
Mesoamerican Cypripedium: Mycorrhizal Contributions to Promote Their Conservation as Critically Endangered Species


Abstract
:1. Introduction
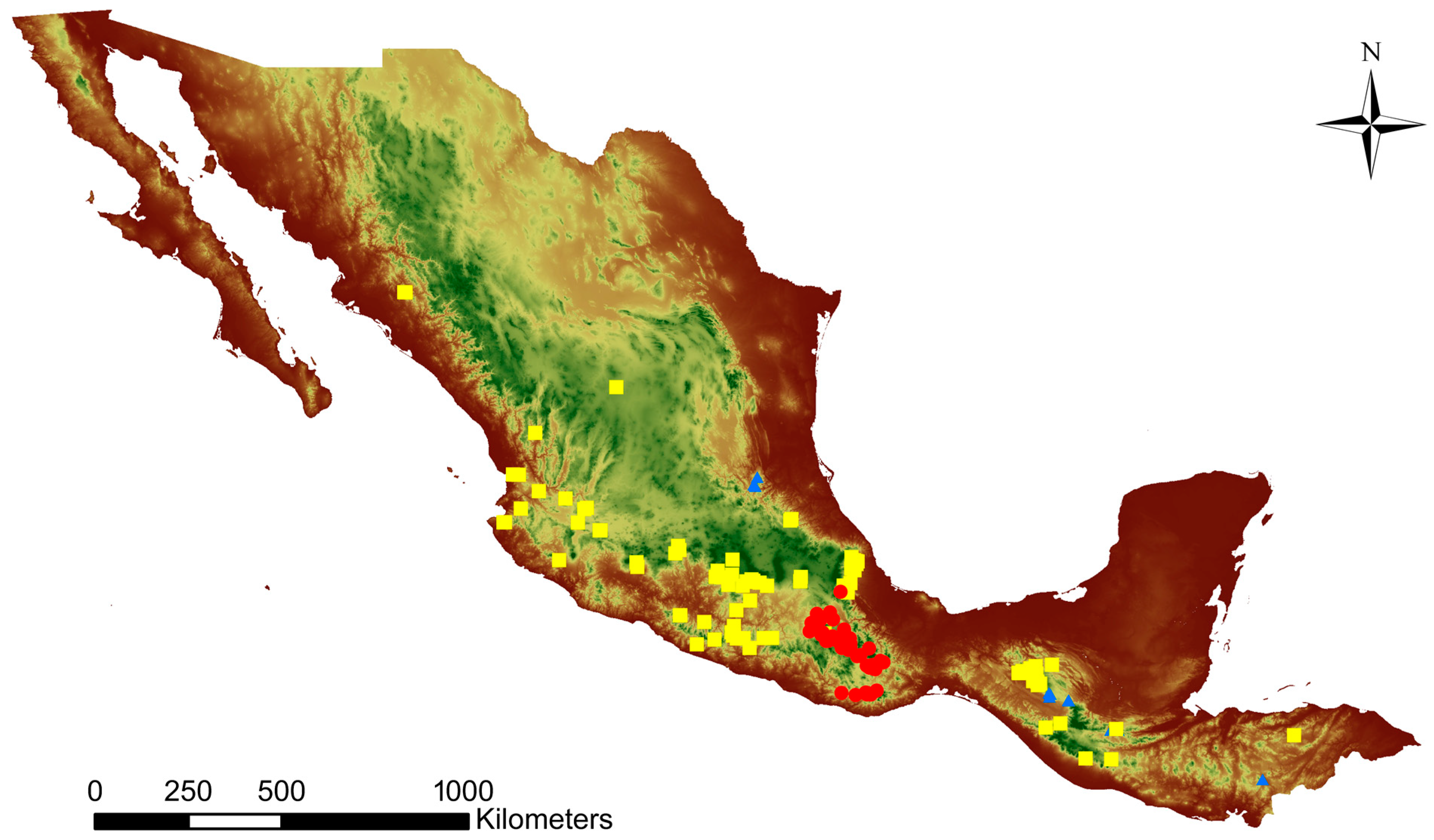
2. Mesoamerican Slipper Orchids: Unique and Critically Endangered
3. Underground Growth Pattern in Different Habitats
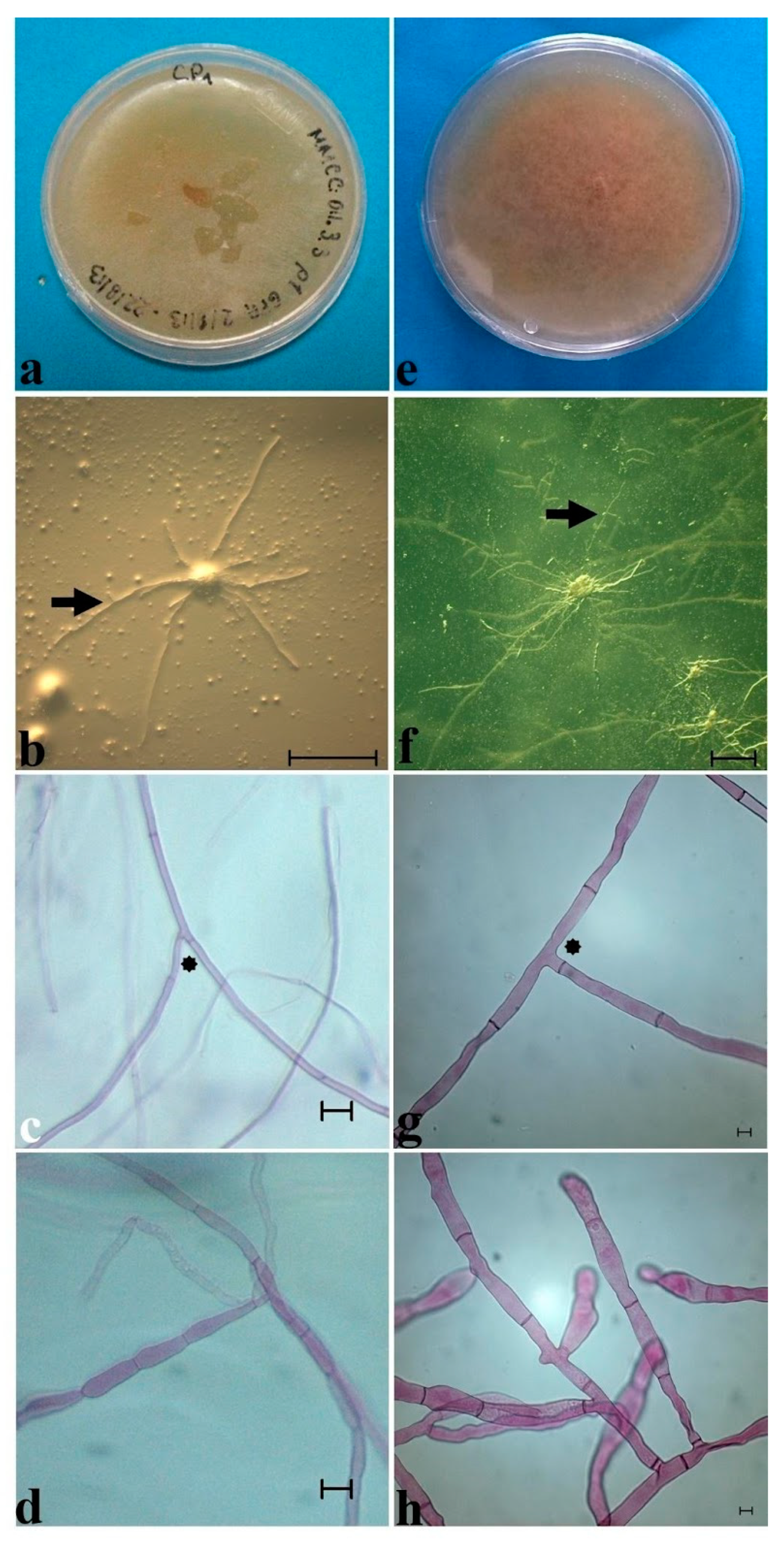
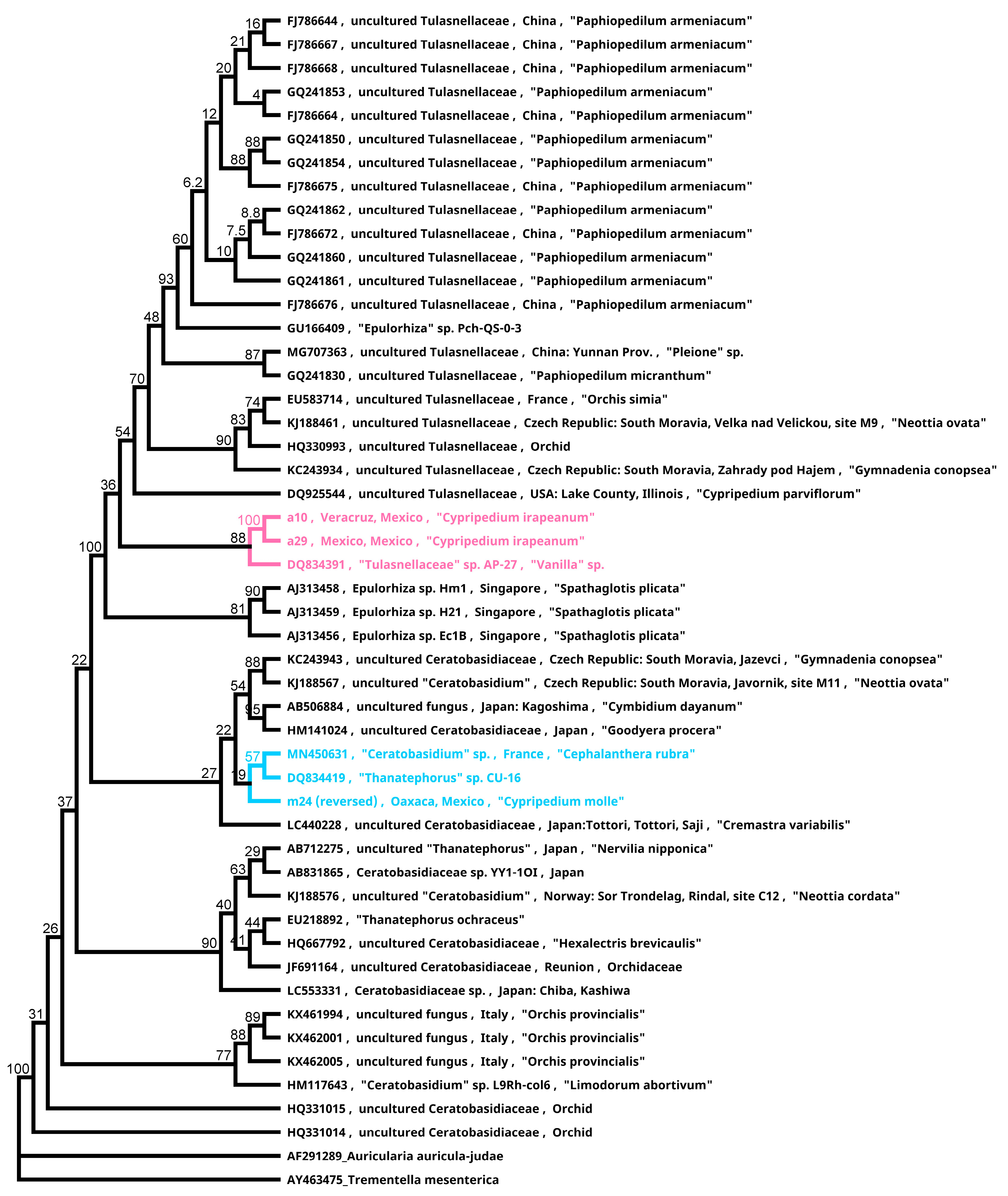
4. Mycorrhiza Studies for Conservation Purposes
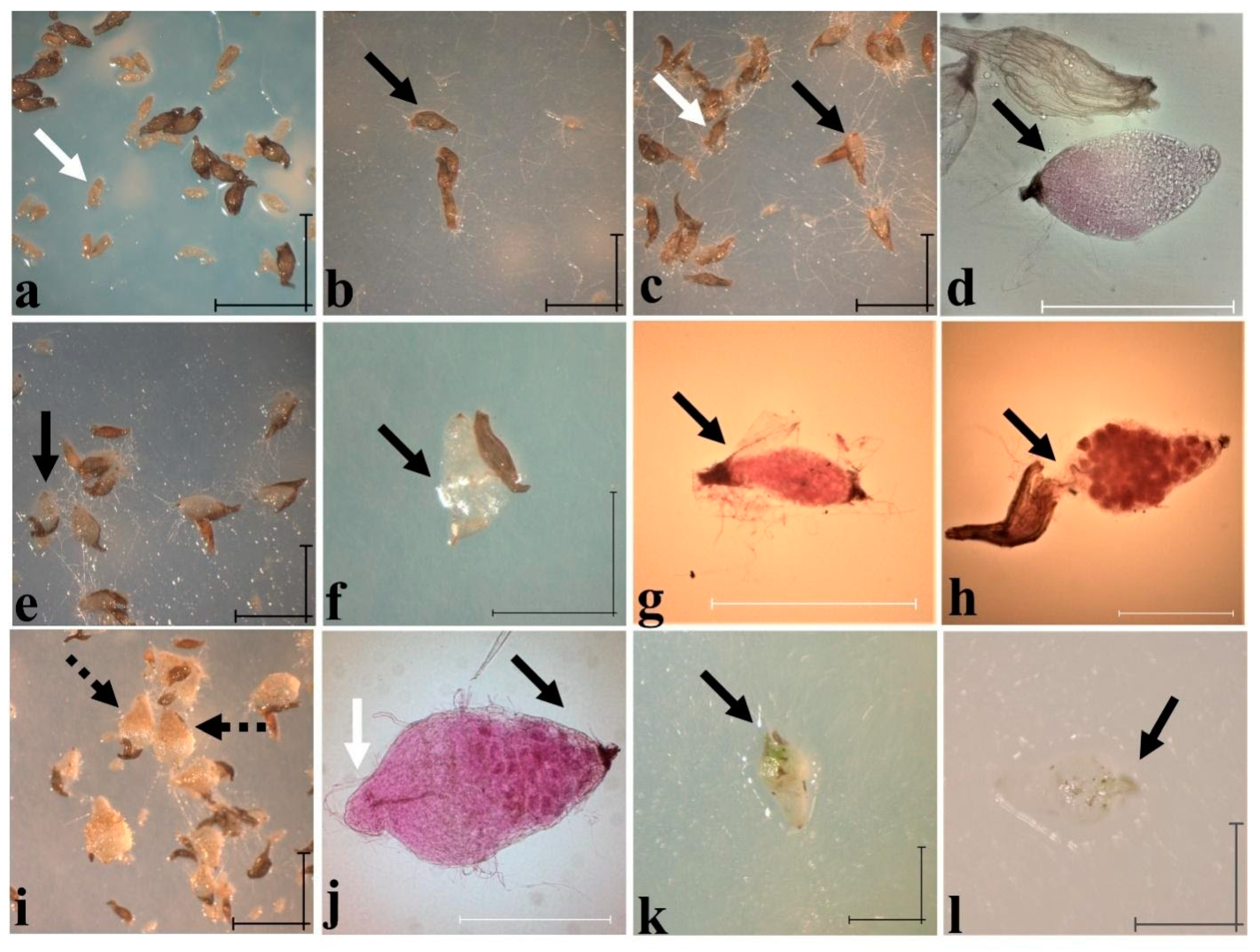
5. Symbiotic and Asymbiotic Germination and Development
6. Final Remarks
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chase, M.W. Classification of Orchidaceae in the Age of DNA Data. Curtis’s Bot. Mag. 2005, 22, 2–7. [Google Scholar] [CrossRef]
