
Phylogenetic Diversity of Wetland Plants across China



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
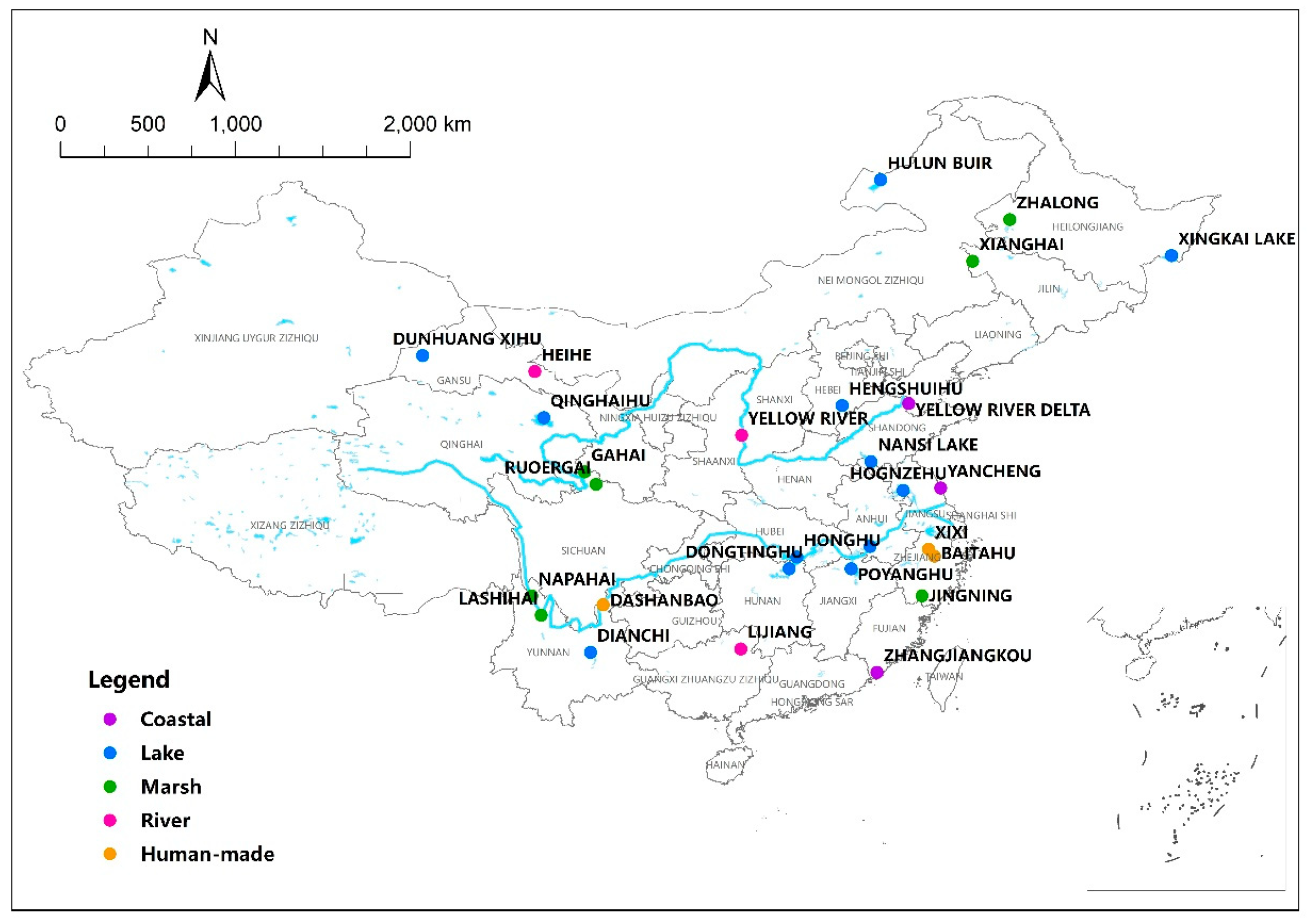
2.1. Study Site and Data Collection
2.2. Phylogenetic Diversity
3. Results
4. Discussion
4.1. Perennial Herbs
4.2. Phylogenetic Diversity
4.3. Suggestions on Wetland Restorations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hu, S.; Niu, Z.; Chen, Y.; Li, L.; Zhang, H. Global wetlands: Potential distribution, wetland loss, and status. Sci. Total Environ. 2017, 586, 319–327. [Google Scholar] [CrossRef]
- Mitra, S.; Wassmann, R.; Vlek, P. Global Inventory of Wetlands and Their Role in the Carbon Cycle; ZEF—Discussion Papers on Development Policy No. 64; Center for Development Research: Bonn, Germany, 2003; p. 44. [Google Scholar]
- Millennium Ecosystem Assessment, Ecosystems and Human Well Being: Wetlands and Water Synthesis; Water Resources Institute: Washington, DC, USA, 2005.
