
Relationship between β-Carotene Accumulation and Geranylgeranyl Pyrophosphate Synthase in Different Species of Dunaliella



Abstract
:1. Introduction
2. Materials and Methods
2.1. Algae Species and Cultivation
2.2. Observations of Cell Morphology and Determination of Absorbance
2.3. Determination of Photosynthetic Pigment Content and the Chlorophyll Fluorescence Parameters
2.4. Determination of β-Carotene Content
2.5. Determination of the GGPS Activity
2.6. Amplification, Sequencing, and Characterization of GGPS in Dunaliella
3. Results
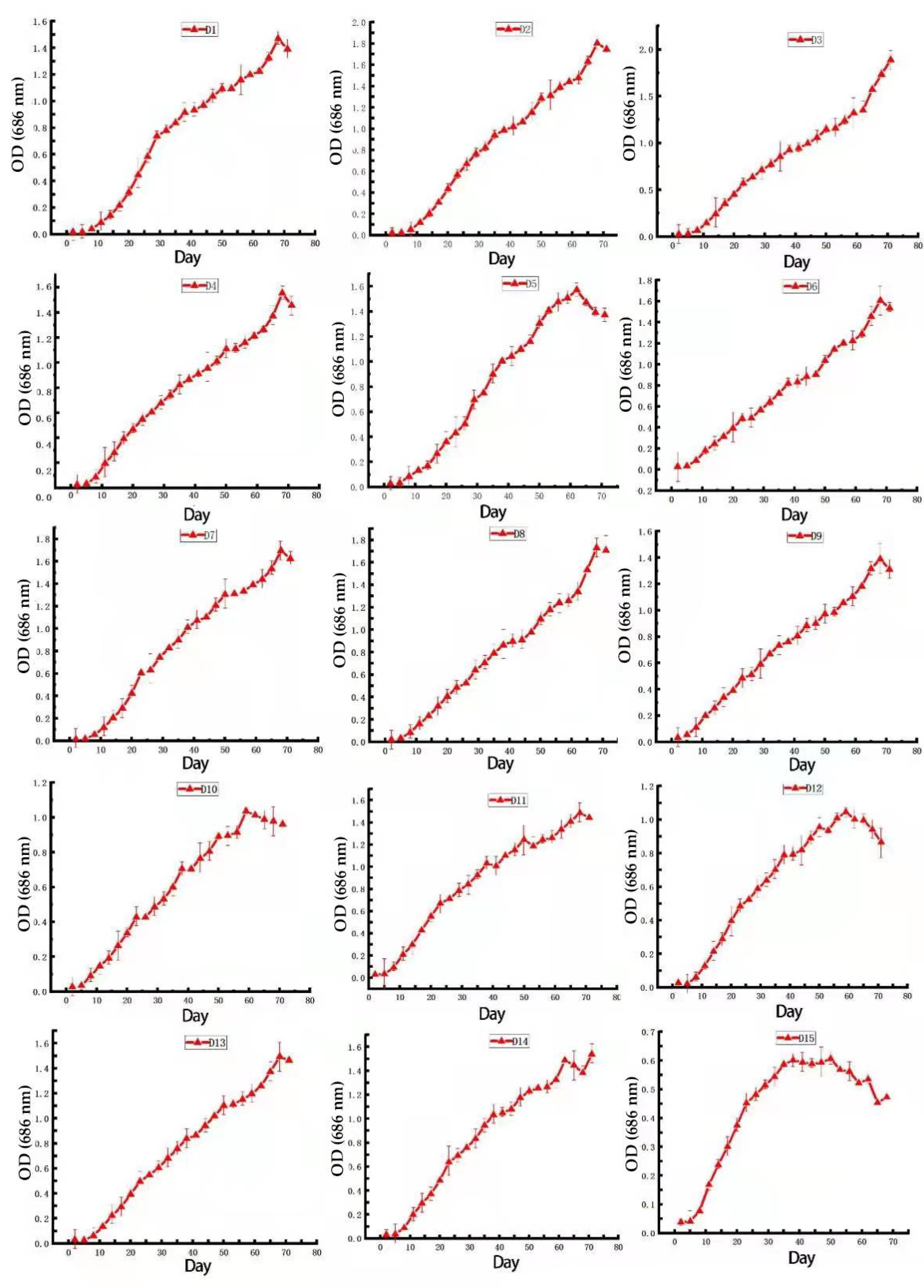
3.1. Dunaliella Growth and Cell Morphology
3.2. Comparison of Photosynthetic Pigment Contents and Chlorophyll Fluorescence Parameters
3.3. Comparison of β-Carotene Content
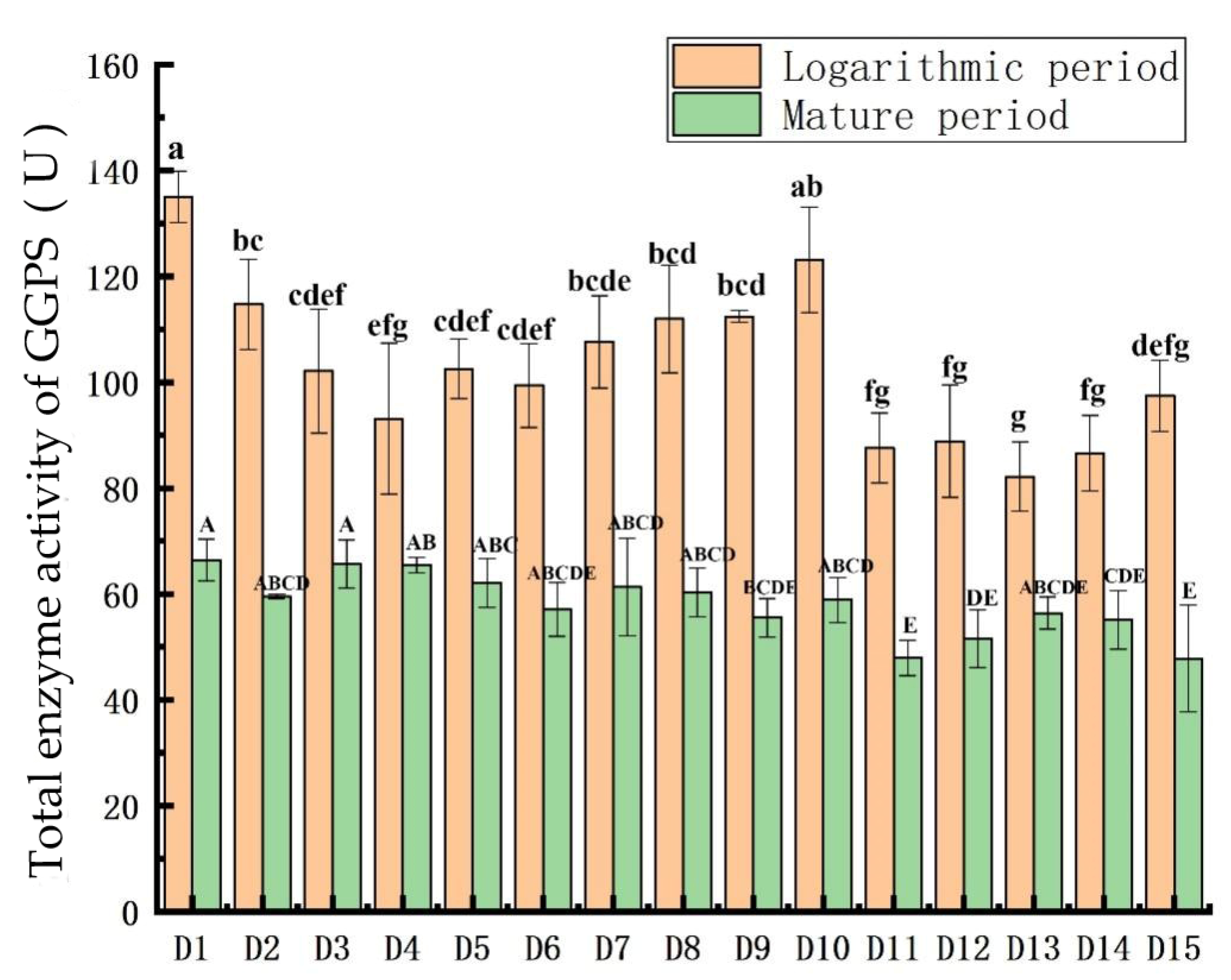
3.4. Comparison of GGPS Activity
3.5. Relationship between β-Carotene Content and GGPS Activity
3.6. Amplification and Characterization of the GGPS Gene
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Phadwal, K.; Singh, P.K. Isolation and characterization of an indigenous isolate of Dunaliella sp. for beta-carotene and glycerol production from a hypersaline lake in India. J. Basic Microbiol. 2003, 43, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Sadka, A.; Lers, A.; Zamir, A. A critical examination of the role of de novo protein synthesis in the osmotic adaptation of the halotolerant alga Dunaliella. FEBS Lett. 1989, 244, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Borowitzka, M.A. High-value products from microalgae their development and commercialisation. J. Appl. Phycol. 2013, 25, 743–756. [Google Scholar] [CrossRef]
- Benamotz, A.; Avron, M. The biotechnology of cultivating the halotolerant alga Dunaliella. Trends Biotechnol. 1990, 8, 121–126. [Google Scholar] [CrossRef]
- Arun, N.; Singh, D.P. A review on pharmacological applications of halophilic alga Dunaliella. Indian J. Geo-Mar. Sci. 2016, 45, 440–447. [Google Scholar]
- Ben-Amotz, A. New Mode of Dunaliella Biotechnology: Two-phase Growth for Beta-Carotene Production. J. Appl. Phycol. 1995, 7, 65–68. [Google Scholar] [CrossRef]
