
Rapid On-Farm Testing of Resistance in Lolium rigidum to Key Pre- and Post-Emergence Herbicides

 ,
,  ,
,
Abstract
:
1. Introduction
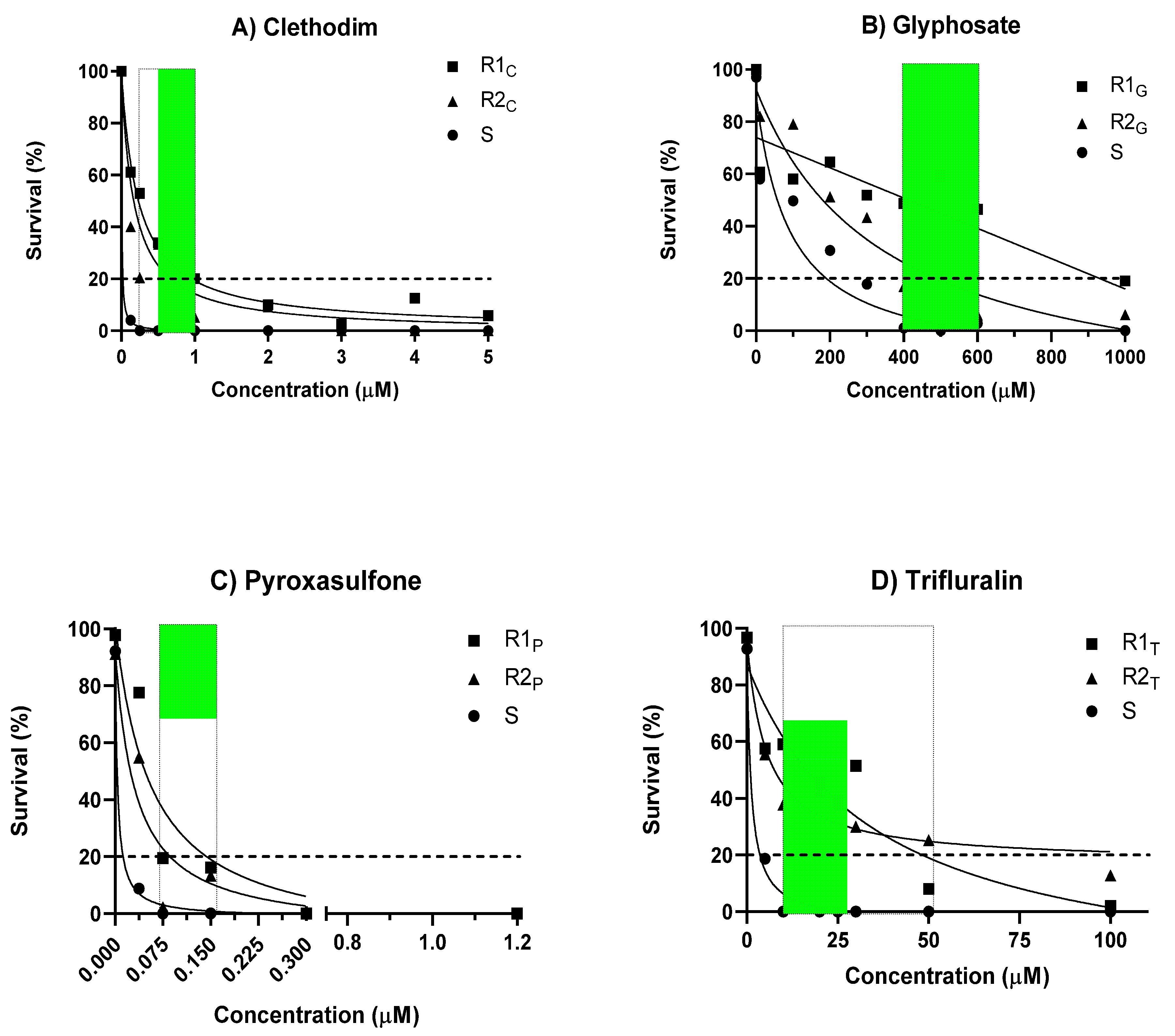
2. Results
2.1. Development of the Agar Test and Comparison with the Pot Test
2.2. Validation of the Agar Test with Field Populations
2.3. Interaction between Seed Dormancy and the Agar Test
2.4. Stability of Herbicides in Stock Solutions and Agar Plates
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Optimization of the Agar Test
4.3. Pot-Based Dose-Response Experiments
4.4. Validation of the Agar Test with Uncharacterized Populations
4.5. Investigation of Interference from Seed Dormancy
4.6. Herbicide Stability in Agar
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kloot, P.M. The genus Lolium in Australia. Aust. J. Bot. 1983, 31, 421–435. [Google Scholar] [CrossRef]
- Smith, D.F.; Levick, G.R.T. The effect of infestation by Lolium rigidum Gaud. (annual ryegrass) on the yield of wheat. Aust. J. Agric. Res. 1974, 25, 381–393. [Google Scholar] [CrossRef]
- Llewellyn, R.S.; Ronning, D.; Ouzman, J.; Walker, S.; Mayfield, A.; Clarke, M. Impact of weeds on Australian grain production: The cost of weed to Australian grain growers and the adoption of weed management and tillage practices. In Report for GRDC; CSIRO: Canberra, Australia, 2016; p. 112. [Google Scholar]
- D’Emden, F.H.; Llewellyn, R.S. No-tillage adoption decisions in southern Australian cropping and the role of weed management. Aust. J. Exp. Agric. 2006, 46, 563–569. [Google Scholar] [CrossRef]
- Kreiner, J.M.; Stinchcombe, J.R.; Wright, S.I. Population genomics of herbicide resistance: Adaptation via evolutionary rescue. Ann. Rev. Plant Biol. 2018, 69, 611–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broster, J.C.; Pratley, J.E.; Ip, R.H.L.; Ang, L.; Seng, K.P. A quarter of a century of monitoring herbicide resistance in Lolium rigidum in Australia. Crop Pasture Sci. 2019, 70, 283–293. [Google Scholar] [CrossRef]
- Heap, I.M. International Herbicide-Resistant Weed Database. 2021. Available online: http://www.weedscience.org (accessed on 15 July 2021).
- Owen, M.J.; Martinez, N.J.; Powles, S.B. Multiple herbicide-resistant Lolium rigidum (annual ryegrass) now dominates across the Western Australian grain belt. Weed Res. 2014, 54, 314–324. [Google Scholar] [CrossRef]
- Somerville, G.J.; Powles, S.B.; Walsh, M.J.; Renton, M. Why was resistance to shorter-acting pre-emergence herbicides slower to evolve? Pest Manag. Sci. 2017, 73, 844–851. [Google Scholar] [CrossRef]
- Beckie, H.J.; Heap, I.M.; Smeda, R.J.; Hall, L.M. Screening for herbicide resistance in weeds. Weed Technol. 2000, 14, 428–445. [Google Scholar] [CrossRef]
- Norsworthy, J.K.; Ward, S.M.; Shaw, D.R.; Llewellyn, R.S.; Nichols, R.L.; Webster, T.M.; Bradley, K.W.; Frisvold, G.; Powles, S.B.; Burgos, N.R. Reducing the risks of herbicide resistance: Best management practices and recommendations. Weed Sci. 2012, 60, 31–62. [Google Scholar] [CrossRef] [Green Version]