- Russi, D.; ten Brink, P.; Farmer, A.; Badura, T.; Coates, D.; Förster, J.; Kumar, R.; Davidson, N. The Economics of Ecosystems and Biodiversity for Water and Wetlands; IEEP: London, UK; Brussels, Belgium; Ramsar Secretariat: Gland, Switzerland, 2013. [Google Scholar]
- McNicol, G.; Sturtevant, C.S.; Knox, S.H.; Dronova, I.; Baldocchi, D.D.; Silver, W.L. Effects of seasonality, transport pathway, and spatial structure on greenhouse gasfluxes in a restored wetland. Glob. Chang. Biol. 2017, 23, 2768–2782. [Google Scholar] [CrossRef] [Green Version]
- Gong, P.; Niu, Z.G.; Cheng, X.; Zhao, K.Y.; Zhou, D.M.; Guo, J.H.; Liang, L.; Wang, X.F.; Li, D.D.; Huang, H.B.; et al. China’s wetland change (1990–2000) determined by remote sensing. Sci. China Earth Sci. 2010, 53, 1036–1042. [Google Scholar] [CrossRef]
- Davidson, N.C. How much wetland has the world lost? Long-term and recent trends in global wetland area. Mar. Freshw. Res. 2014, 65, 934–941. [Google Scholar] [CrossRef]
- Decleer, K.; Wouters, J.; Jacobs, S.; Staes, J.; Spanhove, T.; Meire, P.; Diggelen, R.V. Mapping wetland loss and restoration potential in Flanders (Belgium): An ecosystem service perspective. Ecol. Soc. 2016, 21, 46. [Google Scholar] [CrossRef] [Green Version]
- Yan, F.; Zhang, S. Ecosystem service decline in response to wetland loss in the Sanjiang Plain, Northeast China. Ecol. Eng. 2019, 130, 117–121. [Google Scholar] [CrossRef]
- Crooks, S.; Sutton-Grier, A.E.; Troxler, T.G.; Herold, N.; Wirth, T. Coastal wetland management as a contribution to the us national greenhouse gas inventory. Nat. Clim. Chang. 2018, 8, 1109–1112. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, C.M.; Ervin, G.N.; DiOrio, E.W. Interplay of water quality and vegetation in restored wetland plant assemblages from an agricultural landscape. Ecol. Eng. 2017, 108, 255–262. [Google Scholar] [CrossRef]
- Waltham, N.J.; Burrows, D.; Wegscheidl, C.; Buelow, C.; Ronan, M.; Connolly, N.; Groves, P.; Marie-Audas, D.; Creighton, C.; Sheaves, M. Lost Floodplain Wetland Environments and Efforts to Restore Connectivity, Habitat, and Water Quality Settings on the Great Barrier Reef. Front. Mar. Sci. 2019, 6, 71. [Google Scholar] [CrossRef]
- Moreno-Mateos, D.; Meli, P.; Vara-Rodríguez, M.I.; Aronson, J. Ecosystem response to interventions: Lessons from restored and created wetland ecosystems. J. Appl. Ecol. 2015, 52, 1528–1537. [Google Scholar] [CrossRef]
- Moreno-Mateos, D.; Power, M.E.; Comín, F.A.; Yockteng, R. Structural and functional loss in restored wetland ecosystems. PLoS Biol. 2012, 10, e1001247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- State Forestry Bureau. China Wetland resources: Master Volume; China Forestry Publishing House: Beijing, China, 2015. (In Chinese)
- Niu, C.; Lou, A.; Sun, R.; Li, Q. Foundations in Ecology; Higher Education Press: Beijing, China, 2002. (In Chinese) [Google Scholar]
- Swenson, N.G. The role of evolutionary processes in producing biodiversity patterns, and the interrelationships between taxonomic, functional and phylogenetic biodiversity. Am. J. Bot. 2011, 98, 472–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swenson, N.G.; Worthy, S.J. Phylogenetic Resolution and Metrics of Biodiversity and Signal in Conservation. In Phylogenetic Diversity; Scherson, R., Faith, D., Eds.; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Jin, Y.; Qian, H. V.PhyloMaker: An R package that can generate very large phylogenies for vascular plants. Ecography 2019, 42, 1353–1359. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 3 July 2021).