- Vanitha, A.; Narayan, M.S.; Murthy, K.N.C. Comparative study of lipid composition of two halotolerant alga, Dunaliella bardawil and Dunaliella salina. Int. J. Food Sci. Nutr. 2007, 58, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Del Campo, J.A.; García-González, M.; Guerrero, M.G. Outdoor cultivation of microalgae for carotenoid production: Current state and perspectives. Appl. Microbiol. Biotechnol. 2007, 74, 1163–1174. [Google Scholar] [CrossRef]
- Hosseini, T.A.; Shariati, M. Dunaliella biotechnology: Methods and applications. J. Appl. Microbiol. 2009, 107, 14–35. [Google Scholar] [CrossRef] [PubMed]
- Sui, Y.; Vlaeminck, S.E. Dunaliella microalgae for nutritional protein: An undervalued asset. Trends Biotechnol. 2020, 38, 10–12. [Google Scholar] [CrossRef]
- Ye, Z.W.; Jiang, J.G.; Wu, G.H. Biosynthesis and regulation of carotenoids in Dunaliella: Progresses and prospects. Biotechnol. Adv. 2008, 26, 352–360. [Google Scholar] [CrossRef]
- Shewmaker, C.K.; Sheehy, J.A.; Daley, M. Seed-specific overexpression of phytoene synthase: Increase in carotenoids and other metabolic effects. Plant J. 1999, 20, 401–412. [Google Scholar] [CrossRef]
- Schmidt-Dannert, C. Engineering novel carotenoids in microorganisms. Curr. Opin. Biotechnol. 2000, 11, 255–261. [Google Scholar] [CrossRef]
- Albrecht, V.; Klein, A.; Hugueney, P. Molecular cloning and functional expression in E. coli of a novel plant enzyme mediating ζ-carotene desaturation. FEBS Lett. 1995, 372, 199–202. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Feng, J.; Xie, S.L. Effect of culture conditions on the content of β-carotene in Dunaliella. Sci. Technol. Food Ind. 2014, 35, 177–181. [Google Scholar]
- Chisti, Y. Biodiesel from microalgae beats bioethanol. Trends Biotechnol. 2008, 26, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Mera, R.; Torres, E.; Abalde, J. Effect of sodium sulfate on the freshwater microalga Chlamydomonas moewusii: Implications for the optimization of algal culture media. J. Phycol. 2016, 52, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Markou, G.; Muylaert, K. Effect of light intensity on the degree of ammonia toxicity on PSII activity of Arthrospira platensis and Chlorella vulgaris. Bioresour. Technol. 2016, 216, 453–461. [Google Scholar] [CrossRef]
