorganism, Culture Conditions and Treatments
2.3. Biodegradation of Woodblocks
2.4. Anatomical Characterization during Biodegradation of Oil Palm Woodblocks
Scanning Electron Microscopy (SEM) Analysis
2.5. Chemical Characterization during Biodegradation of Oil Palm Woodblocks
2.5.1. Fourier Transform Infrared (FT-IR) Spectroscopy Analysis
2.5.2. Thermogravimetry (TGA) Analysis
2.6. Statistical Analyses
3. Results
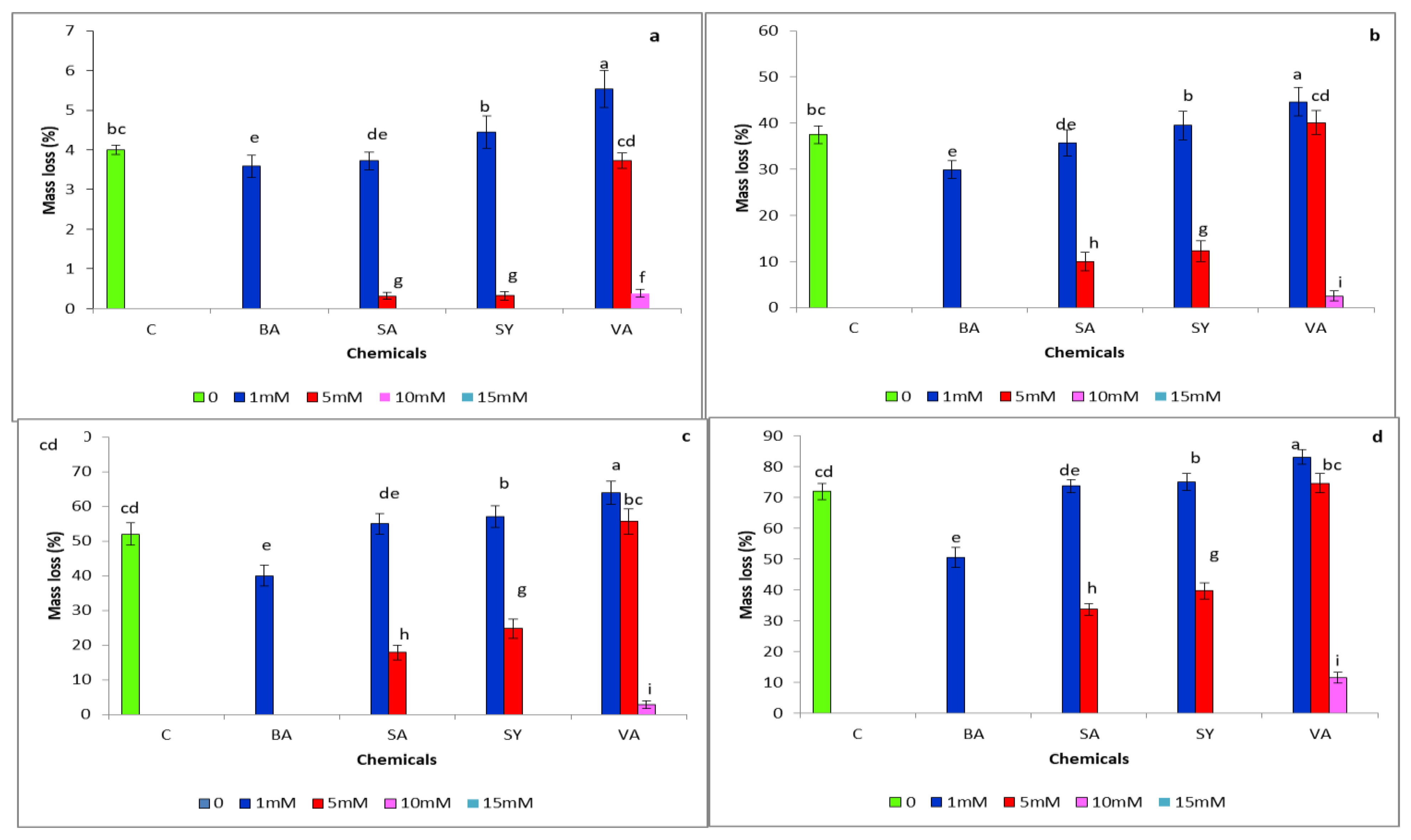
3.1. Mass Loss
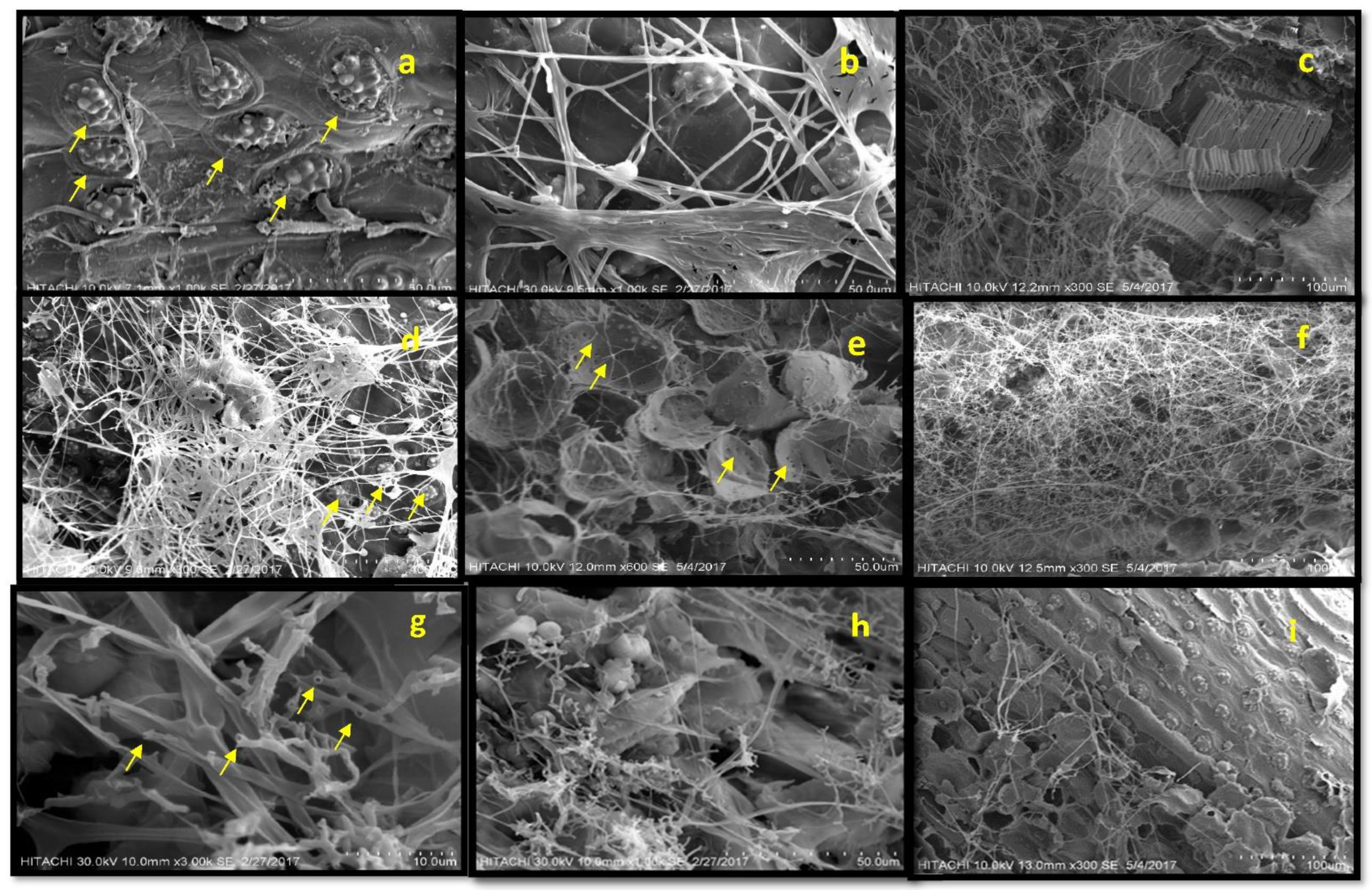
3.2. Anatomical Characterization during Biodegradation of Oil Palm Wood
Scanning Electron Microscopy (SEM) Analysis
3.3. Chemical Characterization during Biodegradation of Oil Palm Woodblocks
3.3.1. Fourier Transform Infrared (FT-IR) Spectroscopy Analysis
3.3.2. Thermogravimetry (TGA) Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mohammed, C.; Rimbawanto, A.; Page, D. Management of basidiomycete root-and stem-rot diseases in oil palm, rubber and tropical hardwood plantation crops. For. Pathol. 2014, 44, 428–446. [Google Scholar] [CrossRef]
- Olaniyi, O.N.; Szulczyk, K.R. Estimating the economic damage and treatment cost of basal stem rot striking the Malaysian oil palms. For. Policy Econ. 2020, 116, 102163. [Google Scholar] [CrossRef]
- Midot, F.; Lau, S.Y.L.; Wong, W.C.; Tung, H.J.; Yap, M.L.; Lo, M.L.; Jee, M.S.; Dom, S.P.; Melling, L. Genetic Diversity and Demographic History of Ganoderma boninense in Oil Palm Plantations of Sarawak, Malaysia Inferred from ITS Regions. Microorganisms 2019, 7, 464. [Google Scholar] [CrossRef] [Green Version]
- Roslan, A.; Idris, A. Economic impact of Ganoderma incidence on Malaysian oil palm plantation—A case study in Johor. Oil Palm Ind. Econ. J. 2012, 12, 24–30. [Google Scholar]
- Chung, G. Management of Ganoderma diseases in oil palm plantations. Planter 2011, 87, 325–339. [Google Scholar]
- Flood, J.; Hasan, Y.; Turner, P.; O’Grady, E. The spread of Ganoderma from infective sources in the field and its implications for management of the disease in oil palm. In Ganoderma Diseases of Perennial Crops; CABI: Wallingford, UK, 2000; pp. 101–112. [Google Scholar]
- Pilotti, C.; Gorea, E.; Bonneau, L. Basidiospores as sources of inoculum in the spread of Ganoderma boninense in oil palm plantations in Papua New Guinea. Plant Pathol. 2018, 67, 1841–1849. [Google Scholar] [CrossRef]
- Susanto, A.; Sudharto, P.; Purba, R. Enhancing biological control of basal stem rot disease (Ganoderma boninense) in oil palm plantations. Mycopathologia 2005, 159, 153–157. [Google Scholar] [CrossRef]
