
Genetic Diversity of Phenotypic and Biochemical Traits in VIR Radish (Raphanus sativus L.) Germplasm Collection



Abstract
:1. Introduction
2. Results
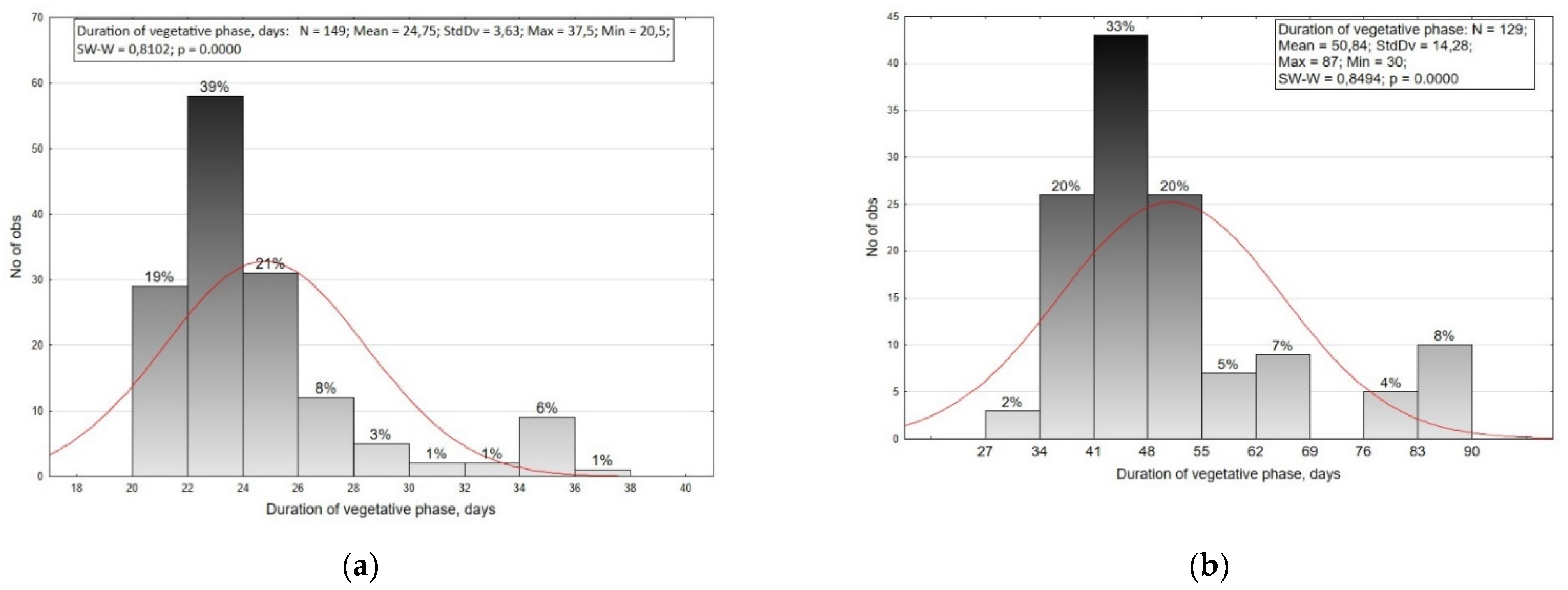
2.1. Variability of Phenotypic Traits
2.2. Variability of Biochemical Characteristics
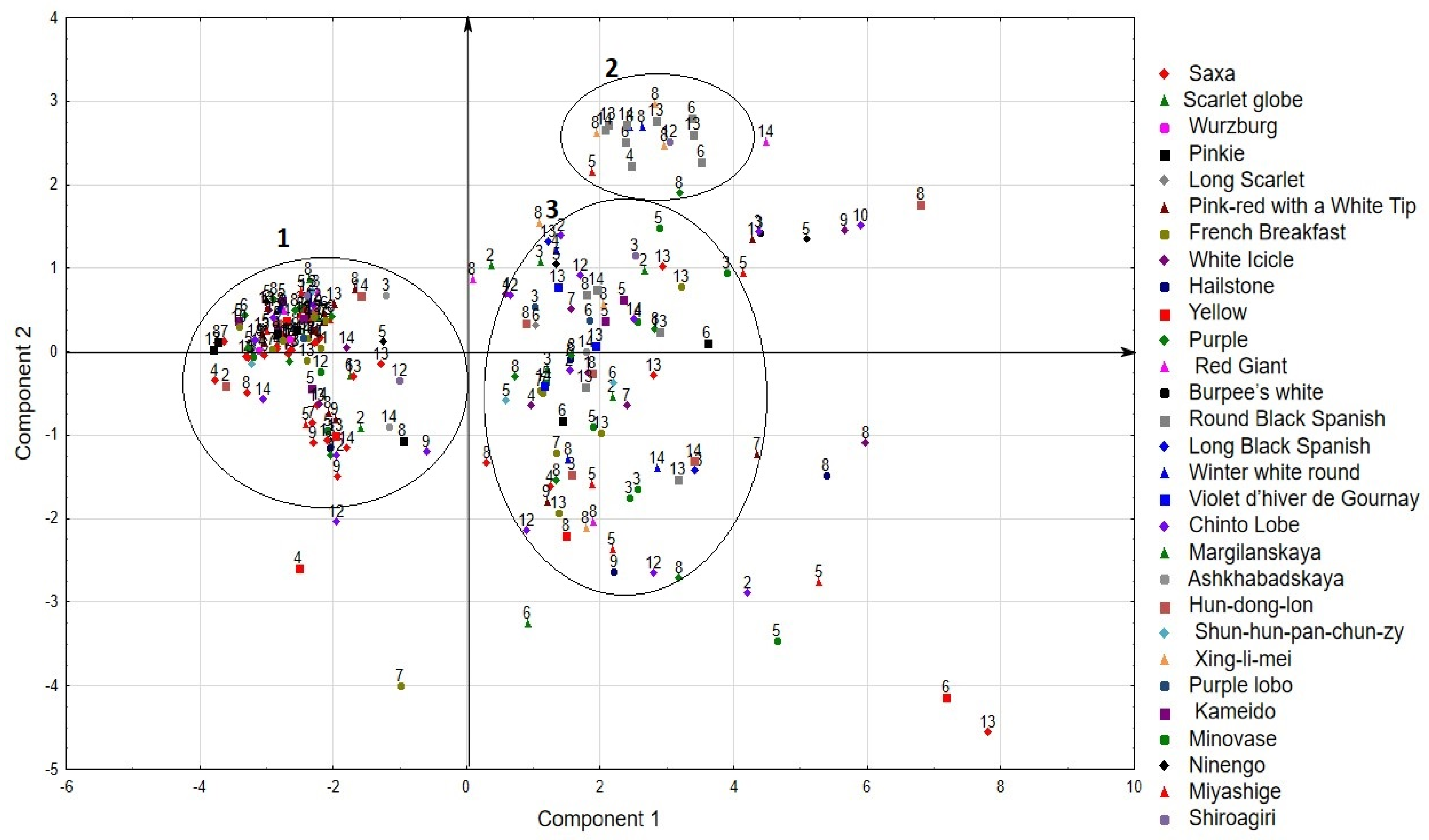
2.3. Principal Component Analysis (PCA)
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Field Evaluation and Agronomic Practices
4.3. Biochemical Analysis
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sinskaya, E.N. The oleiferous plants and root crops of the family Cruciferae. Bull. Appl. Biol. 1928, 19, 448–534. (In Russian) [Google Scholar]
- Schulz, O.E. Cruciferae. In Die Natürlichen Pflanzenfamilien, 2nd ed.; Engler, A., Prantle, K., Eds.; Engelmann: Leipzig, Germany, 1936; Volume 17, pp. 227–658. [Google Scholar]
- Gómez-Campo, C. Morphology and morphotaxonomy of the Tribe Brassiceae. In Brassica Crops and Wild Allies; Tsunoda, S., Hinata, K., Gómez-Campo, C., Eds.; Japan Scientific Societies Press: Tokyo, Japan, 1980; pp. 3–31. [Google Scholar]
- Lewis-Jones, L.J.; Thorpe, J.P.; Wallis, G.P. Genetic divergence in four species of the genus Raphanus: Implications for the ancestry of the domestic radish R. sativus. Biol. J. Linn. Soc. 1982, 18, 35–48. [Google Scholar] [CrossRef]
- Kaneko, Y.; Kimizuka-Takagi, C.; Bang, S.W.; Matsuzawa, Y. Radish. In Genome Mapping and Molecular Breeding in Plants; Kole, C., Ed.; Springer: New York, NY, USA, 2007; Volume 5, pp. 141–160. [Google Scholar]
- Vavilov, N.I. Studies on the origin of cultivated plants. Bull. Appl. Biol. 1926, 16, 139–248. (In Russian) [Google Scholar]
- Vavilov, N.I. The problems of breeding, the role of Eurasia and New World in origin of cultivated plants. In Selected Works; USSR Academy of Science Press: Moscow-Leningrad, Russia, 1960; Volume 2, p. 520. (In Russian) [Google Scholar]
- Bunin, M.S.; Esikawa, X. Genetic resources of the Japanese radish subspecies daikon and its introduction in northern regions of Eurasia. Selskokhozyaystvennaya Biol. Agric. Biol. 1993, 1, 19–32. (In Russian) [Google Scholar]
- Sazonova, L.V. Intraspecific classification of Raphanus sativus L. root crops. Bull. Appl. Biol. 1971, 15, 12–24. (In Russian) [Google Scholar]
- Sazonova, L.V.; Vlasova, E.A. Root Crops; Agropromizdat: Leningrad, Russia, 1990; p. 296. (In Russian) [Google Scholar]
- Yamagishi, H. Speciation and diversification of radish. In The Radish Genome; Nishio, T., Kitashiba, H., Eds.; Springer: Cham, Switzerland, 2017; pp. 11–30. [Google Scholar]
- Lü, N.; Yamane, K.; Ohnishi, O. Genetic diversity of cultivated and wild radish and phylogenetic relationships among Raphanus and Brassica species revealed by the analysis of trnK/matK sequence. Breed. Sci. 2008, 58, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, R.M.P.; Perez, R.L. Raphanus sativus (Radish): Their Chemistry and Biology. Sci. World J. 2004, 4, 811–837. [Google Scholar] [CrossRef]
- Ramirez, D.; Abellán-Victorio, A.; Beretta, V.; Camargo, A.; Moreno, D.A. Functional Ingredients from Brassicaceae Species: Overview and Perspectives. Int. J. Mol. Sci. 2020, 21, 1998. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, D.; Konishi, T. Anthocyanins and anthocyanin-rich extracts: Role in diabetes and eye function. Asia Pac. J. Clin. Nutr. 2007, 16, 200–208. [Google Scholar]
- Wang, L.-S.; Stoner, G.D. Anthocyanins and their role in cancer prevention. Cancer Lett. 2008, 269, 281–290. [Google Scholar] [CrossRef] [Green Version]
- Banihani, S.A. Radish (Raphanus sativus) and Diabetes. Nutrients 2017, 9, 1014. [Google Scholar] [CrossRef] [Green Version]
- Manivannan, A.; Kim, J.-H.; Kim, D.-S.; Lee, E.-S.; Lee, H.-E. Deciphering the Nutraceutical Potential of Raphanus sativus—A Comprehensive Overview. Nutrients 2019, 11, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, X.; Zhang, H.; Duan, Y.; Chen, G. Protective effects of radish (Raphanus sativus L.) leaves extract against hydrogen peroxide-induced oxidative damage in human fetal lung fibroblast (MRC-5) cells. Biomed. Pharmacother. 2018, 103, 406–414. [Google Scholar] [CrossRef]
- Nishio, T.; Sakamoto, K. Polymorphism of self-incompatibility genes. In The Radish Genome; Nishio, T., Kitashiba, H., Eds.; Springer: Cham, Switzerland, 2017; pp. 177–188. [Google Scholar]