- El-Baz, F.K.; Abdel Jaleel, G.A.; Saleh, D.O.; Hussein, R.A. Protective and therapeutic potentials of Dunaliella salina on aging-associated cardiac dysfunction in rats. Asian Pac. J. Trop. Biomed. 2018, 8, 403–410. [Google Scholar]
- Lu, B.J.; Zhai, Y.X.B. Application of Thin Layer Chromatography in Food Analysis, 1st ed.; Peking University Press: Beijing, China, 1991; pp. 135–138. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Lemberkovics, E.; Kéry, A.; Simándi, B. Influence of extraction methods on the composition of essential oils. Acta Pharm Hung. 2004, 74, 166–170. [Google Scholar] [PubMed]
- Higgins, D.G.; Sharp, P.M. Clustal: A package for performing multiple sequence alignment on a microcomputer. Elsevier 1988, 73, 237–244. [Google Scholar] [CrossRef]
- Telles, G.P.; Araújo, G.S.; Walter, M.E.M.T. Live neighbor-joining. BMC Bioinf. 2018, 19, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Jiang, J.G. Osmotic responses of Dunaliella to the changes of salinity. J. Cell Physiol. 2009, 219, 251–2588. [Google Scholar] [CrossRef] [PubMed]
- Polle, J.E.W.; Struwe, L.; Jin, E. Identification and characterization of a new strain of the unicellular green alga Dunaliella salina (Teod.) from Korea. J. Microbiol. Biotchnol. 2008, 18, 821–827. [Google Scholar]
- Borowitzka, M.A.; Siva, C.J. The taxonomy of the genus Dunaliella (Chlorophyta, Dunaliella) with emphasis on the marine and halophilic species. J. Appl. Phycol. 2007, 19, 567–590. [Google Scholar] [CrossRef]
- Rizza, F.; Pagani, D.; Stanca, A.M. Use of chlorophyll fluorescence to evaluate the cold acclimation and freezing tolerance of winter and spring oats. Plant Breed. 2001, 120, 389–396. [Google Scholar] [CrossRef]
- Havaux, M.; Strasser, R.J.; Greppin, H. A theoretical and experimental analysis of the qP and qN coefficients of chlorophyll fluorescence quenching and their relation to photochemical and nonphotochemical events. Photosynth Res. 1991, 27, 41–55. [Google Scholar] [CrossRef] [PubMed]