- Kaundun, S.S.; Hutchings, S.J.; Dale, R.P.; Bailly, G.C.; Glanfield, P. Syngenta ‘RISQ’ test: A novel in-season method for detecting resistance to post-emergence ACCase and ALS-inhibitor herbicides in grass weeds. Weed Res. 2011, 51, 284–293. [Google Scholar] [CrossRef]
- Burgos, N.R. Whole-plant and seed bioassays for resistance confirmation. Weed Sci. 2015, 63, 152–165. [Google Scholar] [CrossRef] [Green Version]
- Burgos, N.R.; Tranel, P.J.; Streibig, J.C.; Davis, V.M.; Shaner, D.; Norsworthy, J.K.; Ritz, C. Confirmation of resistance to herbicides and evaluation of resistance levels. Weed Sci. 2013, 61, 4–20. [Google Scholar] [CrossRef]
- Busi, R.; Beckie, H.J. Are herbicide mixtures immune to resistance? A case study with Lolium rigidum. Weed Res. 2020, 61, 92–99. [Google Scholar]
- Owen, M.; Powles, S.B. Current levels of herbicide resistance in key weed species in the WA grain belt. In Proceedings of the GRDC Grains Research Update, Perth, Australia, 26–27 February 2018. [Google Scholar]
- Moss, S.R. Techniques for determining herbicide resistance. In Proceedings of the British Crop Protection Conference—Weeds, Brighton, UK, 20–23 November 1995; pp. 547–556. [Google Scholar]
- Carringer, R.D.; Weber, J.B.; Monaco, T.J. Adsorption-desorption of selected pesticides by organic matter and montmorillonite. J. Agric. Food Chem. 1975, 23, 568–572. [Google Scholar] [CrossRef]
- Peter, C.J.; Weber, J.B. Adsorption and efficacy of trifluralin and butralin as influenced by soil properties. Weed Sci. 1985, 33, 861–867. [Google Scholar] [CrossRef]
- Brand, R.M.; Mueller, C. Transdermal penetration of atrazine, alachlor, and trifluralin: Effect of formulation. Toxicol. Sci. 2002, 68, 18–23. [Google Scholar] [CrossRef] [Green Version]
- Busi, R.; Dayan, F.E.; Francis, I.; Goggin, D.; Lerchl, J.; Porri, A.; Powles, S.B.; Sun, C.; Beckie, H.J. Cinmethylin controls multiple herbicide-resistant Lolium rigidum and its wheat selectivity is P450-based. Pest Manag. Sci. 2020, 76, 2601–2608. [Google Scholar] [CrossRef] [PubMed]
- Steadman, K.J. Dormancy release during hydrated storage in Lolium rigidum seeds is dependent on temperature, light quality, and hydration status. J. Exp. Bot. 2004, 55, 929–937. [Google Scholar] [CrossRef]
- Goggin, D.E.; Emery, R.J.N.; Powles, S.B.; Steadman, K.J. Initial characterisation of low and high seed dormancy populations of Lolium rigidum produced by repeated selection. J. Plant Physiol. 2010, 167, 1282–1288. [Google Scholar] [CrossRef] [PubMed]
- Neve, P.; Powles, S. Recurrent selection with reduced herbicide rates results in the rapid evolution of herbicide resistance in Lolium rigidum. Theor. Appl. Genet. 2005, 110, 1154–1166. [Google Scholar] [CrossRef] [PubMed]
- Owen, M.J.; Walsh, M.J.; Llewellyn, R.S.; Powles, S.B. Widespread occurrence of multiple herbicide resistance in Western Australian annual ryegrass (Lolium rigidum) populations. Aust. J. Agric. Res. 2007, 58, 711–718. [Google Scholar] [CrossRef]
