
Physiological and Structural Responses to Prolonged Water Deficit in Young Trees of Two Olive Cultivars


 , , ,
, , , 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Plant Water Status and Gas Exchange
2.2.1. Leaf Stomatal Conductance
2.2.2. Stem Water Potential
2.2.3. Transpiration, Photosynthesis, and Water Use Efficiency
2.3. Trunk Cross-Sectional Area and Stem and Hydraulic Conductance
2.4. Diel Dynamics of Trunk Diameter and Leaf Turgor Pressure
2.5. Statistical Analysis
3. Results and Discussion
3.1. Greenhouse Environmental Conditions and Irrigation
3.2. Plant Water Status and Gas Exchange
3.2.1. Leaf Stomatal Conductance and Stem Water Potential
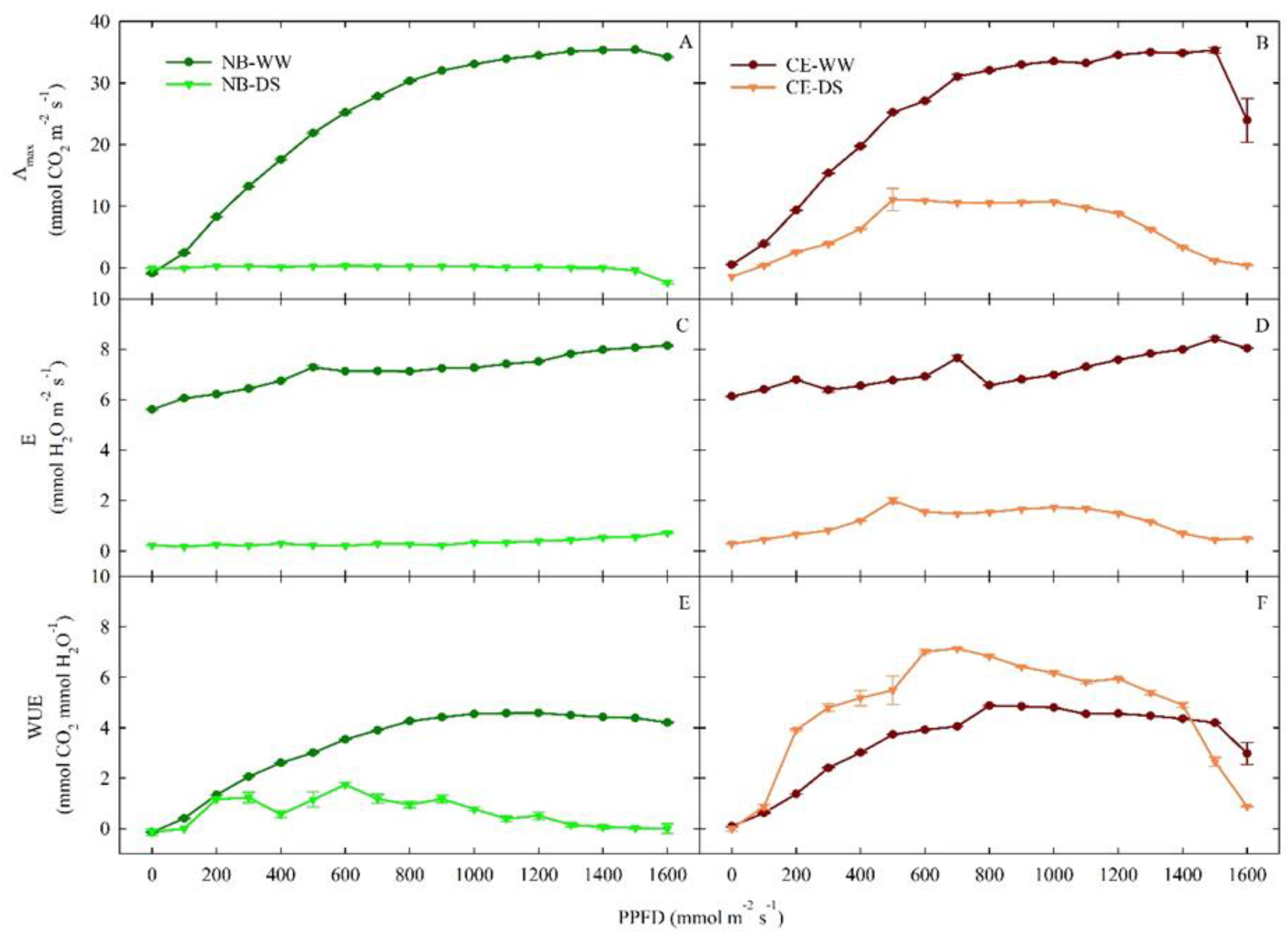
3.2.2. Transpiration, Photosynthesis, and Water Use Efficiency
3.3. Trunk Cross-Sectional Area and Stem and Root Hydraulic Conductance
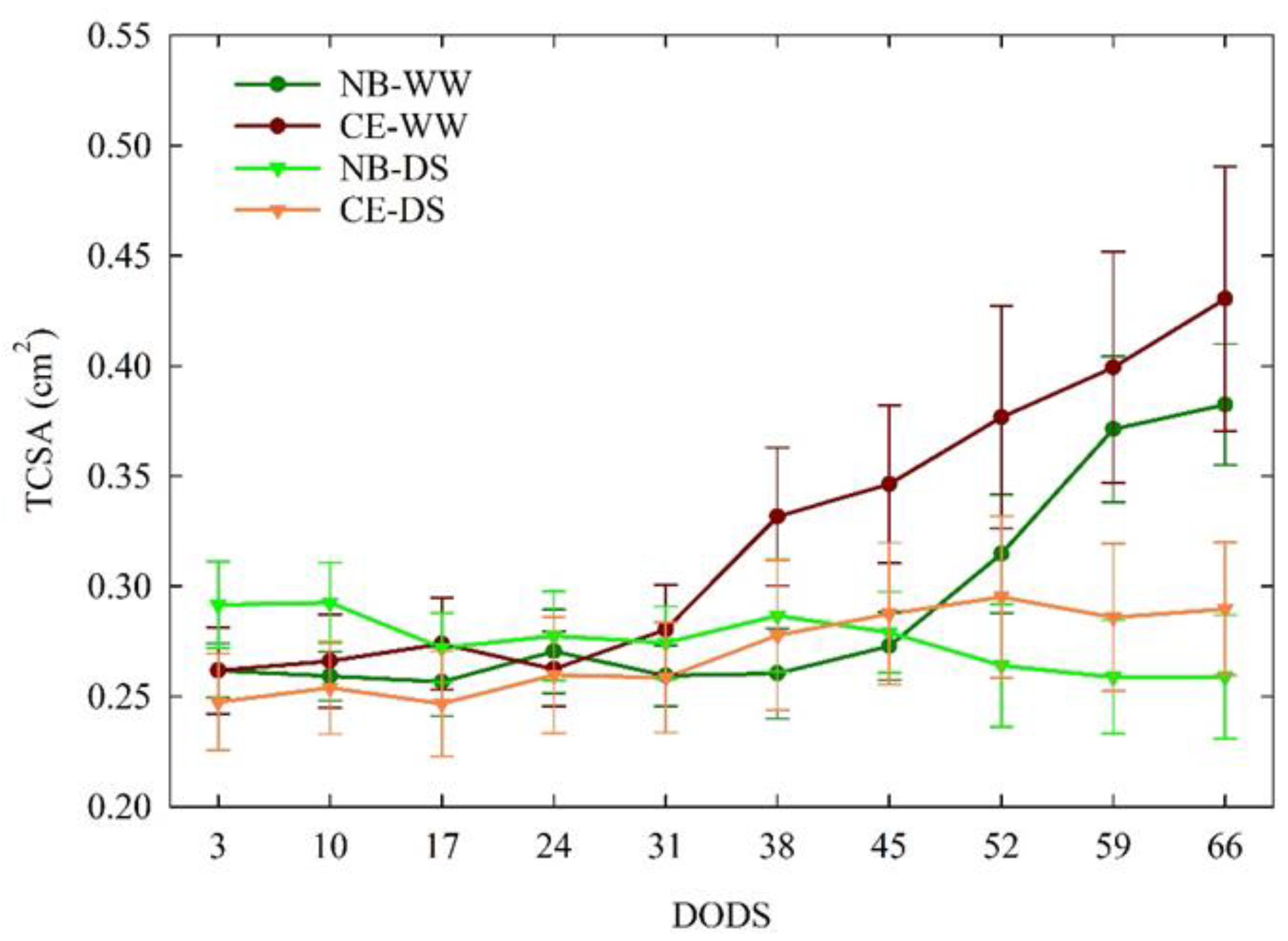
3.3.1. Trunk Cross-Sectional Area
3.3.2. Hydraulic Conductance
3.4. Diel Dynamics of Trunk Diameter and Leaf Turgor Pressure
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cammarano, D.; Ceccarelli, S.; Grando, S.; Romagosa, I.; Benbelkacem, A.; Akar, T.; Al-Yassin, A.; Pecchioni, N.; Francia, E.; Ronga, D. The impact of climate change on barley yield in the Mediterranean basin. Eur. J. Agron. 2019, 106, 1–11. [Google Scholar] [CrossRef]
- Linares, C.; Díaz, J.; Negev, M.; Martínez, G.S.; Debono, R.; Paz, S. Impacts of climate change on the public health of the Mediterranean Basin population-current situation, projections, preparedness and adaptation. Environ. Res. 2020, 182, 109107. [Google Scholar] [CrossRef] [PubMed]
- Schwabe, W.W.; Lionakis, S.M. Leaf attitude in olive in relation to drought resistance. J. Hortic. Sci. 1996, 71, 157–166. [Google Scholar] [CrossRef]
- Tous, J.; Hermoso, J.F.; Romero, A. New trends in olive orchard design for continuous mechanical harvesting. Adv. Hort. Sci. 2010, 24, 43–52. [Google Scholar]
- Motilva, M.J.; Tovar, M.J.; Romero, M.P.; Alegre, S.; Girona, J. Influence of regulated deficit irrigation strategies applied to olive trees (Arbequina cultivar) on oil yield and oil composition during the fruit ripening period. J. Sci. Food Agric. 2000, 80, 2037–2043. [Google Scholar] [CrossRef]
- Patumi, M.; D’andria, R.; Marsilio, V.; Fontanazza, G.; Morelli, G.; Lanza, B. Olive and oil quality after intensive monocone olive growing (Olea europaea L. cv. Kalamata) in different irrigation regimes. Food Chem. 2002, 77, 27–34. [Google Scholar] [CrossRef]