- Oukarroum, A. Change in Photosystem II Photochemistry During Algal Growth Phases of Chlorella vulgaris and Scenedesmus obliquus. Curr. Microbiol. 2016, 72, 692. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.F.; Lou, S.L.; You, L.Y. Extraction and content comparison of β-carotene from ten species (Species) of Dunaliella. J. Xiamen Univ. Nat. Sci. 1995, 34, 94–98. [Google Scholar]
- Sun, J.; Sun, X.X.; Tang, P.W. Molecular cloning and functional expression of two key carotene synthetic genes derived from Blakeslea trispora into E. coli for increased β-carotene production. Biotechnol. Lett. 2012, 34, 2077–2082. [Google Scholar] [CrossRef] [PubMed]
- Preetha, K.; John, L.; Subin, C.S.; Vijayan, K.K. Phenotypic and genetic characterization of Dunaliella (Chlorophyta) from Indian salinas and their diversity. Aquat Biosyst. 2012, 8, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Strain | Source |
|---|---|---|
| D1 | D. primolecta | Culture Collection of Algae and Protozoa (CCAP 11/34) |
| D2 | D. maritima | Culture Collection of Algae and Protozoa (CCAP 19/1) |
| D3 | D. peircei | Culture Collection of Algae and Protozoa (CCAP 19/2) |
| D4 | D. viridis | Culture Collection of Algae and Protozoa (CCAP 19/3) |
| D5 | D. bioculata | Culture Collection of Algae and Protozoa (CCAP 19/4) |
| D6 | D. quartolecta | Culture Collection of Algae and Protozoa (CCAP 19/8) |
| D7 | D. parva | Culture Collection of Algae and Protozoa (CCAP 19/10) |
| D8 | D. salina | Culture Collection of Algae and Protozoa (CCAP 19/12) |
| D9 | D. polymorpha | Culture Collection of Algae and Protozoa (CCAP 19/14) |
| D10 | D. tertiolecta | Culture Collection of Algae and Protozoa (CCAP 19/22) |
| D11 | D. acidophila | Culture Collection of Algae and Protozoa (CCAP 19/35) |
| D12 | D. bioculata_C33 | Culture Collection of Algae at the University of Texas at Austin (LB199) |
| D13 | D. salina | Salt Lake in Yuncheng City, Shanxi Province |
| D14 | Dunaliella sp. | Institute of Hydrobiology, Chinese Academy of Sciences |
| D15 | D. apiculata | National Center for Marine Algae and Microbiota (UW 481) |
| Formulate Content | Components | Dosage |
|---|---|---|
| Dunaliella medium | NaCl | 87.69 g |
| NaNO3 | 0.42 g | |
| NaH2PO4·2H2O | 0.0156 g | |