- Guo, Y.Y.; Luo, Y.B.; Liu, Z.J.; Wang, X.Q. Evolution and biogeography of the slipper orchids: Eocene vicariance of the conduplicate genera in the old and New World tropics. PLoS ONE 2012, 7, e38788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bunch, W.D.; Cowden, C.C.; Wurzburger, N.; Shefferson, R.P. Geography and soil chemistry drive the distribution of fungal associations in lady’s slipper orchid, Cypripedium acaule. Botany 2013, 91, 850–856. [Google Scholar] [CrossRef]
- Cribb, P.J. The Genus Cypripedium, 1st ed.; Green, P., Ed.; Timber Press: Portland, OR, USA, 1997; ISBN 9780881924039. [Google Scholar]
- Gargiulo, R.; Pironon, S.; Zheleznaya, E.; Sanchez, M.D.; Balázs, Z.R.; Podar, D.; Wilkinson, T.; Jäkäläniemi, A.; Kull, T.; Väre, H.; et al. Phylogeography and post-glacial dynamics in the clonal-sexual orchid Cypripedium calceolus L. J. Biogeogr. 2019, 46, 526–538. [Google Scholar] [CrossRef]
- Kellof, C.L.; Kass, L.B. Databasing and Georeferencing Historical Collections to Discover Potential Sites for Rare and Endangered Plants of New York, U.S.A. J. Bot. Res. Inst. Tex. J. Bot. Res. 2018, 12, 323–368. [Google Scholar] [CrossRef]
- Li, J.; Liu, Z.; Salazar, G.A.; Bernhardt, P.; Perner, H.; Tomohisa, Y.; Jin, X.; Chung, S.; Luo, Y. Molecular phylogeny of Cypripedium (Orchidaceae: Cypripedioideae) inferred from multiple nuclear and chloroplast regions. Mol. Phylogenet. Evol. 2011, 61, 308–320. [Google Scholar] [CrossRef]
- Pérez-García, E.; Mó, E. Die Cypripedioideae von Mesoamerika. Teil 1a Cypripedium. Die Orchidee. 2014, 65, 476–482. [Google Scholar]
- Unruh, S.A.; McKain, M.R.; Lee, Y.I.; Yukawa, T.; McCormick, M.K.; Shefferson, R.P.; Smithson, A.; Leebens-Mack, J.H.; Pires, J.C. Phylotranscriptomic Analysis and Genome Evolution of the Cypripedioideae (Orchidaceae). Am. J. Bot. 2018, 105, 631–640. [Google Scholar] [CrossRef] [Green Version]
- CITES Appendices | CITES. Available online: https://cites.org/eng/app/appendices.php (accessed on 19 August 2020).
- Mcgough, H.N.; Roberts, D.L.; Brodie, C. CITES y Las Zapatillas de Venus. Royal Botanic Gardens, Kew, United Kingdom. 2006. Available online: https://www.yumpu.com/es/document/read/14398272/cites-y-las-zapatillas-de-venus-royal-botanic-gardens-kew (accessed on 19 August 2020).