- Yu, Q.; Cairns, A.; Powles, S.B. Glyphosate, paraquat and ACCase multiple herbicide resistance evolved in a Lolium rigidum biotype. Planta 2007, 225, 499–513. [Google Scholar] [CrossRef] [PubMed]
- Lorraine-Colwill, D.F.; Powles, S.B.; Hawkes, T.R.; Hollinshead, P.H.; Warner, S.A.J.; Preston, C. Investigations into the mechanisms of glyphosate resistance in Lolium rigidum. Pestic. Biochem. Physiol. 2003, 74, 62–72. [Google Scholar] [CrossRef]
- Busi, R.; Powles, S.B. Cross-resistance to prosulfocarb + S-metolachlor and pyroxasulfone selected by either herbicide in Lolium rigidum. Pest Manag. Sci. 2016, 72, 1664–1672. [Google Scholar] [CrossRef]
- Heap, I.; Knight, R.A. population of ryegrass tolerant to the herbicide diclofop-methyl. J. Aust. Inst. Agric. Sci. 1982, 48, 156–157. [Google Scholar]
- Chen, J.; Yu, Q.; Owen, M.; Han, H.; Powles, S. Dinitroaniline herbicide resistance in a multiple-resistant Lolium rigidum population. Pest Manag. Sci. 2018, 74, 925–932. [Google Scholar] [CrossRef] [PubMed]
- Busi, R.; Gaines, T.A.; Walsh, M.J.; Powles, S.B. Understanding the potential for resistance evolution to the new herbicide pyroxasulfone: Field selection at high doses vs. recurrent selection at low doses. Weed Res. 2012, 52, 489–499. [Google Scholar] [CrossRef]
- McAlister, F.M.; Holtum, J.A.M.; Powles, S.B. Dinitroaniline herbicide resistance in rigid ryegrass (Lolium rigidum). Weed Sci. 1995, 43, 55–62. [Google Scholar] [CrossRef]
- Powles, S.B.; Lorraine-Colwill, D.F.; Dellow, J.J.; Preston, C. Evolved resistance to glyphosate in rigid ryegrass (Lolium rigidum) in Australia. Weed Sci. 1998, 46, 604–607. [Google Scholar] [CrossRef]
- Yu, Q.; Collavo, A.; Zheng, M.Q.; Owen, M.; Sattin, M.; Powles, S.B. Diversity of acetyl-coenzyme A carboxylase mutations in resistant Lolium populations: Evaluation using clethodim. Plant Physiol. 2007, 145, 547–558. [Google Scholar] [CrossRef] [Green Version]
- Sandin-España, P.; Sevilla-Moran, B.; López-Goti, C.; Mateo-Miranda, M.M.; Alonso-Prados, J.L. Rapid photodegradation of clethodim and sethoxydim herbicides in soil and plant surface model systems. Arab. J. Chem. 2016, 9, 694–703. [Google Scholar] [CrossRef] [Green Version]
- Ibáñez, M.; Pozo, Ó.J.; Sancho, J.V.; López, F.J.; Hernández, F. Residue determination of glyphosate, glufosinate and aminomethylphosphonic acid in water and soil samples by liquid chromatography coupled to electrospray tandem mass spectrometry. J. Chrom. A 2005, 1081, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Ritz, C.; Streibig, J.C. Bioassay analysis using R. J. Stat. Softw. 2005, 12. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Herbicide | Population | Agar Test (µM) | Pot Test (g ha−1) | ||
|---|---|---|---|---|---|
| LD50 (95% CI) | RI | LD50 (95% CI) 1 | RI | ||
| Clethodim | R1C | 0.23 (0.13–0.43) | 4.6 | >750 | >17 |
| Clethodim | R2C | 0.18 (0.05–0.76) | 3.6 | >750 | >17 |