| CaCl2·2H2O | 0.044 g | |
| KCl | 0.074 g | |
| MgSO4·7H2O | 1.23 g | |
| NaHCO3 | 0.84 g | |
| FeC6H5O7 | 0.002 g | |
| UPW | 1000 mL | |
| A5 | H3BO3 | 286 mg |
| MnCl2·4H2O | 181 mg | |
| ZnSO4·7H2O | 22.2 mg | |
| Na2MoO4·2H2O | 39 mg | |
| CuSO4·5H2O | 7.9 mg | |
| Co (NO3)2·6H2O | 4.9 mg | |
| UPW | 100 mL |
| Primer | Forward Primer | Reverse Primer | Tm′ |
|---|---|---|---|
| A | 5′-AAGAGCGAAAACGAGGAGGT-3′ | 5′-ATTCAGGGCGCTTCAAGGTT-3′ | 56 °C |
| B | 5′-GGTGCACACCATGAGCCT-3′ | 5′-TGGCAGGGCACTGCTCTTA-3′ | 56 °C |
| C | 5′-AGGGGGTTGTAGACCGTGTT-3′ | 5′-CTCCACTGTCACGTCGTCCG-3′ | 54 °C |
| Species | No. | Height (H) | Width (W) | Shape | Anova (H/W) |
|---|---|---|---|---|---|
| D. primolecta | D1 | 7.7 ± 0.8 μm | 4 ± 0.2 μm | Oval | A |
| D. maritima | D2 | 8.3 ± 1.2 μm | 6.3 ± 0.9 μm | Pear | AB |
| D. peircei | D3 | 7.3 ± 0.6 μm | 7.3 ± 0.5 μm | Oval | AB |
| D. viridis | D4 | 9 ± 1.3 μm | 7 ± 1 μm | Oval | AB |
| D. bioculata | D5 | 8.7 ± 1.1 μm | 4.7 ± 0.3 μm | Pear | AB |
| D. quartolecta | D6 | 9 ± 1.3 μm | 7 ± 1.2 μm | Oval | AB |
| D. parva | D7 | 9.3 ± 1.2 μm | 6.3 ± 1.1 μm | Oval | AB |
| D. salina | D8 | 10.3 ± 2 μm | 10.3 ± 1.9 μm | Round | B |
| D. polymorpha | D9 | 6.9 ± 0.5 μm | 4.6 ± 0.2 μm | Round | AB |
| D. tertiolecta | D10 | 7.3 ± 0.8 μm | 6.7 ± 0.8 μm | Round | AB |
| D. acidophila | D11 | 9.3 ± 1.1 μm | 6.9 ± 0.5 μm | Oval | AB |
| D. bioculata_C33 | D12 | 8.7 ± 1 μm | 6.7 ± 0.8 μm | Oval | AB |
| D. salina_42 | D13 | 9.2 ± 1.5 μm | 7.2 ± 1.2 μm | Oval | AB |
| Dunaliella sp. | D14 | 8.7 ± 1 μm | 6.8 ± 1 μm | Oval | AB |
| D. apiculata | D15 | 9.7 ± 1.4 μm | 6.7 ± 0.7 μm | Pear | A |
| D8 | D10 | D7 | D5 | D2 | D1 | D9 | D11 | D13 | D15 | D14 | D3 | D6 | D12 | D4 | Chlamydomonas | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| D8 | 0 | 0 | 0 | 0 | 3 | 2 | 0 | 0 | 0 | 1 | 3 | 7 | 14 | 29 | 635 | |
| D10 | 0.00 | 0 | 0 | 0 | 3 | 2 | 0 | 0 | 0 | 1 | 3 | 7 | 14 | 29 | 635 | |
| D7 | 0.00 | 0.00 | 0 | 0 | 3 | 2 | 0 | 0 | 0 | 1 | 3 | 7 | 14 | 29 | 635 | |
| D5 | 0.00 | 0.00 | 0.00 | 0 | 3 | 2 | 0 | 0 | 0 | 1 | 3 | 7 | 14 | 29 | 635 | |
| D2 | 0.00 | 0.00 | 0.00 | 0.00 | 3 | 2 | 0 | 0 | 0 | 1 | 3 | 7 | 14 | 29 | 635 | |