- Hagsater, E.; Soto Arenas, M.A.; Salazar, G.; Machorro, J.; López Rosas, M.; Dressler, R. Orchids of Mexico, 1st ed.; Productos Farmacéuticos, S.A. de C.V.; Instituto Chinoin & Redacta: Ciudad de México, México, 2005; ISBN 968788908X. [Google Scholar]
- Soto-Arenas, M.A.; Solano-Gómez, R.A. Ficha Técnica de Cypripedium dickinsonianum. In Información Actualizada Sobre las Especies de Orquídeas del PROY-NOM-059-ECOL-2000; Soto-Arenas, M.A., Solano Gómez, R., Eds.; SNIB-CONABIO: Ciudad de México, Mexico, 2007; pp. 1–7. [Google Scholar]
- Soto-Arenas, M.A.; Solano-Gómez, R.A. Ficha Técnica de Cypripedium irapeanum. In Información Actualizada Sobre las Especies de Orquídeas del PROY-NOM-059-ECOL-2000; Soto-Arenas, M.A., Solano Gómez, R., Eds.; SNIB-CONABIO: Ciudad de México, Mexico, 2007; pp. 1–12. [Google Scholar]
- Soto Arenas, M.Á.; Gómez, R.S.; Hágsater, E. Risk of Extinction and Patterns of Diversity Loss in Mexican Orchids. Lankesteriana 2007, 7, 114–121. [Google Scholar] [CrossRef]
- González-Tamayo, J.R.; Hernández, L. Las Orquídeas del Occidente de México; González-Tamayo, J.R., Hernández, H.L., Eds.; COECYTJAL: Guadalajara, México, 2010; Volume 1, ISBN 9786070040412. [Google Scholar]
- McVaugh, R. Flora Novo-Galiciana: A Descriptive Account of the Vascular Plants of Western Mexico: Orchidaceae, 1st ed.; Anderson, W.R., Ed.; University of Michigan Press: Ann Arbor, MI, USA, 1985; Volume 16, ISBN 0472048163. [Google Scholar]
- Szlachetko, D.L.; Kolanowska, M.; Muller, F.; Vannini, J.; Rojek, J.; Górniak, M. First Guatemalan Record of Natural Hybridisation between Neotropical Species of the Lady’s Slipper Orchid (Orchidaceae, Cypripedioideae). PeerJ 2017, 5, e4162. [Google Scholar] [CrossRef]
- Pérez-García, E.; Mó-Mó, E. Die Cypripedioideae von Mesoamerika. Teil 2b Phragmipedium. Die Orchidee. 2015, 66, 216–225. [Google Scholar]
- Rankou, H.; Salazar, G.A. Conservation Status of Cypripedium irapeanum. Available online: https://www.iucnredlist.org/species/43316630/43327669 (accessed on 19 August 2020).
- NOM-059-SEMARNAT-2010, N.O.M.; NORMA Oficial Mexicana NOM-059-SEMARNAT-2010. Diario Oficial de la Federación: México, 2019. Available online: https://www.dof.gob.mx/normasOficiales/4254/semarnat/semarnat.htm (accessed on 1 April 2022).
- California Academy of Sciences, 2008. Una Comunidad Para Naturalistas. INaturalist. Available online: https://www.inaturalist.org/ (accessed on 12 March 2020).
- INEGI Instituto Nacional de Estadística y Geografía. Available online: http://www.inegi.org.mx/ (accessed on 4 November 2018).
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum Entropy Modeling of Species Geographic Distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Proal, J.P. Las Secuelas del Boom Mezcalero: Sabor Adulterado, Daños al Ecosistema, Corrupción. Available online: https://www.proceso.com.mx/428051/las-secuelas-del-boom-mezcalero-sabor-adulterado-danos-al-ecosistema-corrupcion?fbclid=IwAR3ENT16jaT081WKYWFcgjQ0EsSRCUH6xgvvp116vxgr2_HrErewDMFzrGk (accessed on 26 May 2022).
- Fick, S.E.; Hijmans, R.J. Worldclim 2: New 1-Km Spatial Resolution Climate Surfaces for Global Land Areas. Available online: http://worldclim.org/version2 (accessed on 26 May 2022).
- Phillips, S.J.; Dudík, M.; Schapire, R.E. Maxent Software for Modeling Species Niches and Distributions; (open-source software) (Version 3.4.1); Available online: http://biodiversityinformatics.amnh.org/open_source/maxent/ (accessed on 14 August 2018).
- Global Soil Data Task Group Global Gridded Surfaces of Selected Soil Characteristics (IGBP-DIS). Available online: https://daac.ornl.gov/cgi-bin/dsviewer.pl?ds_id=569 (accessed on 26 May 2022).
- ESRI. Esri: ArcGIS© Pro (Version 2.5), 2016. Available online: https://support.esri.com/en/products/desktop/arcgis-desktop/arcgis-pro/2-5-1 (accessed on 1 April 2022).