- Khairudin, H. Basal Stem Rot of Oil Palm Caused by Ganoderma boninense: An Update, Proceedings of the International Palm Oil Congress ‘Update and Vision’ (Agriculture), Bangi, Selangor, Malaysia, 20–25 September 1993; Paper No. 46; Palm Oil Research Institute of Malaysia: Bangi, Selangor, Malaysia, 1993.
- Jourdan, C.; Rey, H. Architecture and development of the oil-palm (Elaeis guineensis Jacq.) root system. Plant Soil 1997, 189, 33–48. [Google Scholar] [CrossRef]
- Surendran, A.; Siddiqui, Y.; Saud, H.M.; Manickam, N. The antagonistic effect of phenolic compounds on ligninolytic and cellulolytic enzymes of Ganoderma Boninense, causing basal stem rot in Oil Palm. Int. J. Agric. Biol. 2017, 19, 1437–1446. [Google Scholar]
- Surendran, A.; Siddiqui, Y.; Saud, H.M.; Ali, N.S.; Manickam, S. Inhibition and kinetic studies of lignin degrading enzymes of Ganoderma boninense by naturally occurring phenolic compounds. J. Appl. Microbiol. 2018, 125, 876–887. [Google Scholar] [CrossRef]
- Schirp, A.; Wolcott, M.P. Influence of fungal decay and moisture absorption on mechanical properties of extruded wood-plastic composites. Wood Fiber Sci. 2005, 37, 643–652. [Google Scholar]
- Bucher, V.; Hyde, K.; Pointing, S.; Reddy, C. Production of wood decay enzymes, mass loss and lignin solubilization in wood by marine ascomycetes and their anamorphs. Fungal Divers. 2004, 15, 1–4. [Google Scholar]
- Popescu, C.-M.; Popescu, M.-C.; Vasile, C. Structural changes in biodegraded lime wood. Carbohydr. Polym. 2010, 79, 362–372. [Google Scholar] [CrossRef]
- Nelson, M.L.; O’Connor, R.T. Relation of certain infrared bands to cellulose crystallinity and crystal lattice type. Part II. A new infrared ratio for estimation of crystallinity in celluloses I and II. J. Appl. Polym. Sci. 1964, 8, 1325–1341. [Google Scholar] [CrossRef]
- Fan, M.; Dai, D.; Huang, B. Fourier transform infrared spectroscopy for natural fibres. In Fourier Transform-Materials Analysis; Intechopen: London, UK, 2012. [Google Scholar]
- Yang, H.; Yan, R.; Chin, T.; Liang, D.T.; Chen, H.; Zheng, C. Thermogravimetric analysis− Fourier transform infrared analysis of palm oil waste pyrolysis. Energy Fuels 2004, 18, 1814–1821. [Google Scholar] [CrossRef]
- Pandey, K.K.; Pitman, A. FTIR studies of the changes in wood chemistry following decay by brown-rot and white-rot fungi. Int. Biodeterior. Biodegrad. 2003, 52, 151–160. [Google Scholar] [CrossRef]
- Blanchette, R.A. A review of microbial deterioration found in archaeological wood from different environments. Int. Biodeterior. Biodegrad. 2000, 46, 189–204. [Google Scholar] [CrossRef]