- Moriana, A.; Orgaz, F.; Pastor, M.; Fereres, E. Yield responses of a mature olive orchard to water deficits. J. Am. Soc. Hortic. Sci. 2003, 128, 425–431. [Google Scholar] [CrossRef]
- Gucci, R.; Lodolini, E.; Rapoport, H.F. Productivity of olive trees with different water status and crop load. J. Hort. Science Biotech. 2007, 82, 648–656. [Google Scholar] [CrossRef]
- Servili, M.; Esposto, S.; Lodolini, E.; Selvaggini, R.; Taticchi, A.; Urbani, S.; Montedoro, G.F.; Serravalle, M.; Gucci, R. Irrigation effects on quality, phenolic composition and selected volatiles of virgin olive oil cv Leccino. J. Agric. Food Chem. 2007, 55, 6609–6618. [Google Scholar] [CrossRef]
- Pannelli, G.; Servili, M.; Baldioli, M.; Selvaggini, R.; Montedoro, G.F. Effect of agronomic and seasonal factors on olive (Olea europaea L.) production and on the qualitative characterization of the oil. Acta Hortic. 1994, 356, 239–243. [Google Scholar] [CrossRef]
- Rieger, M. Offsetting effects of reduced root hydraulic conductivity and osmotic adjustment following drought. Tree Physiol. 1995, 15, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Chartzoulakis, K.; Patakas, A.; Bosabalidis, A.M. Changes in water relations, photosynthesis and leaf anatomy induced by intermittent drought in two olive cultivars. Environ. Exp. Bot. 1999, 42, 113–120. [Google Scholar] [CrossRef]
- Xiloyannis, C.; Dichio, B.; Nuzzo, V.; Celano, G. Defense strategies of olive against water stress. Acta Hortic. 1999, 474, 423–426. [Google Scholar] [CrossRef]
- Dichio, B.; Xiloyannis, C.; Angelopoulos, K.; Nuzzo, V.; Bufo, S.A.; Celano, G. Drought-induced variations of water relations parameters in Olea europaea. Plant Soil 2003, 257, 381–389. [Google Scholar] [CrossRef]
- Dichio, B.; Sofo, A.; Xiloyannis, C. Adaptive mechanisms of olive tree to drought. In Proceedings of the InterDrought-II-The 2nd International Conference on Integrated Approaches to Sustain and Improve Plant Production Under Drought Stress, Rome, Italy, 24–28 September 2005. [Google Scholar]
- Lo Bianco, R.; Scalisi, A. Water relations and carbohydrate partitioning of four greenhouse-grown olive genotypes under long-term drought. Trees 2017, 31, 717–727. [Google Scholar] [CrossRef]
- Abd-El-Rahman, A.A.; El-Sharkawi, H.M. Response of olive and almond orchards to partial irrigation under dry-farming practices in semi-arid regions. Plant Soil 1974, 41, 13–31. [Google Scholar] [CrossRef]