- Food and Agriculture Organization of the United Nations, Teams on International Investment and Tropical fruits Trade and Market Division. Banana Market Review: Preliminary Results. 2019. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 11 December 2019).
- Leunov, V.I. Directions in the breeding and seed production of vegetable root crops. Potatoes Veg. 2017, 10, 6–9. (In Russian) [Google Scholar]
- Vetrova, S.A.; Vjurtts, T.S.; Zayachkovskaya, T.V.; Stepanov, V.A. Current state of the vegetable root crop market in the Russian Federation and ways to solve the problem of food security. Veg. Crop. Russ. 2020, 2, 16–22. [Google Scholar] [CrossRef]
- Yanaeva, D.A. Cultivars and hybrids of the garden radish for the growing according to cassette technology. Potato Veg. 2015, 2, 19–21. (In Russian) [Google Scholar]
- Kosenko, M.A.; Timakova, L.N. Breeding of source material for radish breeding. Potato Veg. 2020, 9, 33–36. [Google Scholar] [CrossRef]
- Fedorova, M.I.; Stepanov, V.A. Root vegetables, breeding trends, results. Veg. Crop. Russ. 2017, 4, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Shebalina, M.A.; Sazonova, L.V. Kulturnaya Flora SSSR. L.; Agropromizdat: Leningrad, Russia, 1985; pp. 156–324. (In Russian) [Google Scholar]
- Bokhan, A.I.; Yudaeva, V.E. Gene Pool and Breeding of Root Crops of the Raphanus Sativus L. (Small Radish, Radish, Daikon, Lobo); VSTISP: Мoscow, Russia, 2015; p. 134. (In Russian) [Google Scholar]
- Ermakov, A.I.; Arasimovich, V.V. Biochemistry of Vegetable Crops; Leningrad: Мoscow, Russia, 1961; p. 544. (In Russian) [Google Scholar]
- Ndjiondjop, M.-N.; Semagn, K.; Gouda, A.C.; Kpeki, S.B.; Tia, D.D.; Sow, M.; Goungoulou, A.; Sie, M.; Perrier, X.; Ghesquiere, A.; et al. Genetic Variation and Population Structure of Oryza glaberrima and Development of a Mini-Core Collection Using DArTseq. Front. Plant. Sci. 2017, 8, 1748. [Google Scholar] [CrossRef] [Green Version]
- Linde, C.C.; Selmes, H. Genetic Diversity and Mating Type Distribution of Tuber melanosporum and Their Significance to Truffle Cultivation in Artificially Planted Truffiéres in Australia. Appl. Environ. Microbiol. 2012, 78, 6534–6539. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Nevado, C.; Lam, T.Y.; Holmes, E.; Pagán, I. The impact of host genetic diversity on virus evolution and emergence. Ecol. Lett. 2017, 21, 253–263. [Google Scholar] [CrossRef]
- Vavilov, N.I. World plant resources and their use in breeding. In Mathematics and Natural Science in the USSR; Leningrad: Мoscow, Russia, 1938; pp. 575–595. (In Russian) [Google Scholar]
- Jatoi, S.A.; Siddiqui, S.U.; Masood, M.S.; Javaid, A.; Iqbal, M.; Sayal, O.U. Genetic diversity in radish germplasm for morpho-logical traits and seed storage proteins. Pak. J. Bot. 2011, 43, 2507–2512. [Google Scholar]
- Bhandari, H.R.; Bhanu, A.N.; Srivastava, K. Assessment of Genetic Diversity in Crop Plants—An Overview. Adv. Plants Agric. Res. 2017, 7, 1–8. [Google Scholar] [CrossRef]
- Singh, B.K.; Koley, T.K.; Karmakar, P.; Tripathi, A.; Singh, B.; Singh, M. Pigmented radish (Raphanus sativus): Genetic varia-bility, heritability and interrelationships of total phenolics, anthocyanins and antioxidant activity. Indian J. Agric. Sci. 2017, 87, 1600–1606. [Google Scholar]
- Savchenko, I.V. Genetic resources are the basis of food security in Russia. Hortic. Vitic. 2017, 1, 9–13. (In Russian) [Google Scholar]
- Witzel, K.; Kurina, A.B.; Artemyeva, A.M. Opening the Treasure Chest: The Current Status of Research on Brassica oleracea and B. rapa Vegetables From ex situ Germplasm Collections. Front. Plant. Sci. 2021, 12, 643047. [Google Scholar] [CrossRef] [PubMed]
- Likhenko, I.E.; Artemova, G.V.; Stepochkin, P.I. Gene pool and breeding of agricultural plants. Sib. Bull. Agric. Sci. 2014, 5, 35–41. [Google Scholar]
- Ryabchoun, V.K.; Boguslavskiy, R.L. Wheat and triticale genetic resources in Ukraine. In Cereal Genetic Resources in Europe, Proceedings of the Cereals Network, First Meeting, Yerevan, Armenia, 3–5 July 2003; International Plant Genetic Resources Institute: Rome, Italy, 2003; pp. 186–192. [Google Scholar]
- Diwan, N.; McIntosh, M.S.; Bauchan, G.R. Methods of developing a core collection of annual Medicago species. Theor. Appl. Genet. 1995, 90, 755–761. [Google Scholar] [CrossRef] [PubMed]