- Liu, H.; Jacquemyn, H.; He, X.; Chen, W.; Huang, Y.; Yu, S.; Lu, Y.; Zhang, Y. The Impact of Human Pressure and Climate Change on the Habitat Availability and Protection of Cypripedium (Orchidaceae) in Northeast China. Plants 2021, 10, 84. [Google Scholar] [CrossRef] [PubMed]
- Curtis, J.T. Germination and Seedling Development in Five Species of Cypripedium L. Am. J. Bot. 1943, 30, 199–206. [Google Scholar] [CrossRef]
- Rasmussen, H.N. Terrestrial Orchids, 1st ed.; Danish Institute of Plant and Soil Science, Ed.; Cambridge University Press: New York, NY, USA, 1995; ISBN 9780521451659. [Google Scholar]
- Weldy, T.W.; Mlodozeniec, H.T.; Wallace, L.E.; Case, M.A. The Current Status of Cypripedium kentuckiense (Orchidaceae) Including a Morphological Analysis of a Newly Discovered Population in Eastern Virginia. SIDA Contrib. Bot. 1996, 17, 423–435. [Google Scholar]
- Zhongjian, L.; Lijun, C.; Wenhui, R.; Liqiang, L.; Yuting, Z. Correlation between Numerical Dynamics and Reproductive Behavior in Cypripedium lentiginosum. Acta Ecol. Sin. 2008, 28, 111–121. [Google Scholar] [CrossRef]
- Moreno Camarena, M. Caracterización Molecular de Endofitos Micorrízicos y Germinación Simbiótica de Cypripedium irapeanum (Orchidaceae); Universidad Nacional Autónoma de México: Ciudad de México, Mexico, 2016. [Google Scholar]
- Shefferson, R.P.; Weiß, M.; Kull, T.; Taylor, D.L. High Specificity Generally Characterizes Mycorrhizal Association in Rare Lady’s Slipper Orchids, Genus Cypripedium. Mol. Ecol. 2005, 14, 613–626. [Google Scholar] [CrossRef]
- Shefferson, R.P.; Taylor, D.L.; Weiß, M.; Garnica, S.; McCormick, M.K.; Adams, S.; Gray, H.M.; McFarland, J.W.; Kull, T.; Tali, K.; et al. The Evolutionary History of Mycorrhizal Specificity among Lady’s Slipper Orchids. Evolution 2007, 61, 1380–1390. [Google Scholar] [CrossRef]
- Shefferson, R.P.; Kull, T.; Tali, K.; Kellett, K.M. Linking Vegetative Dormancy to Fitness in Two Long-Lived Herbaceous Perennials. Ecosphere 2012, 3, art13. [Google Scholar] [CrossRef]
- Shefferson, R.P.; Kull, T.; Hutchings, M.J.; Selosse, M.A.; Jacquemyn, H.; Kellett, K.M.; Menges, E.S.; Primack, R.B.; Tuomi, J.; Alahuhta, K.; et al. Drivers of Vegetative Dormancy across Herbaceous Perennial Plant Species. Ecol. Lett. 2018, 21, 724–733. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, H.N.; Dixon, K.W.; Jersáková, J.; Těšitelová, T. Germination and Seedling Establishment in Orchids: A Complex of Requirements. Ann. Bot. 2015, 116, 391–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez-Apolinar, M.; Gutierrez-Paredes, C.C.; Sanchez-Gallen, I.; Aguirre, E.; Perez García, E.A. Ecological Aspects of Cypripedium irapeanum La Llave & Lex., an Endangered Mexican orchid Species. Slipper Orchid Alliance Newsl. 2012, 13, 1–5. [Google Scholar]
- Rasmussen, H.N.; Pedersen, H.Æ. Cypripedium calceolus Germination in situ: Seed Longevity, and Dormancy Breakage by Long Incubation and Cold Winters. Eur. J. Enviromental Sci. 2012, 1, 69–70. [Google Scholar] [CrossRef] [Green Version]
- Shimura, H.; Koda, Y. Enhanced Symbiotic Seed Germination of Cypripedium macranthos Var. rebunense Following Inoculation after Cold Treatment. Physiol. Plant. 2005, 123, 281–287. [Google Scholar] [CrossRef]
- Yuan, L.; Yang, Z.L.; Li, S.Y.; Hu, H.; Huang, J.L. Mycorrhizal Specificity, Preference, and Plasticity of Six Slipper Orchids from South Western China. Mycorrhiza 2010, 20, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Antonelli, A.; Dahlberg, C.J.; Carlgren, K.H.I.; Appelqvist, T. Pollination of the Lady’s Slipper Orchid (Cypripedium calceolus) in Scandinavia—Taxonomic and Conservational Aspects. Nord. J. Bot. 2009, 27, 266–273. [Google Scholar] [CrossRef]
- Sugiura, N.; Fujie, T.; Inoue, K.; Kitamura, K. Flowering Phenology, Pollination, and Fruit Set of Cypripedium macranthos var. rebunense, a Threatened Lady’s Slipper (Orchidaceae). J. Plant Res. 2001, 114, 171–178. [Google Scholar] [CrossRef]
- Suetsugu, K.; Fukushima, S. Pollination Biology of the Endangered Orchid Cypripedium japonicum in a Fragmented Forest of Japan. Plant Species Biol. 2014, 29, 294–299. [Google Scholar] [CrossRef]
- Shefferson, R.P.; Bunch, W.; Cowden, C.C.; Lee, Y.I.; Kartzinel, T.R.; Yukawa, T.; Downing, J.; Jiang, H. Does Evolutionary History Determine Specificity in Broad Ecological Interactions? J. Ecol. 2019, 107, 1582–1593. [Google Scholar] [CrossRef]
- Colin Rivera, J.; Ortega-Larrocea, M.P. Colonización Micorrizica de Cypripedium irapeanum La Llave & Lex. (Orchidaceae) En Un Bosque de Quercus. In Conservación de orquídeas en México; Téllez-Velasco, M.d.l.A., Ed.; Instituto de Biología, UNAM: Ciudad de México, México, 2012; pp. 186–192. [Google Scholar]