- Sofo, A.; Dichio, B.; Xiloyannis, C.; Masia, A. Effects of different irradiance levels on some antioxidant enzymes and on malondialdehyde content during rewatering in olive tree. Plant Sci. 2004, 166, 293–302. [Google Scholar] [CrossRef]
- Fernández, J.E.; Moreno, F.; Girón, I.F.; Blázquez, O.M. Stomatal control of water use in olive tree leaves. Plant Soil 1997, 190, 179–192. [Google Scholar] [CrossRef] [Green Version]
- Angelopoulos, K.; Dichio, B.; Xiloyannis, C. Inhibition of photosynthesis in olive tree (Olea europaea L.) during water stress and rewatering. J. Exp. Bot. 1996, 47, 1093–1100. [Google Scholar] [CrossRef] [Green Version]
- Xiloyannis, C.; Dichio, B.; Nuzzo, V.; Celano, G. L’olivo: Pianta esempio per la sua capacità di resistenza in condizioni di estrema siccità. In Seminari di Olivicoltura; Accademia Nazionale Olivo e dell’Olio: Spoleto (PG), Italy, 1998; pp. 79–111. [Google Scholar]
- Dichio, B.; Xiloyannis, C.; Sofo, A.; Montanaro, G. Osmotic adjustment in leaves and roots of olive tree (Olea europaea L.) during drought stress and rewatering. Tree Physiol. 2006, 26, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Sofo, A.; Nuzzo, V.; Palese, A.M.; Xiloyannis, C.; Celano, G.; Zukowskyj, P.; Dichio, B. Net CO2 storage in Mediterranean olive and peach orchards. Sci. Hortic. 2005, 107, 17–24. [Google Scholar] [CrossRef]
- Fernández, J.E.; Moreno, F.; Martin-Aranda, J.; Rapoport, H.F. Anatomical response of olive roots to dry and irrigated soils. Adv. Hortic. Sci. 1994, 8, 141–144. [Google Scholar]
- Bosabalidis, A.M.; Kofidis, G. Comparative effects of drought stress on leaf anatomy of two olive cultivars. Plant Sci. 2002, 163, 375–379. [Google Scholar] [CrossRef]
- Lo Bianco, R.; Losciale, P.; Manfrini, L.; Corelli Grappadelli, L. Possible role of mannitol as an oxygen radical scavenger in olive. Acta Hortic. 2011, 924, 83–88. [Google Scholar] [CrossRef]
- Ennajeh, M.; Tounekti, T.; Vade, A.M.; Khemira, H.; Cochard, H. Water relations and drought-induced embolism in olive (Olea europaea) varieties ‘Meski’ and ‘Chemlali’. Tree Physiol. 2008, 28, 971–976. [Google Scholar] [CrossRef]
- Gucci, R. Implicazioni delle relazioni idriche nella gestione dell’irrigazione dell’olivo. Italus Hortus 2006, 13, 61–70. [Google Scholar]
- Sperry, J.S. Hydraulic constraints on plant gas exchange. Agric. For. Meteorol. 2000, 104, 13–23. [Google Scholar] [CrossRef]