- Kirk, T.K.; Moore, W.E. Removing lignin from wood with white-rot fungi and digestibility of resulting wood. Wood Fiber Sci. 2007, 4, 72–79. [Google Scholar]
- Alfredsen, G.; Bader, T.K.; Dibdiakova, J.; Filbakk, T.; Bollmus, S.; Hofstetter, K. Thermogravimetric analysis for wood decay characterisation. Eur. J. Wood Wood Prod. 2012, 70, 527–530. [Google Scholar] [CrossRef]
- Su, Y.; Yu, X.; Sun, Y.; Wang, G.; Chen, H.; Chen, G. Evaluation of screened lignin-degrading fungi for the biological pretreatment of corn stover. Sci. Rep. 2018, 8, 5385. [Google Scholar] [CrossRef] [Green Version]
- Luna, M.L.; Murace, M.A.; Keil, G.D.; Otaño, M.E. Patterns of decay caused by Pycnoporus sanguineus and Ganoderma lucidum (Aphyllophorales) in poplar wood. IAWA J. 2004, 25, 425–433. [Google Scholar] [CrossRef] [Green Version]
- Kües, U.; Mai, C.; Militz, H.; Kües, U. 14. Biological Wood Protection against Decay, Microbial Staining, Fungal Moulding and Insect Pests. In Wood Production, Wood Technology, and Biotechnological Impacts; 2007; pp. 273–294. Available online: https://www.researchgate.net/publication/262179411_Biological_Wood_Protection_against_Decay_Microbial_Staining_Fungal_Moulding_and_Insect_Pests.
- Del Rıo, J.; Speranza, M.; Gutiérrez, A.; Martınez, M.; Martınez, A. Lignin attack during eucalypt wood decay by selected basidiomycetes: A Py-GC/MS study. J. Anal. Appl. Pyrolysis 2002, 64, 421–431. [Google Scholar] [CrossRef] [Green Version]
- Chappell, J.; Hahlbrock, K. Transcription of plant defence genes in response to UV light or fungal elicitor. Nature 1984, 311, 76–78. [Google Scholar] [CrossRef]
- Bari, E.; Nazarnezhad, N.; Kazemi, S.M.; Ghanbary, M.A.T.; Mohebby, B.; Schmidt, O.; Clausen, C.A. Comparison between degradation capabilities of the white rot fungi Pleurotus ostreatus and Trametes versicolor in beech wood. Int. Biodeterior. Biodegrad. 2015, 104, 231–237. [Google Scholar] [CrossRef]
- Darwish, S.S.; El Hadidi, N.; Mansour, M. The effect of fungal decay on Ficus sycomoru wood. Int. J. Conserv. Sci. 2013, 4, 271–282. [Google Scholar]
- Susanto, A.; Suprianto, E.; Purba, A.; Prasetyo, A. Oil palm breeding for tolerance to G. boninense. In Disease Management Strategies in Plantations, Yagyakarta; Indonesian Oil Palm Research Institute: Medan, Indonesia, 2009; pp. 4–8. [Google Scholar]
- Naidu, Y.; Siddiqui, Y.; Rafii, M.Y.; Saud, H.M.; Idris, A.S. Inoculation of oil palm seedlings in Malaysia with white-rot hymenomycetes: Assessment of pathogenicity and vegetative growth. Crop Prot. 2018, 110, 146–154. [Google Scholar] [CrossRef]