- Faith, D.P. Conservation evaluation and phylogenetic diversity. Biol. Conserv. 1992, 61, 1–10. [Google Scholar] [CrossRef]
- Webb, C.O.; Ackerly, D.D.; McPeek, M.A.; Donoghue, M.J. Phylogenies and community ecology. Annu. Rev. Ecol. Syst. 2002, 33, 475–505. [Google Scholar] [CrossRef] [Green Version]
- Tsirogiannis, C.; Sandel, B. PhyloMeasures: Fast and Exact Algorithms for Computing Phylogenetic Biodiversity Measures. R Package Version 2.1. 2017. Available online: https://CRAN.R-project.org/package=PhyloMeasures (accessed on 3 July 2021).
- Laliberté, E.; Legendre, P.; Shipley, B. FD: Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology. R Package Version 1.0-12. 2014. Available online: https://mran.microsoft.com/snapshot/2014-11-17/web/packages/FD/FD.pdf (accessed on 3 July 2021).
- Kembel, S.W.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef] [Green Version]
- Silk, J.W.F.; Franklin, J.; Arroyo-Rodríguez, V.; Field, R.; Aguilar, S.; Aguirre, N.; Ahumada, J.; Aiba, S.-I.; Alves, L.F.; K, A.; et al. Phylogenetic classification of the world’s tropical forests. Proc. Natl. Acad. Sci. USA 2018, 155, 1837–1842. [Google Scholar] [CrossRef] [Green Version]
- Yu, H. The Flora, Distribution Pattern, Diffusion pathway of Alien Species of Aquatic palnts in China. Doctoral Degree Dissertation, Wuhan University, Wuhan, China, 2017. (In Chinese). [Google Scholar]
- O’Connell, J.L.; Johnson, L.A.; Beas, B.J.; Smith, L.M.; Haukos, D.A. Predicting dispersal-limitation in plants: Optimizing planting decisions for isolated wetland restoration in agricultural landscapes. Biol. Conserv. 2013, 159, 343–354. [Google Scholar] [CrossRef]
- Houlahan, J.E.; Keddy, P.A.; Makkay, K.; Findlay, C.S. The effects of adjacent land use on wetland species richness and community composition. Wetlands 2006, 26, 79–96. [Google Scholar] [CrossRef]
- Zhao, K. Marshes in China; Science Press: Beijing, China, 1999. (In Chinese) [Google Scholar]
- Zhao, L.; Duan, K.; Zhao, C.; Wang, J.; Wen, J. Responses of phylogenetic structure of plant to different habitats in Jiayuguan Caohu Wetland. Chin. J. Ecol. 2020, 39, 2123–2130. (In Chinese) [Google Scholar]
- Qian, H.; Deng, T.; Jin, T.; Mao, L.; Ricklefs, R.E. Phylogenetic dispersion and diversity in regional assemblages of seed plants in china. Proc. Natl. Acad. Sci. USA 2019, 116, 23192–23201. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Shi, W.; Zhou, M. Destruction of 3 million Populus × euramericana in the Dongting Lake. Ctry. Agric. Farmer 2018, 2A, 33–35. (In Chinese) [Google Scholar]
- Divíšek, J.; Chytrý, M.; Beckage, B.; Gotelli, N.J.; Lososová, Z.; Pyšek, P.; Richardson, D.M.; Molofsky, J. Similarity of introduced plant species to native ones facilitates naturalization, but differences enhance invasion success. Nat. Commun. 2018, 9, 4631. [Google Scholar] [CrossRef]
- Ministry of Ecology and Environment of the People’s Republic of China. List of Alien Invasive Species in China. 2003. Available online: https://www.mee.gov.cn/ (accessed on 1 January 2003).
- Lu, C.; Liao, B. Consideration on Ecological Function of Alien Mangrove Plants Sonneratia apetala and Laguncularia racemosa. Wetl. Sci. 2019, 17, 682–688. (In Chinese) [Google Scholar]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, A.; Ying, Z.; Hu, X.; Yu, M. Phylogenetic Diversity of Wetland Plants across China. Plants 2021, 10, 1850. https://doi.org/10.3390/plants10091850
Zhang A, Ying Z, Hu X, Yu M. Phylogenetic Diversity of Wetland Plants across China. Plants. 2021; 10(9):1850. https://doi.org/10.3390/plants10091850
Chicago/Turabian StyleZhang, Aiying, Zhixia Ying, Xunyu Hu, and Mingjian Yu. 2021. "Phylogenetic Diversity of Wetland Plants across China" Plants 10, no. 9: 1850. https://doi.org/10.3390/plants10091850
APA StyleZhang, A., Ying, Z., Hu, X., & Yu, M. (2021). Phylogenetic Diversity of Wetland Plants across China. Plants, 10(9), 1850. https://doi.org/10.3390/plants10091850