- Frankel, O.H. Genetic perspectives of germplasm conservation. In Genetic Manipulation: Impact on Man and Society; Arber, W., Llimensee, K., Peacock, W.J., Starlinger, P., Eds.; Cambridge University Press: Cambridge, UK, 1984. [Google Scholar]
- Goncharov, N.P.; Shumny, V.K. From preservation of genetic collections to organization of national project of plant gene pools’ conservation in permafrost. VOGiS Bull. 2008, 12, 509–523. (In Russian) [Google Scholar]
- Fokina, E.M.; Belyaeva, G.N.; Razantsvey, D.R. The trait collection of soybeans as a basis for the creation of new generation varieties. Vestn. Far. East Branch 2020, 4, 86–92. (In Russian) [Google Scholar] [CrossRef]
- Guzmán, L.F.; Machida-Hirano, R.; Borrayo, E.; Cortés-Cruz, M.; Espíndola-Barquera, M.D.C.; García, E.H. Genetic Structure and Selection of a Core Collection for Long Term Conservation of Avocado in Mexico. Front. Plant. Sci. 2017, 8, 243. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.-M.; Zhang, N.-N.; Liu, X.-J.; Yang, Z.-J.; Jia, H.-Y.; Xu, D.-P. Genetic Diversity and Population Structure Analysis of DalbergiaOdorifera Germplasm and Development of a Core Collection Using Microsatellite Markers. Genes 2019, 10, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mashkey, V.K.; Vikram, B.; Maurya, K.R. Genetic variability for quantitative and qualitative traits of radish (Raphanus sativus L.). Pharma Innov. J. 2021, 10, 636–638. [Google Scholar]
- Mapari, A.V.; Dod, V.N.; Peshattiwar, P.D.; Thorat, A. Genetic variability in radish. The Asian, J. Hort. 2009, 4, 255–258. [Google Scholar]
- Roopa, V.R.; Hadimani, H.P.; Hanchinamani, C.N.; Tatagar, M.H.; Nishani, S.; Kamble, C. Genetic variability in radish (Raphanus sativus L.). Int. J. Chem. Stud. 2018, 6, 2877–2879. [Google Scholar]
- Raihan, M.S.; Jahan, N.A. Genetic variability assessment in selected genotypes of radish (Raphanus sativus L.) using morphological markers. J. Res. Opin. 2019, 6, 2495–2501. [Google Scholar] [CrossRef]
- Muminović, J.; Merz, A.; Melchinger, A.E. Genetic structure and diversity among radish varieties as inferred from AFLP and ISSR analyses. J. Amer. Soc. Hort. Sci. 2005, 130, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Kitamoto, N.; Ohsawa, R.; Fujimura, T. Genetic diversity of radish (Raphanus sativus) germplasms and relationships among worldwide accessions analyzed with AFLP markers. Breed. Sci. 2008, 58, 107–112. [Google Scholar] [CrossRef] [Green Version]
- Ivy, N.A.; Biswas, M.S.; Rasul, G.; Hossain, T.; Mian, M.A.K. Variations of genotypes of radish at molecular level using iso-zyme analysis for the identification of self-incompatible lines. Glob. J. Biotechnol. Biochem. 2010, 5, 19–26. [Google Scholar]
- Kong, Q.; Li, X.; Xiang, C.; Wang, H.; Song, J.; Zhi, H. Genetic Diversity of Radish (Raphanus sativus L.) Germplasm Resources Revealed by AFLP and RAPD Markers. Plant. Mol. Biol. Rep. 2010, 29, 217–223. [Google Scholar] [CrossRef]
- Cruz, S.M.; Nery, M.C.; Pinho, E.V.; Luiz, M. Molecular characterization of radish cultivars. Rev. Ciênc. Agron. 2014, 45, 815–822. [Google Scholar] [CrossRef] [Green Version]
- Arro, J.; Labate, J.A. Genetic variation in a radish (Raphanus sativus L.) geodiversity collection. Genet. Resour. Crop. Evol. 2021, 1–9. [Google Scholar] [CrossRef]
- Lee, Y.J.; Mun, J.H.; Jeong, Y.M.; Joo, S.H.; Yu, H.J. Assembly of a radish core collection for evaluation and preservation of genetic diversity. Hortic. Environ. Biotechnol. 2018, 59, 711–721. [Google Scholar] [CrossRef]
- Rudakova, A.S.; Rudakov, S.V.; Artemyeva, A.M.; Kurina, A.B.; Kocherina, N.V.; Chesnokov, Y.V. Evaluation of esterase polymorphisms in mature seeds of radish (raphanus sativus l.) Accessions of vir collection. Veg. Crop. Russ. 2018, 5, 3–8. [Google Scholar] [CrossRef] [Green Version]
- Rudakova, A.S.; Rudakov, S.V.; Artemyeva, А.М.; Kurina, А.B.; Kocherina, N.V.; Chesnokov, Y.V. Polymorphism of esterase isoenzymes of ripe seeds of samples of radish (Raphanus sativus L.). Sel’skokhozyaistvennaya Biol. Agric. Biol. 2020, 55, 956–969. (In Russian) [Google Scholar]
- Kurina, A.; Artemyeva, A. Trait collection of VIR small radish (Raphanus sativus L.). In Proceedings of the Brassica 2018, 21st Crucifer Genetics Conference, Saint-Malo, France, 1–4 July 2018; p. 23. [Google Scholar]