- Cragg, S.M.; Beckham, G.T.; Bruce, N.C.; Bugg, T.D.; Distel, D.L.; Dupree, P.; Etxabe, A.G.; Goodell, B.S.; Jellison, J.; McGeehan, J.E. Lignocellulose degradation mechanisms across the Tree of Life. Curr. Opin. Chem. Biol. 2015, 29, 108–119. [Google Scholar] [CrossRef] [Green Version]
- Guillén, F.; Martínez, M.J.; Gutiérrez, A.; Del Rio, J. Biodegradation of lignocellu-losics: Microbial, chemical, and enzymatic aspects of the fungal attack of lignin. Int. Microbiol. 2005, 8, 195–204. [Google Scholar]
- Buswell, J.; Ander, P.; Pettersson, B.; Eriksson, K.-E. Oxidative decarboxylation of vanillic acid by Sporotrichum pulverulentum. FEBS Lett. 1979, 103, 98–101. [Google Scholar] [CrossRef] [Green Version]
- Gupta, J.K.; Hamp, S.G.; Buswell, J.A.; Eriksson, K.-E. Metabolism of trans-ferulic acid by the white-rot fungus Sporotrichum pulverulentum. Arch. Microbiol. 1981, 128, 349–354. [Google Scholar] [CrossRef]
- Mishra, V.; Jana, A.K.; Jana, M.M.; Gupta, A. Fungal pretreatment of sweet sorghum bagasse with supplements: Improvement in lignin degradation, selectivity and enzymatic saccharification. 3 Biotech 2017, 7, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surendran, A.; Siddiqui, Y.; Ali, N.S.; Manickam, S. Inhibition and kinetic studies of cellulose-and hemicellulose-degrading enzymes of Ganoderma boninense by naturally occurring phenolic compounds. J. Appl. Microbiol. 2018, 124, 1544–1555. [Google Scholar] [CrossRef] [PubMed]
- Carrier, M.; Loppinet-Serani, A.; Denux, D.; Lasnier, J.-M.; Ham-Pichavant, F.; Cansell, F.; Aymonier, C. Thermogravimetric analysis as a new method to determine the lignocellulosic composition of biomass. Biomass Bioenergy 2011, 35, 298–307. [Google Scholar] [CrossRef]
- Hastrup, A.C.S.; Howell, C.; Larsen, F.H.; Sathitsuksanoh, N.; Goodell, B.; Jellison, J. Differences in crystalline cellulose modification due to degradation by brown and white-rot fungi. Fungal Biol. 2012, 116, 1052–1063. [Google Scholar] [CrossRef] [PubMed]
- Hall, M.; Bansal, P.; Lee, J.H.; Realff, M.J.; Bommarius, A.S. Cellulose crystallinity–a key predictor of the enzymatic hydrolysis rate. FEBS J. 2010, 277, 1571–1582. [Google Scholar] [CrossRef]
- Dixon, R.A.; Paiva, N.L. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085. [Google Scholar] [CrossRef]
- Idris, A.; Maizatul, S. Stump treatment with dazomet for controlling Ganoderma disease in oil palm. MPOB TS Inf. Ser. 2012, 107, 615–616. [Google Scholar]