| Clethodim | S | 0.05 (0.004–0.007) | -- | 45 (21–68) | -- |
| Glyphosate | R1G | 370 (189–1110) | 4.0 | >1350 | >7 |
| Glyphosate | R2G | 347 (164–770) | 3.8 | >1350 | >7 |
| Glyphosate | S | 91 (38–184) | -- | 184 (143–225) | -- |
| Pyroxasulfone | R1P | 0.06 (0.03–0.095) | 15 | 46 (26–66) | 7.7 |
| Pyroxasulfone | R2P | 0.03 (0.015–0.06) | 7.5 | 22 (14–30) | 3.7 |
| Pyroxasulfone | S | 0.004 (0.0004–0.007) | -- | 6.0 (2.1–9.9) | -- |
| Trifluralin | R1T | 35 (6–391) | 29 | >720 | >8 |
| Trifluralin | R2T | 5.4 (0.3–41) | 4.5 | 203 (54–352) | 2.1 |
| Trifluralin | S | 1.2 (0.6–1.8) | -- | 96 (80–112) | -- |
| Agar | |||||
|---|---|---|---|---|---|
| Clethodim | Glyphosate | Pyroxasulfone | Trifluralin | ||
| Clethodim | 0.426 *** | ||||
| Pots | Glyphosate | −0.034 ns | 0.161 ns | ||
| Pyroxasulfone | 0.017 ns | −0.092 ns | −0.098 ns | ||
| Trifluralin | 0.076 ns | −0.023 ns | −0.211 ns | 0.337 ** | |
| Population | Resistance Profile | Year Collected | Population Origin/Code | Reference |
|---|---|---|---|---|
| S | Susceptible | 1985 | VLR1 | Neve and Powles [24] |
| VD4 | Susceptible | 2010 1 | Wongan Hills, WA | Goggin et al. [23] |
| R1C | Clethodim | 2003 | M1/25 | Owen et al. [25] |
| R2C | Clethodim | 2015 | H3/3 | Owen et al. [8] |
| CR | Clethodim | 2018 | WA4-18 | R. Busi [unpublished data] |
| R1G | Glyphosate | 2015 | AFLR2 | Yu et al. [26] |
| R2G | Glyphosate | 2015 | NLR70 | Lorraine-Colwill et al. [27] |
| GR | Glyphosate | 2018 | WA124-18 | R. Busi [unpublished data] |
| R1P | Pyroxasulfone | 2009 1 | MR-P3 2 | Busi et al. [25] |
| R2P | Pyroxasulfone | 2014 1 | MR-P6 2 | Busi and Powles [28] |
| PR | Pyroxasulfone | 2019 1 | M3/54-P4 2 | R. Busi [unpublished data] |
| R1T | Trifluralin | 1982 | SLR31 | Heap and Knight [29] |
| R2T | Trifluralin | 2012 | WA678-13 | P. Boutsalis [unpublished data] |
| TR 3 | Trifluralin | 2018 | WA3-18 | R. Busi [unpublished data] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perez, M.B.; Beckie, H.J.; Cawthray, G.R.; Goggin, D.E.; Busi, R. Rapid On-Farm Testing of Resistance in Lolium rigidum to Key Pre- and Post-Emergence Herbicides. Plants 2021, 10, 1879. https://doi.org/10.3390/plants10091879
Perez MB, Beckie HJ, Cawthray GR, Goggin DE, Busi R. Rapid On-Farm Testing of Resistance in Lolium rigidum to Key Pre- and Post-Emergence Herbicides. Plants. 2021; 10(9):1879. https://doi.org/10.3390/plants10091879
Chicago/Turabian StylePerez, Martina Badano, Hugh J. Beckie, Gregory R. Cawthray, Danica E. Goggin, and Roberto Busi. 2021. "Rapid On-Farm Testing of Resistance in Lolium rigidum to Key Pre- and Post-Emergence Herbicides" Plants 10, no. 9: 1879. https://doi.org/10.3390/plants10091879
APA StylePerez, M. B., Beckie, H. J., Cawthray, G. R., Goggin, D. E., & Busi, R. (2021). Rapid On-Farm Testing of Resistance in Lolium rigidum to Key Pre- and Post-Emergence Herbicides. Plants, 10(9), 1879. https://doi.org/10.3390/plants10091879