- Kurina, A.B.; Artemeva, A.M. Trait-specific collection of Raphanus sativus L. at VIR. Book of abstracts. In Proceedings of the International Conference “125 Years of Applied Botany in Russia”, Saint-Petersburg, Russia, 25–28 November 2019; p. 155. (In Russian). [Google Scholar] [CrossRef]
- Kurina, A.B.; Khmelinskaya, T.V.; Artemyeva, A.M. Genetic diversity of vir collections of the Raphanus sativus L. (small radish and radish). Veg. Crop. Russ. 2018, 5, 9–13. [Google Scholar] [CrossRef] [Green Version]
- Kurina, A.B.; Artemyeva, A.M. Morphological and physiological variability of small radish (Raphanus sativus L.) grown un-der various conditions. Agrophysics 2020, 1, 52–58. (In Russian) [Google Scholar] [CrossRef]
- Kurina, A.B.; Kosareva, I.A.; Artemyeva, A.M. Genetic diversity of VIR Raphanus sativus L. collections on aluminum tolerance. Vavilov J. Genet. Breed. 2020, 24, 613–624. [Google Scholar] [CrossRef]
- Kurina, A.B.; Artemyeva, A.M. Molecular Genetic Analysis of VIR Radish (Raphanus Sativus L.) Collection. In Proceedings of the International Conference, Devoted to the 85 Anniversaries of the Agrophysical Research Institute «Agrophysical Trends: From Actual Challenges in Arable Farming and Crop Growing towards Advanced Technologies», Saint-Petersburg, Russia, 27–29 September 2017; St. Agrophysical Research Institute: Petersburg, Russia, 2017; p. 864. [Google Scholar]
- Sazonova, L.V. Guidelines for the Study and Maintenance of the World Collection of Root Vegetables; VIR: Leningrad, Russia, 1989; p. 88. (In Russian) [Google Scholar]
- Pavlov, L.V.; Soldatenko, A.V. Guidelines for Approbation of Vegetable and Cucurbit Crops; FSBSI FSVC: Moskov, Russia, 2018; p. 224. (In Russian) [Google Scholar]
- Ermakov, A.I.; Arasimovich, V.V.; Yarosh, N.P. Methods of Biochemical Research of Plants; Agropromiz-dat: Leningrad, Russia, 1987; p. 430. (In Russian) [Google Scholar]
- State Standard 8756. 13–87. Fruit and Vegetable Products. Methods for Determination of Sugars; Standartinform Publ.: Moscow, Russia, 2010; p. 11. (In Russian) [Google Scholar]
- State Standard 24556-89. Products of Fruits and Vegetables Processing. Methods for Determination of Vitamin C; Standartinform Publ.: Moscow, Russia, 2003; p. 11. (In Russian) [Google Scholar]
- Cattell, R.B. The Scree test for the number of factors. Multivar. Behav. Res. 1966, 1, 245–276. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type (Number of Accessions) | Duration of Vegetative Phase. Days | Rosette Height. cm | Rosette Diameter. cm | Leaf Length. cm | Leaf Width. cm | Root Length. cm | Root Diameter. cm | Root Index (Length/Diameter) | Plant Weight. g | Root Weight. g |
|---|---|---|---|---|---|---|---|---|---|---|
| R. sativus subsp. sativus convar. radicula (Pers.) Sazon. | ||||||||||
| var. rubescens Sinsk. | ||||||||||
| Saxa (44) | 22.54 ± 1.30 agj | 18.82 ± 3.34 bdefj | 15.06 ± 1.52 bdf | 10.88 ± 1.65 acdegj | 6.77 ± 1.09 aef | 3.32 ± 0.40 a | 3.01 ± 0.30 a | 1.11 ± 0.14 a | 35.48 ± 8.79 abd | 19.69 ± 4.40 abcd |
| 20.50–25.80 | 11.10–26.30 | 12.80–18.70 | 7.50–15.00 | 4.60–9.30 | 2.30–4.30 | 2.20–3.50 | 0.90–1.50 | 17.70–50.10 | 10.20–28.10 | |
| 5.80% | 17.80% | 10.10% | 15.20% | 16.10% | 12.00% | 10.10% | 12.60% | 24.80% | 22.40% | |
| Scarlet globe (5) | 22.90 ± 0.74 abegj | 17.50 ± 6.09 bdefj | 14.24 ± 1.98 bdf | 9.72 ± 2.87 bdef | 6.00 ± 1.51 bdf | 3.58 ± 0.36 a | 2.86 ± 0.40 a | 1.28 ± 0.26 a | 29.94 ± 8.95 ab | 15.82 ± 2.94 ad |
| 21.80–23.50 | 11.30–25.60 | 11.90–16.40 | 6.90–13.80 | 4.40–7.70 | 3.20–3.90 | 2.30–3.20 | 1.00–1.70 | 19.20–43.00 | 12.40–20.20 | |
| 3.20% | 34.80% | 13.90% | 29.50% | 25.10% | 10.00% | 13.90% | 20.20% | 29.90% | 18.60% | |
| Würzburg (4) | 25.48 ± 0.45 bdef | 16.89 ± 2.36 bdefj | 14.82 ± 1.28 bdf | 10.77 ± 1.17 acdegj | 7.35 ± 0.92 aeg | 3.53 ± 0.57 a | 3.07 ± 0.40 a | 1.16 ± 0.14 a | 36.33 ± 4.68 abcd | 19.93 ± 3.43 abcd |
| 24.83–25.83 | 14.32–19.90 | 13.47–16.28 | 9.47–11.94 | 6.14–8.36 | 2.72–4.06 | 2.76–3.65 | 0.99–1.33 | 31.43–41.55 | 16.23–23.22 | |
| 1.80% | 14.0% | 8.70% | 10.90% | 12.60% | 16.20% | 13.20% | 12.20% | 12.90% | 17.20% | |
| Pinkie (7) | 24.36 ± 1.14 ab | 15.60 ± 2.44 b | 14.33 ± 1.73 b | 8.79 ± 1.20 b | 5.21 ± 0.63 b | 3.31 ± 0.35 a | 3.00 ± 0.39 a | 1.11 ± 0.13 a | 32.43 ± 14.27 ab | 20.75 ± 9.69 ac |