- Srivastava, P.; Andersen, P.C.; Marois, J.J.; Wright, D.L.; Srivastava, M.; Harmon, P.F. Effect of phenolic compounds on growth and ligninolytic enzyme production in Botryosphaeria isolates. Crop Prot. 2013, 43, 146–156. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Wave Number (cm−1) | Assignment | Source | Observation | References |
|---|---|---|---|---|
| Lignin Bands | ||||
| 1700 | C=O stretching of conjugated or aromatic ketones | Lignin | Appeared in wood samples treated with 1 mM benzoic, 5 mM salicylic and 15 mM vanillic acids. Later disappeared on the 120th day. | [19] |
| 1612 | Unconjugated carboxyl stretch of both lignin and cellulose | Lignin | A high-intensity band appeared in woodblocks treated with 1 mM syringic and vanillic acids. A medium-intensity band was observed in 1 mM benzoic acid initially, and was not observed in the later stages of degradation. | [20,21] |
| 1620 | C=C stretching in the aromatic groups of lignin | Lignin | This band was observed in all the treated as well as the control wood samples. | [21,22] |
| 1670 | C=O stretching in the conjugated p-substituted aryl ketone | Lignin | This band was observed in all the treated as well as the control wood samples. | [19,23] |
| 1270 | C-O stretching in xylene and hemicellulose and guaiacyl structure in lignin | Lignin | The intensity of the band decreased when the lignin and the adjacent hemicellulose degraded appeared in the control wood samples, along with wood samples treated with 1 mM phenolic compounds. It appeared in all the other wood samples as the degradation proceeded. | [24] |
| 1034 | Deformation vibration of C-H bond in aromatic rings | Lignin | This band was observed in all the treated as well as the control wood samples. | [25] |
| 1247 | C-O stretching in lignin (Guaiacyl units) and hemicellulose | Lignin | This band was observed in all the treated as well as the control wood samples. However, the highest intensity of this band was observed in wood samples treated with 1 mM vanillic acid. | [19,23] |
| Carbohydrate Bands | ||||
| 1336 | OH in-plane bending cellulose | Cellulose | A weak band appeared only at the end of the degradation period. | [26,27] |
| 1320 | C-H variation in cellulose and C-O stretching in syringyl unit of lignin | Cellulose and lignin | The intensity of the band decreased when the lignin and the adjacent hemicellulose degraded. | [28] |
| 128 | C-H stretching | Starch | A weak band appeared only at the end of the degradation period. | [14] |
| Hydroxy Bands | ||||