- Porras-Alfaro, A.; Bayman, P. Mycorrhizal Fungi of Vanilla: Diversity, Specificity and Effects on Seed Germination and Plant Growth. Mycologia 2007, 99, 510–525. [Google Scholar] [CrossRef]
- Shimura, H.; Sadamoto, M.; Matsuura, M.; Kawahara, T.; Naito, S.; Koda, Y. Characterization of Mycorrhizal Fungi Isolated from the Threatened Cypripedium macranthos in a Northern Island of Japan: Two Phylogenetically Distinct Fungi Associated with the Orchid. Mycorrhiza 2009, 19, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Clements, M.A.; Muir, H.; Cribb, P.J. A Preliminary Report on the Symbiotic Germination of European Terrestrial Orchids. Kew Bull. 1986, 41, 437. [Google Scholar] [CrossRef]
- Taylor, D.L.; McCormick, M.K. Internal Transcribed Spacer Primers and Sequences for Improved Characterization of Basidiomycetous Orchid Mycorrhizas. New Phytol. 2008, 177, 1020–1033. [Google Scholar] [CrossRef] [PubMed]
- Egidi, E.; May, T.W.; Franks, A.E. Seeking the Needle in the Haystack: Undetectability of Mycorrhizal Fungi Outside of the Plant Rhizosphere Associated with an Endangered Australian Orchid. Fungal Ecol. 2018, 33, 13–23. [Google Scholar] [CrossRef]
- Chen, L.; Wang, Y.C.; Qin, L.Y.; He, H.Y.; Yu, X.L.; Yang, M.Z.; Zhang, H.B. Dynamics of Fungal Communities during Gastrodia elata Growth. BMC Microbiol. 2019, 19, 1–11. [Google Scholar] [CrossRef]
- Oja, J.; Kohout, P.; Tedersoo, L.; Kull, T.; Kõljalg, U. Temporal Patterns of Orchid Mycorrhizal Fungi in Meadows and Forests as Revealed by 454 Pyrosequencing. New Phytol. 2015, 205, 1608–1618. [Google Scholar] [CrossRef]
- Valdés, M.; Bautista Guerrero, H.; Martínez, L.; Víquez, R. The Root Colonizing Fungi of the Terrestrial Orchid Cypripedium irapeanum. Lankesteriana 2011, 11, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Zelmer, C.D.; Cuthbertson, L.; Currah, R.S. Fungi Associated with Terrestrial Orchid Mycorrhizas, Seeds and Protocorms. Mycoscience 1996. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T. ITS Primers with Enhanced Specifity for Basidiomycetes—Application to the Identification of Mycorrhizae and Rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Guindon, S.; Gascuel, O. A Simple, Fast, and Accurate Algorithm to Estimate Large Phylogenies by Maximum Likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guindon, S.; Dufayard, J.-F.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Bell, J.; Yokoya, K.; Kendon, J.P.; Sarasan, V. Diversity of Root-Associated Culturable Fungi of Cephalanthera rubra (Orchidaceae) in Relation to Soil Characteristics. PeerJ 2020, 2020, e8695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramsay, M.M.; Stewart, J. Re-Establishment of the Ladt’s Slipper Orchid (Cypripedium calceolus L.) in Britain. Bot. J. Linn. Soc. 1998, 126, 173–181. [Google Scholar]
- Barsberg, S.; Rasmussen, H.N.; Kodah, N. Composition of Cypripedium calceolus (Orchidaceae) Seeds Analyzed by Attenuated Total Reflectance Ir Spectroscopy: In Search of Understanding Longevity in the Ground. Am. J. Bot. 2013, 100, 2066–2073. [Google Scholar] [CrossRef]
- Zeng, S.; Zhang, Y.; Teixeira da Silva, J.A.; Wu, K.; Zhang, J.; Duan, J. Seed Biology and in vitro Seed Germination of Cypripedium. Crit. Rev. Biotechnol. 2014, 34, 358–371. [Google Scholar] [CrossRef]
- Lee, Y.I.; Chung, M.C.; Yeung, E.C.; Lee, N. Dynamic Distribution and the Role of Abscisic Acid during Seed Development of a Lady’s Slipper Orchid, Cypripedium formosanum. Ann. Bot. 2015, 116, 403–411. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Lee, Y.I.; Deng, L.; Zhao, S. Asymbiotic Germination of Immature Seeds and the Seedling Development of Cypripedium macranthos Sw., an Endangered Lady’s Slipper Orchid. Sci. Hortic. 2013, 164, 130–136. [Google Scholar] [CrossRef]
- Pedersen, H.A.; Rasmussen, H.N.; Kahandawala, I.M.; Fay, M.F. Genetic Diversity, Compatibility Patterns and Seed Quality in Isolated Populations of Cypripedium calceolus (Orchidaceae). Conserv. Genet. 2012, 13, 89–98. [Google Scholar] [CrossRef]
- Jiang, H.; Chen, M.C.; Lee, Y.I. In vitro Germination and Low-Temperature Seed Storage of Cypripedium lentiginosum P.J. Cribb & S.C. Chen, a Rare and Endangered Lady’s Slipper Orchid. Sci. Hortic. 2017, 225, 471–479. [Google Scholar] [CrossRef]
- SIGMA Phytamax TM. Orchid Maintenance Medium. Available online: https://www.sigmaaldrich.com/catalog/product/sigma/p6668?lang=es®ion=MX (accessed on 26 May 2022).