| 23.00–26.50 | 12.98–18.80 | 12.18–17.00 | 7.40–10.21 | 4.40–6.21 | 2.92–3.81 | 2.50–3.71 | 1.01–1.40 | 21.00–58.88 | 13.80–40.79 | |
| 4.70% | 15.70% | 12.10% | 13.60% | 12.00% | 10.60% | 13.10% | 12.00% | 44.00% | 46.70% | |
| Long Scarlet (6) | 27.83 ± 1.46 ch | 27.01 ± 3.44 h | 20.84 ± 1.30 h | 13.61 ± 1.56 h | 8.95 ± 0.86 h | 9.67 ± 2.13 d | 2.10 ± 0.44 b | 4.99 ± 2.41 d | 53.31 ± 9.54 e | 26.64 ± 6.34 ce |
| 25.50–29.80 | 22.05–30.50 | 18.23–21.70 | 11.50–15.10 | 7.48–9.90 | 7.90–13.83 | 1.41–2.80 | 3.30–9.82 | 42.00–69.20 | 20.40–37.60 | |
| 5.30% | 12.70% | 6.30% | 11.50% | 9.70% | 22.10% | 21.10% | 48.30% | 17.90% | 6.30% | |
| var. striatus Sinsk. | ||||||||||
| Pink-red with a White Tip (18) | 23.38 ± 1.13 a | 19.94 ± 4.06 a | 16.91 ± 2.22 a | 10.59 ± 1.73 a | 6.83 ± 1.23 a | 3.24 ± 0.60 a | 3.00 ± 0.48 a | 1.14 ± 0.42 a | 34.54 ± 9.64 a | 19.28 ± 5.36 ac |
| 21.30–25.00 | 10.17–27.30 | 11.58–19.90 | 6.33–12.90 | 4.33–8.90 | 2.50–4.96 | 1.93–3.90 | 0.81–2.57 | 20.10–52.50 | 10.70–32.00 | |
| 4.80% | 20.40% | 13.10% | 16.30% | 17.90% | 18.50% | 16.20% | 36.50% | 27.90% | 27.80% | |
| French Breakfast (17) | 22.28 ± 1.04 g | 21.76 ± 3.04 acg | 17.52 ± 1.54 aeg | 11.23 ± 1.31 aceg | 7.42 ± 0.89 aceg | 5.61 ± 0.90 c | 2.20 ± 0.23 b | 2.59 ± 0.53 c | 33.54 ± 7.50 abd | 17.86 ± 3.34 acd |
| 20.50–24.08 | 12.83–25.53 | 13.33–19.56 | 7.33–13.06 | 4.83–8.88 | 4.25–7.92 | 1.87–2.69 | 1.66–3.84 | 19.99–43.94 | 11.09–25.03 | |
| 4.70% | 14.00% | 8.80% | 11.60% | 12.00% | 16.10% | 10.70% | 20.40% | 22.30% | 18.70% | |
| var. radicula | ||||||||||
| White Icicle (12) | 26.83 ± 2.06 c | 20.24 ± 3.15 ac | 18.34 ± 2.38 c | 10.83 ± 1.33 ac | 7.38 ± 1.24 ac | 7.71 ± 1.69 b | 2.09 ± 0.19 b | 3.78 ± 1.10 b | 40.07 ± 8.52 ac | 21.36 ± 3.64 abc |
| 22.50–30.50 | 14.17–25.80 | 13.28–21.30 | 8.44–13.20 | 5.10–9.10 | 5.30–11.56 | 1.80–2.40 | 2.20–6.19 | 25.50–52.50 | 14.50–26.96 | |
| 7.70% | 15.60% | 13.00% | 12.30% | 16.80% | 22.00% | 9.20% | 29.20% | 21.30% | 17.00% | |
| Hailstone (6) | 24.22 ± 1.00 abde | 17.34 ± 2.29 bde | 17.52 ± 2.48 acde | 10.92 ± 1.10 abcde | 6.84 ± 0.82 ace | 3.69 ± 1.09 a | 3.31 ± 1.08 a | 1.13 ± 0.09 a | 47.16 ± 23.69 acd | 27.28 ± 14.14 abc |
| 23.00–26.00 | 14.90–21.10 | 14.10–20.20 | 9.90–13.00 | 6.00–7.83 | 2.75–5.75 | 2.30–5.36 | 1.00–1.26 | 26.33–114.50 | 11.83–75.20 | |
| 4.10% | 13.20% | 14.20% | 10.10% | 12.10% | 29.40% | 32.80% | 8.20% | 41.50% | 38.50% | |
| var. chloris Alef. | ||||||||||
| Yellow (5) | 25.86 ± 0.91 bcdef | 16.81 ± 1.06 bdef | 14.72 ± 1.23 bdf | 9.24 ± 1.00 bdf | 5.96 ± 0.43 bdef | 3.46 ± 0.45 a | 3.08 ± 0.29 a | 1.13 ± 0.07 a | 34.80 ± 7.70 abcd | 21.74 ± 7.20 abc |
| 24.50–26.80 | 15.30–18.10 | 12.83–15.80 | 8.00–10.71 | 5.48–6.50 | 3.20–4.25 | 2.87–3.54 | 1.03–1.20 | 26.90–45.74 | 14.00–33.38 | |
| 3.50% | 6.30% | 8.40% | 10.80% | 7.30% | 12.90% | 9.30% | 6.20% | 22.10% | 33.10% | |
| var. violaceus | ||||||||||
| Purple (12) | 24.79 ± 0.84 bd | 17.88 ± 2.68 bd | 15.89 ± 1.58 bd | 10.13 ± 0.70 abcd | 5.98 ± 0.79 bd | 3.31 ± 0.35 a | 2.93 ± 0.27 a | 1.13 ± 0.11 a | 35.63 ± 6.40 abc | 19.93 ± 4.03 abc |
| 23.50–26.00 | 13.25–21.82 | 13.10–18.03 | 8.81–11.36 | 5.00–7.82 | 2.77–3.96 | 2.45–3.27 | 1.00–1.40 | 27.63–50.42 | 13.11–27.82 | |
| 3.4 | 15.00% | 9.90% | 6.90% | 13.20% | 10.50% | 9.30% | 9.50% | 18.00% | 20.20% | |
| R. sativus subsp. sinensis Sazon. et Stankev. convar. sinensis | ||||||||||
| var. roseus Sazon. | ||||||||||
| Red Giant (10) | 34.44 ± 1.57 i | 21.44 ± 1.86 acgi | 18.97 ± 1.48 ce | 12.88 ± 1.70 hi | 8.16 ± 0.53 c | 7.22 ± 2.07 e | 2.17 ± 0.24 b | 3.35 ± 1.01 e | 50.80 ± 18.16 de | 27.31 ± 8.79 ced |
| 32.00–37.50 | 19.60–25.60 | 16.70–21.25 | 10.90–15.20 | 7.60–9.00 | 4.20–11.50 | 1.80–2.44 | 2.10–4.80 | 24.00–94.00 | 12.50–45.63 | |
| 4.50% | 8.70% | 7.80% | 13.20% | 6.50% | 28.70% | 11.10% | 30.10% | 35.80% | 32.2% | |
| var. sinensis | ||||||||||
| Burpee’s white (3) | 34.77 ± 0.51 i | 21.43 ± 3.11 acgij | 17.60 ± 1.20 acdeg | 12.33 ± 1.02 aceghi | 7.60 ± 0.52 aceg | 3.30 ± 0.35 a | 3.07 ± 0.32 a | 1.07 ± 0.06 a | 52.17 ± 5.75 de | 28.83 ± 3.86 ced |
| 34.20–35.20 | 18.20–24.40 | 16.40–18.80 | 11.60–13.50 | 7.30–8.20 | 3.10–3.70 | 2.70–3.30 | 1.00–1.10 | 46.40–57.90 | 25.10–32.80 | |