| 3427 | Intra-molecular OH stretching in cellulose | Cellulose | Increased intensity of this band indicates that more hydroxyl groups are available resulting from the hydrolysis. The increment in these two bands was observed throughout the degradation process. | [29] |
| 3340 | Bonded OH stretching | Carbohydrate and lignin | ||
| Biodegradation (Days) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Phenolic Compounds (mM) | 10th Day | 30th Day | 45th Day | 120th Day | ||||||||
| Relative (%) Degradation | ||||||||||||
| L | C | H | L | C | H | L | C | H | L | C | H | |
| Control | 49.6 | 37.1 | 68.9 | 55.6 | 46.5 | 69.9 | 64.7 | 46.5 | 70.6 | 72.2 | 60.9 | 71.1 |
| BA 1 | 2.3 | 14.4 | 18.7 | 54.9 | 44.5 | 35.5 | 56.4 | 47.5 | 49.2 | 63.2 | 63.9 | 74.2 |
| SA 1 SA 5 | 47.4 | 15.1 | 33.4 | 54.9 | 40.1 | 48.5 | 57.1 | 49.5 | 66.6 | 72.2 | 61.9 | 70.6 |
| 32.3 | 9.0 | 22.4 | 45.7 | 27.1 | 39.8 | 49.3 | 29.8 | 41.1 | 68.0 | 35.8 | 45.8 | |
| SY 1 SY 5 | 54.9 | 34.8 | 49.2 | 62.4 | 47.5 | 50.5 | 66.9 | 53.8 | 52.5 | 71.4 | 66.2 | 69.2 |
| 10.5 | 5.4 | 23.7 | 25.6 | 13.4 | 27.8 | 32.3 | 25.4 | 33.1 | 49.6 | 29.1 | 49.5 | |
| VA 1 VA 5 VA10 | 62.4 | 40.5 | 64.9 | 69.9 | 48.2 | 67.6 | 72.2 | 52.2 | 71.2 | 85.0 | 69.6 | 76.9 |
| 54.9 | 57.5 | 51.2 | 63.9 | 60.9 | 60.9 | 66.2 | 61.5 | 63.9 | 69.9 | 65.2 | 77.9 | |
| 1.5 | 2.5 | 22.7 | 24.8 | 17.1 | 31.4 | 33.1 | 19.7 | 32.8 | 62.4 | 20.4 | 42.5 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Surendran, A.; Siddiqui, Y.; Ahmad, K.; Fernanda, R. Deciphering the Physicochemical and Microscopical Changes in Ganoderma boninense-Infected Oil Palm Woodblocks under the Influence of Phenolic Compounds. Plants 2021, 10, 1797. https://doi.org/10.3390/plants10091797
Surendran A, Siddiqui Y, Ahmad K, Fernanda R. Deciphering the Physicochemical and Microscopical Changes in Ganoderma boninense-Infected Oil Palm Woodblocks under the Influence of Phenolic Compounds. Plants. 2021; 10(9):1797. https://doi.org/10.3390/plants10091797
Chicago/Turabian StyleSurendran, Arthy, Yasmeen Siddiqui, Khairulmazmi Ahmad, and Rozi Fernanda. 2021. "Deciphering the Physicochemical and Microscopical Changes in Ganoderma boninense-Infected Oil Palm Woodblocks under the Influence of Phenolic Compounds" Plants 10, no. 9: 1797. https://doi.org/10.3390/plants10091797
APA StyleSurendran, A., Siddiqui, Y., Ahmad, K., & Fernanda, R. (2021). Deciphering the Physicochemical and Microscopical Changes in Ganoderma boninense-Infected Oil Palm Woodblocks under the Influence of Phenolic Compounds. Plants, 10(9), 1797. https://doi.org/10.3390/plants10091797