- Meinzer, F.C. Co-ordination of vapor and liquid phase water transport properties in plants. Plant Cell Environ. 2002, 25, 265–274. [Google Scholar] [CrossRef]
- Dichio, B.; Romano, M.; Nuzzo, V.; Xiloyannis, C. Soil wateravailability and relationship between canopy and roots in youngolive trees (cv Coratina). Acta Hortic. 2002, 586, 255–258. [Google Scholar] [CrossRef]
- Xiloyannis, C.; Gucci, R.; Dichio, B. Irrigazione. Olea Trattato Olivic. 2003, 365–389. [Google Scholar]
- Caruso, T.; Marra, F.P.; Costa, F.; Campisi, G.; Macaluso, L.; Marchese, A. Genetic diversity and clonal variation within the main Sicilian olive cultivars based on morphological traits and microsatellite markers. Sci. Hortic Engl. 2014, 180, 130–138. [Google Scholar] [CrossRef]
- Lo Bianco, R.; Avellone, G. Diurnal regulation of leaf water status in high-and low-mannitol olive cultivars. Plants 2014, 3, 196–208. [Google Scholar] [CrossRef] [Green Version]
- Marra, F.P.; Caruso, T.; Costa, F.; Di Vaio, C.; Mafrica, R.; Marchese, A. Genetic relationships, structure and parentage simulation among the olive tree (Olea europaea L. subsp. europaea) cultivated in Southern Italy revealed by SSR markers. Tree Genet. Genomes 2013, 9, 961–973. [Google Scholar] [CrossRef]
- Grisafi, F.; Bonafede, E.; Vecchia, F.D.; Rascio, N. Some morphological, anatomical, physiological responses of different olive cultivars to high temperatures and drought stress. Acta Bot. Gallica 2004, 151, 241–253. [Google Scholar] [CrossRef] [Green Version]
- Scalisi, A.; Marra, F.P.; Caruso, T.; Illuminati, C.; Costa, F.; Lo Bianco, R. Transpiration rates and hydraulic conductance of two olive genotypes with different sensitivity to drought. Acta Hort. 2019, 1253, 421–428. [Google Scholar] [CrossRef]
- Turner, N.C. Measurement of plant water status by the pressure chamber technique. Irrig. Sci. 1988, 9, 289–308. [Google Scholar] [CrossRef]
- Tyree, M.T.; Yang, S.; Cruiziat, P.; Sinclair, B. Novel Methods of Measuring Hydraulic Conductivity of Tree Root Systems and Interpretation Using AMAIZED (A Maize-Root Dynamic Model for Water and Solute Transport). Plant Physiol. 1994, 104, 189–199. [Google Scholar] [CrossRef] [Green Version]
- Scalisi, A.; Morandi, B.; Inglese, P.; Lo Bianco, R. Cladode growth dynamics in Opuntia ficus-indica under drought. Environ. Exp. Bot. 2016, 122, 158–167. [Google Scholar] [CrossRef]