| 1.50% | 14.50% | 6.80% | 8.30% | 6.80% | 10.50% | 10.50% | 5.40% | 11.00% | 13.40% | |
| Mean of collection (149) | 24.75 ± 3.63 | 19.51 ± 3.85 | 16.46 ± 2.39 | 10.90 ± 1.78 | 6.93 ± 1.24 | 4.48 ± 2.09 | 2.75 ± 0.55 | 1.81 ± 1.30 | 37.75 ± 12.67 | 20.86 ± 7.33 |
| 20.50–37.50 | 10.17–30.50 | 11.58–21.72 | 6.33–15.20 | 4.33–9.90 | 2.34–13.83 | 1.41–5.36 | 0.81–9.82 | 17.69–114.50 | 10.22–75.20 | |
| 14.70% | 19.70% | 14.50% | 16.30% | 17.90% | 46.70% | 19.80% | 71.60% | 33.60% | 35.20% | |
| R. sativus subsp. sativus convar. sativus | ||||||||||
| var. sativus | ||||||||||
| Odesskaya (4) | 34.88 ± 1.45 f | 28.78 ± 4.93 abd | 31.00 ± 3.36 acde | 14.30 ± 1.18 ab | 8.60 ± 0.52 de | 6.80 ± 0.74 ef | 6.65 ± 0.66 abcdef | 1.05 ± 0.13 be | 385.08 ± 90.74 ab | 230.22 ± 70.32 abcd |
| 33.80–37.00 | 21.60–32.70 | 27.10–35.00 | 12.60–15.20 | 8.20–9.30 | 6.30–7.90 | 5.80–7.30 | 0.90–1.20 | 259.10–465.00 | 146.80–312.00 | |
| 4.10% | 17.10% | 10.80% | 8.30% | 6.10% | 10.90% | 9.90% | 12.30% | 23.60% | 30.60% | |
| var. rubrus Sinsk. | ||||||||||
| Ostergrus (8) | 35.06 ± 0.73 f | 18.25 ± 2.86 be | 24.80 ± 2.86 af | 13.69 ± 1.77 ab | 8.46 ± 1.17 de | 14.25 ± 7.91 abcdg | 3.96 ± 0.96 cg | 4.07 ± 2.96 cd | 231.75 ± 108.32 b | 131.90 ± 70.54 abc |
| 34.00–36.50 | 14.29–22.50 | 21.00–29.37 | 11.25–16.92 | 6.75–10.42 | 6.14–31.38 | 2.70–4.90 | 1.32 ± 10.24 | 113.40–396.20 | 69.00–278.40 | |
| 2.10% | 15.70% | 11.50% | 13.00% | 13.90% | 55.50% | 24.20% | 52.90% | 46.70% | 53.50% | |
| convar. hybernus (Alef.) Sazon. var niger (L.) Sinsk. | ||||||||||
| Round Black Spanish (14) | 79.32 ± 9.13 e | 29.34 ± 9.38 abd | 33.72 ± 6.15 acd | 16.56 ± 3.16 ab | 11.20 ± 1.61 ab | 6.87 ± 1.12 e | 7.23 ± 0.91 d | 0.94 ± 0.09 e | 401.88 ± 153.38 ab | 258.71 ± 101.52 abc |
| 61.00–87.00 | 22.70–57.75 | 27.80–45.04 | 14.00–25.71 | 10.20–16.46 | 5.30–9.10 | 6.00–8.95 | 0.80–1.10 | 245.10–678.56 | 153.20–454.30 | |
| 11.50% | 32.00% | 18.20% | 19.10% | 14.40% | 16.40% | 12.60% | 9.10% | 38.20% | 39.20% | |
| Long Black Spanish (2) | 65.00 ± 0.00 d | 36.53 ± 3.25 acd | 42.82 ± 3.98 bd | 21.06 ± 3.74 acd | 10.19 ± 1.29 abcde | 24.48 ± 6.06 cd | 4.77 ± 0.23 abfg | 5.17 ± 1.52 cd | 573.90 ± 61.72 ab | 387.08 ± 43.50 abcd |
| 65.00–65.00 | 34.23–38.83 | 40.00–45.63 | 18.42–23.70 | 9.27–11.10 | 20.20–28.76 | 4.61–4.93 | 4.10–6.24 | 530.26–617.54 | 356.32–417.84 | |
| 0.00% | 8.90% | 9.30% | 3.70% | 12.70% | 24.80% | 4.80% | 29.40% | 10.80% | 11.20% | |
| var. hybernus | ||||||||||
| Winter white round (5) | 77.40 ± 14.02 e | 29.15 ± 9.41 abd | 34.78 ± 3.65 acd | 15.58 ± 3.93 ab | 11.25 ± 1.81 abc | 7.88 ± 0.66 bef | 7.66 ± 0.22 de | 1.02 ± 0.11 be | 475.48 ± 51.53 ab | 309.73 ± 31.06 abcd |
| 52.50–86.00 | 21.80–44.54 | 32.30–41.00 | 12.00–22.29 | 10.00–14.33 | 6.90–8.40 | 7.40–8.00 | 0.90–1.10 | 424.90–556.00 | 268.30–355.80 | |
| 18.10% | 32.30% | 10.50% | 25.20% | 16.10% | 8.30% | 2.90% | 10.50% | 10.80% | 10.00% | |
| var. violaceus | ||||||||||
| Violet d’hiver de Gournay (3) | 65.00 ± 0.00 d | 42.88 ± 6.13 c | 47.43 ± 2.39 b | 21.62 ± 2.06 c | 11.71 ± 1.35 a | 20.66 ± 4.47 abcd | 5.30 ± 0.54 abcg | 3.91 ± 0.89 abcd | 574.38 ± 201.11 ab | 361.52 ± 151.95 abcd |
| 65.00–65.00 | 36.65–48.90 | 44.74–49.33 | 19.25–22.90 | 10.55–13.19 | 15.50–23.39 | 4.86–5.91 | 3.02–4.81 | 379.88–781.49 | 211.88–515.67 | |
| 0.00% | 14.30% | 5.00% | 9.50% | 11.50% | 21.60% | 10.30% | 22.80% | 35.00% | 42.00% | |
| R. sativus subsp. sinensis convar. lobo Sazon. et Stankev. | ||||||||||
| var. lobo | ||||||||||
| Chinto Lobe (17) | 47.90 ± 7.26 a | 25.97 ± 6.97 abd | 30.03 ± 4.54 ac | 15.25 ± 2.63 ab | 10.35 ± 1.68 ade | 14.11 ± 3.02 a | 5.83 ± 1.23 a | 2.63 ± 1.06 a | 476.43 ± 113.42 a | 281.14 ± 82.07 abcd |
| 37.50–63.50 | 18.90–49.27 | 21.20–41.18 | 12.40–23.52 | 8.50–15.28 | 8.00–19.40 | 3.50–8.85 | 1.17–5.50 | 362.40–691.50 | 124.40–418.27 | |
| 15.20% | 26.80% | 15.10% | 17.30% | 16.20% | 21.40% | 21.00% | 40.30% | 23.80% | 29.20% | |
| var. virens Sazon. | ||||||||||
| Margilanskaya (8) | 47.56 ± 3.06 abc | 25.62 ± 3.67 abd | 30.27 ± 3.20 acde | 15.19 ± 2.96 ab | 10.55 ± 0.93 abc | 11.07 ± 2.70 abcfg | 5.73 ± 0.42 abcf | 1.98 ± 0.64 abc | 399.12 ± 93.44 ab | 231.68 ± 38.98 abc |
| 41.70–51.00 | 21.20–33.39 | 26.30–36.24 | 11.50–20.87 | 9.60–12.20 | 8.00–16.20 | 5.00–6.40 | 1.30–3.30 | 275.10–571.70 | 161.10–299.00 | |