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Norstog, K. Induction of Embryolike Structures by Kinetin in Cultured Barley Embryos. Dev. Biol. 1970, 23, 665–670. [Google Scholar] [CrossRef]
- Hsu, R.C.C.; Lee, Y.I. Seed Development of Cypripedium debile Rchb. f. in Relation to Asymbiotic Germination. HortScience 2012, 47, 1495–1498. [Google Scholar] [CrossRef] [Green Version]
- Yeung, E.C. A Perspective on Orchid Seed and Protocorm Development Background. Bot. Stud. 2017, 58. [Google Scholar] [CrossRef] [Green Version]
- Fay, M.F. Orchid Conservation: How Can We Meet the Challenges in the Twenty-First Century? Bot. Stud. 2018, 59, 16. [Google Scholar] [CrossRef] [Green Version]
- Jakubska-Busse, A.; Tsiftsis, S.; Śliwiński, M.; Křenová, Z.; Djordjević, V.; Steiu, C.; Kolanowska, M.; Efimov, P.; Hennigs, S.; Lustyk, P.; et al. How to Protect Natural Habitats of Rare Terrestrial Orchids Effectively: A Comparative Case Study of Cypripedium calceolus in Different Geographical Regions of Europe. Plants 2021, 10, 404. [Google Scholar] [CrossRef]
- Swarts, N.D.; Batty, A.L.; Hopper, S.; Dixon, K.W. Does Integrated Conservation of Terrestrial Orchids Work? Lankesteriana 2007, 7, 219–222. [Google Scholar] [CrossRef] [Green Version]
- McCormick, M.K.; Lee Taylor, D.; Juhaszova, K.; Burnett, R.K.; Whigham, D.F.; O’Neill, J.P. Limitations on Orchid Recruitment: Not a Simple Picture. Mol. Ecol. 2012, 21, 1511–1523. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species | Mean Day Growth on PDA Medium | Basal Septa (µm) | Diameter of Hyphae (µm) | Monilioid Cells (µm) | |
|---|---|---|---|---|---|
| Mean ± SD | Width | Length | |||
| Cypripedium irapeanum (Tropical deciduous forest, Veracruz) | 1.14 ± 0.22a | 1.794 ± 1.280a | 3.3 ± 1.1a | 7.7 ± 2.0a | 16.1 ± 4.5a |
| C. irapeanum (Quercus forest, Mexico State) | 0.44 ± 0.25b | 1.439 ± 0.384a | 2.7 ± 0.4a | 7.8 ± 1.2a | 18.5 ± 4.2a |
| C. molle (Quercus forest, Oaxaca) | 2.50 ± 1.20c | 7.368 ± 1.679b | 12.6 ± 1.2b | 20.0 ± 1.9b | 36.2 ± 8.4b |
| Treatments | STAGES (%) | |||||
|---|---|---|---|---|---|---|
| E0 | E1 | E2–3 | E4 | E5 | E6 | |
| Immature and Ungerminated Seeds with Embryos | Imbibition | Rupture of the Testa and Polarization of the Embryo | Foliar Elongation | Root Meristematic Differentiation | Seedling | |
| MS | 49 | 99 | 0 | 1 | 0 | 0 |
| Phytamax ™ | 71 | 88 | 10 | 0 | 2 | 0 |
| Norstog | 52 | 91 | 1 | 3 | 1 | 4 |
| OMA | 49 | 86 | 14 | 0 | 0 | 0 |
| TDF isolate | 29 | 81 | 19 | 1 | 0 | 0 |
| Quercus forest isolate | 26 | 65 | 24 | 11 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno-Camarena, M.; Ortega-Larrocea, M.P. Mesoamerican Cypripedium: Mycorrhizal Contributions to Promote Their Conservation as Critically Endangered Species. Plants 2022, 11, 1554. https://doi.org/10.3390/plants11121554
Moreno-Camarena M, Ortega-Larrocea MP. Mesoamerican Cypripedium: Mycorrhizal Contributions to Promote Their Conservation as Critically Endangered Species. Plants. 2022; 11(12):1554. https://doi.org/10.3390/plants11121554
Chicago/Turabian StyleMoreno-Camarena, Mauricio, and María Pilar Ortega-Larrocea. 2022. "Mesoamerican Cypripedium: Mycorrhizal Contributions to Promote Their Conservation as Critically Endangered Species" Plants 11, no. 12: 1554. https://doi.org/10.3390/plants11121554
APA StyleMoreno-Camarena, M., & Ortega-Larrocea, M. P. (2022). Mesoamerican Cypripedium: Mycorrhizal Contributions to Promote Their Conservation as Critically Endangered Species. Plants, 11(12), 1554. https://doi.org/10.3390/plants11121554