- Zimmermann, D.; Reuss, R.; Westhoff, M.; Geßner, P.; Bauer, W.; Bamberg, E.; Bentrup, F.W.; Zimmermann, U. A novel, non-invasive, online-monitoring, versatile and easy plant-based probe for measuring leaf water status. J. Exp. Bot. 2008, 59, 3157–3167. [Google Scholar] [CrossRef] [Green Version]
- Scalisi, A.; Marino, G.; Marra, F.P.; Caruso, T.; Lo Bianco, R. A Cultivar-Sensitive Approach for the Continuous Monitoring of Olive (Olea europaea L.) Tree Water Status by Fruit and Leaf Sensing. Front. Plant. Sci. 2020, 11, 340. [Google Scholar] [CrossRef]
- Marino, G.; Scalisi, A.; Guzmán-Delgado, P.; Caruso, T.; Marra, F.P.; Lo Bianco, R. Detecting mild water stress in olive with multiple plant-based continuous sensors. Plants 2021, 10, 131. [Google Scholar] [CrossRef] [PubMed]
- Savitzky, A.; Golay, M.J. Smoothing and differentiation of data by simplified least squares procedures. Anal. Chem. 1964, 36, 1627–1639. [Google Scholar] [CrossRef]
- Parent, B.; Hachez, C.; Redondo, E.; Simonneau, T.; Chaumont, F.; Tardieu, F. Drought and abscisic acid effects on aquaporin content translate into changes in hydraulic conductivity and leaf growth rate: A trans-scale approach. Plant Physiol 2009, 149, 2000–2012. [Google Scholar] [CrossRef] [Green Version]
- Tognetti, R.; D’Andria, R.; Lavini, A.; Morelli, G. The effect of deficit irrigation on crop yield and vegetative development of Olea europaea L. (cvs. Frantoio and Leccino). Eur. J. Agron. 2006, 25, 356–364. [Google Scholar] [CrossRef]
- Flexas, J.; Medrano, H. Drought-inhibition of photosynthesis in C3 plants: Stomatal and non-stomatal limitation revisited. Ann. Bot. 2002, 89, 183–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavel, E.W.; Fereres, E. Low soil temperatures induce water deficits in olive (Olea europaea) trees. Physiol. Plantarum 1998, 104, 525–532. [Google Scholar] [CrossRef]
- Sofo, A.; Dichio, B.; Montanaro, G.; Xiloyannis, C. Photosynthetic performance and light response of two olive cultivars under different water and light regimes. Photosynthetica 2009, 47, 602–608. [Google Scholar] [CrossRef]
- Bhusal, N.; Lee, M.; Han, A.R.; Han, A.; Kim, H.S. Responses to drought stress in Prunus sargentii and Larix kaempferi seedlings using morphological and physiological parameters. Forest Ecol. Manag. 2020, 465, 118099. [Google Scholar] [CrossRef]
- Melaouhi, A.; Baraza, E.; Escalona, J.M.; El-AouOuad, H.; Mahjoub, I.; Bchir, A.; Braham, M.; Bota, J. Physiological and biochemical responses to water deficit and recovery of two olive cultivars (Olea europaea L., Arbequina and Empeltre cvs.) under Mediterranean conditions. Theor. Exp. Plant Phys. 2021, 33, 369–383. [Google Scholar] [CrossRef]