| 6.40% | 14.30% | 10.60% | 19.50% | 8.90% | 24.40% | 7.30% | 32.50% | 23.40% | 16.80% | |
| var. rubidus Sazon. | ||||||||||
| Ashkhabadskaya (4) | 52.63 ± 2.95 ab | 22.80 ± 1.09 abde | 28.43 ± 1.39 acd | 13.35 ± 0.58 ab | 9.55 ± 0.40 abcde | 13.40 ± 0.81 ab | 5.93 ± 0.43 ab | 2.28 ± 0.22 ab | 377.75 ± 64.63 ab | 278.50 ± 53.55 abcd |
| 48.50–55.00 | 21.30–23.90 | 27.00–29.90 | 12.80–14.10 | 9.20–10.10 | 12.40–14.20 | 5.40–6.40 | 2.00–2.50 | 282.70–420.40 | 199.60–318.70 | |
| 5.60% | 4.80% | 4.9% | 4.40% | 4.20% | 6.10% | 7.20% | 9.80% | 17.10% | 19.20% | |
| Hun-dong-lon (9) | 47.36 ± 4.73 ac | 32.10 ± 10.92 ab | 36.20 ± 10.29 acde | 17.86 ± 5.85 abd | 11.71 ± 2.77 ab | 7.64 ± 1.02 efg | 6.94 ± 1.05 abcdf | 1.12 ± 0.26 be | 438.64 ± 160.79 ab | 251.74 ± 90.08 abc |
| 41.70–57.50 | 18.10–49.83 | 26.00–57.30 | 9.30–25.00 | 8.50–15.60 | 6.10–9.40 | 5.30–8.28 | 0.90–1.70 | 224.60–616.40 | 116.60–351.22 | |
| 10.00% | 34.00% | 28.40% | 32.80% | 23.70% | 13.40% | 15.10% | 22.90% | 36.70% | 35.80% | |
| Shun-hun-pan-chun-zy (4) | 45.08 ± 6.06 abc | 29.35 ± 5.82 abd | 31.15 ± 4.05 acde | 20.18 ± 4.72 cd | 10.78 ± 1.22 abcde | 14.40 ± 1.94 abcdfg | 5.45 ± 0.87 abcfg | 2.69 ± 0.54 abcd | 460.37 ± 207.86 ab | 321.30 ± 175.26 abcd |
| 37.50–51.50 | 23.10–35.52 | 27.10–36.56 | 16.80–27.17 | 9.40–11.85 | 11.90–16.50 | 4.26–6.25 | 2.00–3.31 | 298.60–765.25 | 210.36–582.93 | |
| 13.50% | 19.80% | 13.00% | 23.40% | 11.30% | 13.40% | 15.90% | 20.10% | 45.20% | 54.60% | |
| var. incarnatus Sazon. | ||||||||||
| Xing-li-mei (7) | 45.57 ± 8.98 abc | 24.40 ± 6.32 abde | 37.08 ± 7.57 cde | 19.13 ± 3.54 cd | 10.49 ± 2.22 abcd | 7.90 ± 2.10 bcefg | 6.27 ± 1.01 abcdef | 1.25 ± 0.17 be | 447.21 ± 84.41 ab | 230.34 ± 117.95 abd |
| 37.00–60.00 | 16.00–35.25 | 25.50–48.63 | 14.00–25.29 | 7.50–14.50 | 5.67–12.00 | 5.00–8.00 | 1.03–1.50 | 348.67–570.00 | 125.00–475.00 c | |
| 19.70% | 25.90% | 20.40% | 18.50% | 21.20% | 26.70% | 16.20% | 13.80% | 18.90% | 51.20% | |
| var. syringeus Sazon. | ||||||||||
| Purple (3) | 48.00 ± 17.09 abc | 23.72 ± 3.87 abde | 27.51 ± 4.90 acd | 16.90 ± 1.13 abc | 9.51 ± 0.76 ade | 16.10 ± 7.71 abc | 5.03 ± 1.80 abcg | 3.75 ± 2.35 abc | 352.65 ± 69.64 ab | 198.60 ± 77.19 abcd |
| 30.00–64.00 | 21.00–28.15 | 23.67–33.03 | 16.17–18.21 | 8.83–10.33 | 7.40–22.10 | 3.80–7.10 | 1.04–5.26 | 309.60–433.00 | 109.80–249.60 | |
| 35.60% | 16.30% | 17.80% | 6.70% | 8.00% | 47.90% | 35.80% | 42.70% | 19.80% | 38.90% | |
| R. sativus subsp. acanthiformis (Blanch.) Stankev. convar. minowase (Kitam.) Sazon. | ||||||||||
| var. minowase | ||||||||||
| Kameido (8) | 45.33 ± 3.83 ac | 24.94 ± 3.86 abd | 29.68 ± 4.82 acdef | 15.26 ± 3.05 ab | 9.69 ± 1.44 acde | 17.09 ± 3.91 bcd | 5.75 ± 1.15 abcf | 3.17 ± 1.22 abcd | 499.26 ± 129.07 a | 323.30 ± 102.27 acd |
| 39.00–51.00 | 18.40–31.50 | 24.30–39.61 | 12.30–20.47 | 7.90–12.73 | 11.42–22.90 | 3.90–8.02 | 1.42–5.50 | 354.70–722.95 | 176.90–501.86 | |
| 8.40% | 15.50% | 16.25% | 20.00% | 14.80% | 22.90% | 20.00% | 38.70% | 25.90% | 31.60% | |
| Minovase (13) | 44.54 ± 5.34 ac | 27.37 ± 4.10 abd | 32.15 ± 5.77 acde | 17.38 ± 4.30 abcd | 10.76 ± 1.99 abc | 22.82 ± 7.81 cd | 5.13 ± 0.76 bcg | 4.56 ± 1.39 cd | 616.90 ± 334.28 a | 439.54 ± 279.39 d |
| 36.25–56.00 | 20.60–36.75 | 24.30–43.88 | 10.00–28.00 | 8.30–14.76 | 13.30–37.64 | 3.90–6.20 | 2.10–6.32 | 324.00–1370.50 | 156.90–1057.13 | |
| 12.00% | 15.00% | 17.90% | 24.80% | 18.50% | 34.20% | 14.80% | 30.50% | 54.20% | 63.60% | |
| Ninengo (4) | 47.30 ± 1.41 abc | 25.03 ± 7.03 abd | 27.83 ± 5.82 acdef | 14.65 ± 3.17 abcd | 9.63 ± 1.21 abcde | 23.28 ± 2.39 cd | 4.45 ± 0.98 bcg | 5.58 ± 1.73 cd | 501.90 ± 82.57 a | 264.48 ± 133.86 abcd |
| 46.00–49.00 | 19.00–34.30 | 22.30–35.80 | 11.50–18.50 | 8.10–10.90 | 21.00–26.40 | 3.00–5.20 | 4.10–8.00 | 422.70–615.30 | 80.50–400.70 | |
| 3.00% | 28.10% | 20.90% | 21.60% | 12.60% | 10.30% | 22.10% | 31.10% | 16.50% | 50.60% | |
| convar. acanthiformis | ||||||||||
| Miyashige (11) | 45.84 ± 5.21 ac | 24.47 ± 3.84 abd | 27.12 ± 2.86 aef | 14.58 ± 2.94 ab | 9.68 ± 1.25 acde | 18.72 ± 5.03 bcd | 5.12 ± 0.76 abc | 3.79 ± 1.22 cd | 444.96 ± 169.67 ab | 297.25 ± 116.94 acd |
| 39.00–57.00 | 19.30–30.90 | 22.00–31.40 | 11.90–20.75 | 7.80–12.1 | 10.70–26.93 | 3.80–6.60 | 1.80–5.30 | 316.00–892.90 | 167.90–587.70 | |