| D1 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 4 | 3 | 3 | 3 | 4 | 6 | 10 | 17 | 32 | 638 | |
| D9 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 2 | 2 | 2 | 3 | 5 | 9 | 16 | 31 | 637 | |
| D11 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0 | 0 | 1 | 3 | 7 | 14 | 29 | 635 | |
| D13 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0 | 1 | 3 | 7 | 14 | 29 | 635 | |
| D15 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1 | 3 | 7 | 14 | 29 | 635 | |
| D14 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 4 | 8 | 15 | 30 | 635 | |
| D3 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 10 | 17 | 30 | 637 | |
| D6 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.01 | 0.01 | 0.00 | 0.00 | 0.00 | 0.00 | 0.01 | 21 | 32 | 639 | |
| D12 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | 43 | 637 | |
| D4 | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 | 0.03 | 640 | |
| Chlamydomonas | 0.39 | 0.39 | 0.39 | 0.39 | 0.39 | 0.39 | 0.39 | 0.39 | 0.39 | 0.39 | 0.39 | 0.39 | 0.39 | 0.39 | 0.39 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, L.; Gao, F.; Feng, J.; Lv, J.; Liu, Q.; Nan, F.; Liu, X.; Xie, S. Relationship between β-Carotene Accumulation and Geranylgeranyl Pyrophosphate Synthase in Different Species of Dunaliella. Plants 2022, 11, 27. https://doi.org/10.3390/plants11010027
Xu L, Gao F, Feng J, Lv J, Liu Q, Nan F, Liu X, Xie S. Relationship between β-Carotene Accumulation and Geranylgeranyl Pyrophosphate Synthase in Different Species of Dunaliella. Plants. 2022; 11(1):27. https://doi.org/10.3390/plants11010027
Chicago/Turabian StyleXu, Lu, Fan Gao, Jia Feng, Junping Lv, Qi Liu, Fangru Nan, Xudong Liu, and Shulian Xie. 2022. "Relationship between β-Carotene Accumulation and Geranylgeranyl Pyrophosphate Synthase in Different Species of Dunaliella" Plants 11, no. 1: 27. https://doi.org/10.3390/plants11010027
APA StyleXu, L., Gao, F., Feng, J., Lv, J., Liu, Q., Nan, F., Liu, X., & Xie, S. (2022). Relationship between β-Carotene Accumulation and Geranylgeranyl Pyrophosphate Synthase in Different Species of Dunaliella. Plants, 11(1), 27. https://doi.org/10.3390/plants11010027