- Kirch, H.; Vera-Estrella, R.; Golldack, D.; Quigley, F.; Michalowski, C.; Barkla, B.; Bohnert, H. Expression of water channel proteins in Mesembryanthemum crystallinum. Plant Physiol. 2000, 123, 111–124. [Google Scholar] [CrossRef] [Green Version]
- Bhusal, N.; Han, S.G.; Yoon, T.M. Impact of drought stress on photosynthetic response, leaf water potential, and stem sap flow in two cultivars of bi-leader apple trees (Malus x domestica Borkh.). Sci. Hortic. 2019, 246, 535–543. [Google Scholar] [CrossRef]
- Bhusal, N.; Bhusal, S.J.; Yoon, T.M. Comparisons of physiological and anatomical characteristics between two cultivars in bi-leader apple trees (Malus x domestica Borkh.). Sci. Hortic. 2018, 231, 73–81. [Google Scholar] [CrossRef]
- Lo Gullo, M.A.; Nardini, A.; Salleo, S.; Tyree, M.T. Changes in root hydraulic conductance (KR) of Olea oleaster seedlings following drought stress and irrigation. New Phytol. 1998, 140, 25–31. [Google Scholar] [CrossRef]
- Clarkson, D.T.; Carvajal, M.; Henzler, T.; Waterhouse, R.N.; Smyth, A.J.; Cooke, D.T.; Steudle, E. Root hydraulic conductance: Diurnal aquaporin expression and the effects of nutrient stress. J. Exp. Bot. 2000, 51, 61–70. [Google Scholar] [CrossRef]
- Tournaire-Roux, C.; Sutka, M.; Javot, H.; Gout, E.; Gerbeau, P.; Trung Luu, D.; Bligny, R.; Maurel, C. Cytosolic pH regulates root water transport during anoxic stress through gating of aquaporins. Nature 2003, 425, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Boursiac, Y.; Chen, S.; Luu, D.T.; Sorieul, M.; Van Den Dries, N.; Maurel, C. Early effects of salinity on water transport in Arabidopsis roots. Molecular and cellular features of aquaporin expression. Plant Physiol. 2005, 139, 790–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bramley, H. Water Flow in the Roots of Three Crop Species: The Influence of Root Structure, Aquaporin Activity and Waterlogging. Ph.D. Thesis, The University of Western Australia, Perth, Australia, 2006. [Google Scholar]
- Afzal, Z.; Howton, T.C.; Sun, Y.; Mukhtar, M.S. The roles of aquaporins in plant stress responses. J. Dev. Biol. 2016, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Secchi, F.; Lovisolo, C.; Schubert, A. Expression of OePIP2.1 aquaporin gene and water relations of Olea europaea twigs during drought stress and recovery. Ann. Appl. Biol. 2007, 150, 163–167. [Google Scholar] [CrossRef]