| 11.40% | 15.70% | 10.50% | 20.20% | 12.90% | 26.90% | 14.80% | 32.10% | 38.10% | 39.30% | |
| Shiroagiri (5) | 41.25 ± 4.19 c | 25.62 ± 2.76 abd | 30.15 ± 0.93 acdef | 15.76 ± 3.23 ab | 9.24 ± 0.91 de | 14.86 ± 4.35 abcd | 5.75 ± 0.53 abcf | 2.61 ± 0.97 abcd | 453.38 ± 83.97 a | 323.39 ± 93.33 ad |
| 35.25–44.90 | 21.90–29.20 | 29.40–31.50 | 13.70–21.49 | 8.20–10.61 | 8.30–19.32 | 4.87–6.30 | 1.40–3.97 | 313.20–512.60 | 170.50–421.90 | |
| 10.20% | 10.80% | 3.10% | 20.50% | 9.90% | 29.30% | 9.20% | 37.10% | 18.50% | 28.90% | |
| Mean of collection (129) | 50.84 ± 14.28 | 26.0 ± 7.27 | 31.56 ± 6.53 | 16.20 ± 3.70 | 10.30 ± 1.76 | 14.00 ± 6.81 | 5.82 ± 1.27 | 2.71 ± 1.76 | 451.42 ± 175.82 | 284.93 ± 140.19 |
| 30.00–87.00 | 14.29–57.75 | 21.00–57.30 | 9.30–28.00 | 6.75–16.46 | 5.30–37.64 | 2.70–8.95 | 0.80–10.24 | 113.40–1370.50 | 69.00–1057.19 | |
| 28.10% | 27.10% | 20.70% | 22.80% | 17.10% | 48.60% | 21.90% | 65.20% | 39.00% | 49.20% | |
| Trait | Small Radish | Radish | |||||
|---|---|---|---|---|---|---|---|
| subsp. sativus | subsp. sinensis | Mean | subsp. sativus | subsp. sinensis | subsp. acanthiformis | Mean | |
| Dry matter. % | 5.59 ± 1.19 a | 7.04 ± 0.57 b | 5.67 ± 1.21 * | 10.44 ± 1.63 a | 7.90 ± 1.81 b | 7.06 ± 0.99 c | 8.14 ± 1.96 ** |
| 3.72–8.56 | 6.52–8.24 | 3.72–8.56 | 7.50–13.52 | 5.28–13.64 | 5.56–10.52 | 5.28–13.64 | |
| 21.34% | 8.09% | 21.36% | 15.57% | 22.89% | 14.00 | 24.10% | |
| Ascorbic acid. mg/100 g | 30.58 ± 6.20 a | 29.41 ± 8.30 a | 30.47 ± 6.29 * | 37.78 ± 14.58 a | 41.84 ± 16.51 a | 44.71 ± 14.82 a | 42.02 ± 15.64 ** |
| 19.04–57.72 | 24.48–48.10 | 19.04–57.72 | 14.00–56.40 | 15.00–73.32 | 11.80–62.04 | 11.80–73.32 | |
| 20.27% | 28.24% | 20.63% | 38.59% | 39.69% | 33.14% | 37.22% | |
| Monosaccharides. % | 1.83 ± 0.99 a | 2.97 ± 1.25 b | 1.89 ± 1.04 * | 2.48 ± 0.50 a | 1.95 ± 0.67 ab | 1.80 ± 0.58 b | 2.00 ± 0.65 * |
| 0.07–4.35 | 1.38–4.64 | 0.07–4.64 | 1.50–3.02 | 0.85–3.18 | 1.00–2.61 | 0.85–3.18 | |
| 54.11% | 42.25% | 54.89% | 20.30% | 34.39% | 32.09% | 32.37% | |
| Total sugar content. % | 2.06 ± 1.06 a | 3.45 ± 1.54 b | 2.14 ± 1.14 * | 2.93 ± 0.38 a | 2.27 ± 0.82 ab | 2.11 ± 0.78 b | 2.34 ± 0.79 * |
| 0.16–5.00 | 1.60–5.45 | 0.16–5.45 | 2.47–3.59 | 1.10–4.31 | 1.03–3.39 | 1.03–4.31 | |
| 51.46% | 44.66% | 53.20% | 13.08% | 36.03% | 36.84% | 33.63% | |
| Variable | Factor Loadings (Varimax Raw) Extraction: Principal Components | |||
|---|---|---|---|---|
| PC 1 | PC 2 | PC 3 | PC 4 | |
| Rosette height | 0.84 | 0.10 | −0.19 | 0.20 |
| Rosette diameter | 0.85 | 0.19 | 0.03 | 0.40 |
| Leaf length | 0.88 | 0.20 | −0.20 | 0.24 |
| Leaf width | 0.86 | 0.17 | −0.01 | 0.32 |
| Root length | 0.49 | 0.83 | 0.19 | 0.06 |
| Root diameter | 0.80 | −0.13 | 0.35 | 0.40 |
| Root index | 0.05 | 0.98 | −0.05 | 0.01 |
| Plant weight | 0.83 | 0.40 | 0.19 | 0.21 |
| Root weight | 0.83 | 0.38 | 0.20 | 0.15 |
| Duration of vegetative phase | 0.54 | 0.07 | 0.32 | 0.70 |
| Dry matter | 0.31 | 0.03 | 0.05 | 0.93 |
| Ascorbic acid | −0.02 | 0.08 | 0.94 | 0.13 |
| Expl.Var | 5.61 | 2.10 | 1.31 | 1.96 |
| Prp.Totl | 0.47 | 0.17 | 0.11 | 0.16 |
| % total variance | 61.32 | 14.14 | 10.88 | 5.17 |
| % cumulative proportion of variance | 61.32 | 75.46 | 86.34 | 91.51 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurina, A.B.; Kornyukhin, D.L.; Solovyeva, A.E.; Artemyeva, A.M. Genetic Diversity of Phenotypic and Biochemical Traits in VIR Radish (Raphanus sativus L.) Germplasm Collection. Plants 2021, 10, 1799. https://doi.org/10.3390/plants10091799
Kurina AB, Kornyukhin DL, Solovyeva AE, Artemyeva AM. Genetic Diversity of Phenotypic and Biochemical Traits in VIR Radish (Raphanus sativus L.) Germplasm Collection. Plants. 2021; 10(9):1799. https://doi.org/10.3390/plants10091799
Chicago/Turabian StyleKurina, Anastasia B., Dmitry L. Kornyukhin, Alla E. Solovyeva, and Anna M. Artemyeva. 2021. "Genetic Diversity of Phenotypic and Biochemical Traits in VIR Radish (Raphanus sativus L.) Germplasm Collection" Plants 10, no. 9: 1799. https://doi.org/10.3390/plants10091799
APA StyleKurina, A. B., Kornyukhin, D. L., Solovyeva, A. E., & Artemyeva, A. M. (2021). Genetic Diversity of Phenotypic and Biochemical Traits in VIR Radish (Raphanus sativus L.) Germplasm Collection. Plants, 10(9), 1799. https://doi.org/10.3390/plants10091799