- Steudle, E. Water uptake by roots: Effects of water deficit. J. Exp. Bot. 2000, 51, 1531–1542. [Google Scholar] [CrossRef] [Green Version]
- Scalisi, A.; Bresilla, K.; Simões Grilo, F. Continuous determination of fruit tree water-status by plant-based sensors. Italus Hortus 2017, 24, 39–50. [Google Scholar] [CrossRef]
- Gucci, R.; Caruso, G.; Gennai, C.; Esposto, S.; Urbani, S.; Servili, M. Fruit growth, yield and oil quality changes induced by deficit irrigation at different stages of olive fruit development. Agric. Water Manag. 2019, 212, 88–98. [Google Scholar] [CrossRef]
- Ehrenberger, W.; Rüger, S.; Rodríguez-Domínguez, C.M.; Díaz-Espejo, A.; Fernández, J.E.; Moreno, D.; Zimmermann, V.; Sukhorukov, L.; Zimmermann, U. Leaf patch clamp pressure probe measurements on olive leaves in a nearly turgorless state. Plant Biol. 2012, 14, 666–674. [Google Scholar] [CrossRef] [PubMed]
- Xiloyannis, C.; Dichio, B.; Nuzzo, V. Meccanismi di risposta dell’olivo alla carenza idrica. In Proceedings of the Atti del convegno Tecniche, Norme e Qualità in Olivicoltura, Potenza, Italy, 15–17 December 1993; pp. 123–136. [Google Scholar]
- Bacelar, E.A.; Moutinho-Pereira, J.M.; Gonçalves, B.C.; Lopes, J.I.; Correia, C.M. Physiological responses of different olive genotypes to drought conditions. Acta Physiol. Plant. 2009, 31, 611–621. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Massenti, R.; Scalisi, A.; Marra, F.P.; Caruso, T.; Marino, G.; Lo Bianco, R. Physiological and Structural Responses to Prolonged Water Deficit in Young Trees of Two Olive Cultivars. Plants 2022, 11, 1695. https://doi.org/10.3390/plants11131695
Massenti R, Scalisi A, Marra FP, Caruso T, Marino G, Lo Bianco R. Physiological and Structural Responses to Prolonged Water Deficit in Young Trees of Two Olive Cultivars. Plants. 2022; 11(13):1695. https://doi.org/10.3390/plants11131695
Chicago/Turabian StyleMassenti, Roberto, Alessio Scalisi, Francesco Paolo Marra, Tiziano Caruso, Giulia Marino, and Riccardo Lo Bianco. 2022. "Physiological and Structural Responses to Prolonged Water Deficit in Young Trees of Two Olive Cultivars" Plants 11, no. 13: 1695. https://doi.org/10.3390/plants11131695
APA StyleMassenti, R., Scalisi, A., Marra, F. P., Caruso, T., Marino, G., & Lo Bianco, R. (2022). Physiological and Structural Responses to Prolonged Water Deficit in Young Trees of Two Olive Cultivars. Plants, 11(13), 1695. https://doi.org/10.3390/plants11131695