
Changes in the Activity of the CLE41/PXY/WOX Signaling Pathway in the Birch Cambial Zone under Different Xylogenesis Patterns

 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Description of 1 and 2 Fractions in Three Phenotypes at Different Dates of Sampling
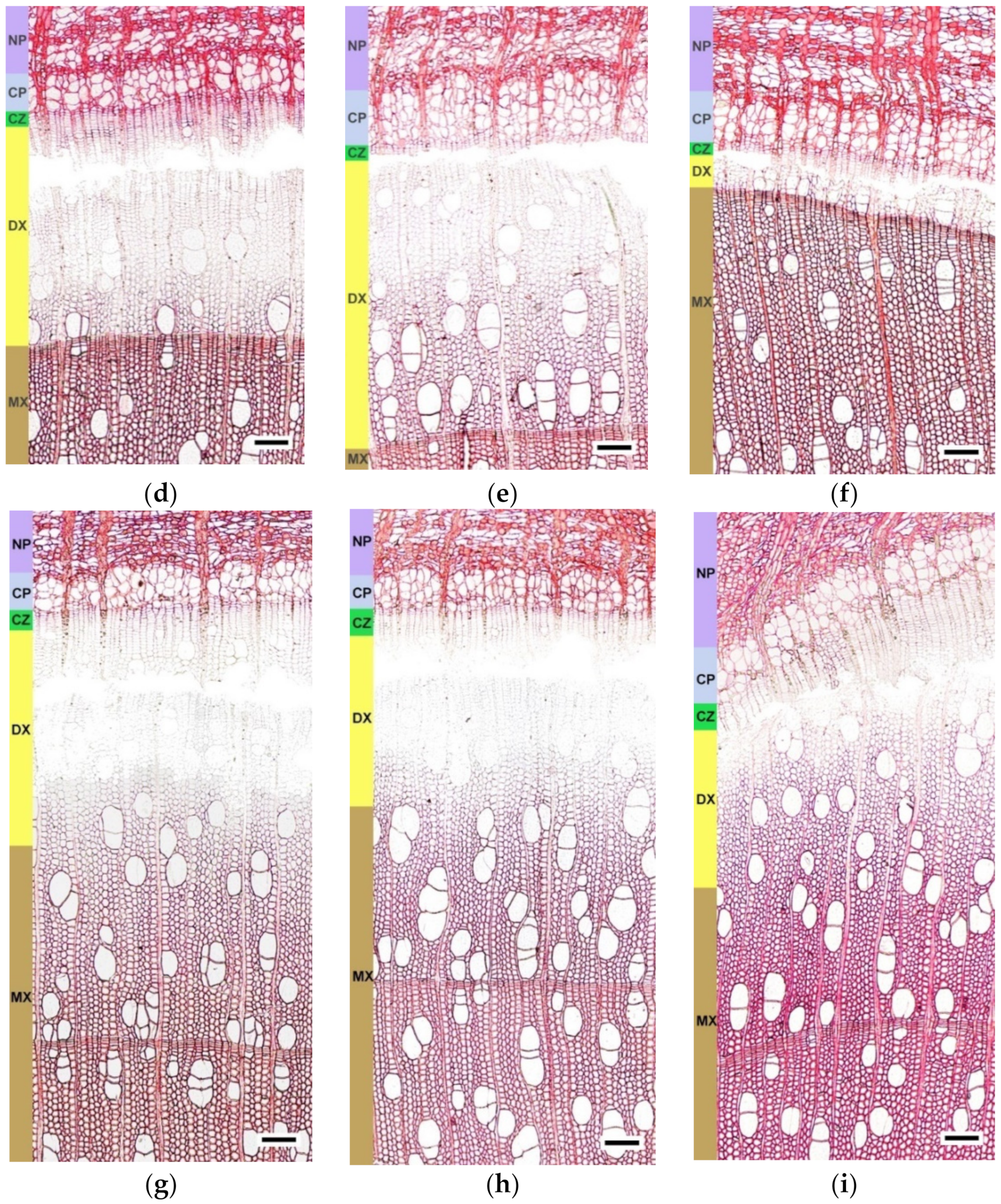
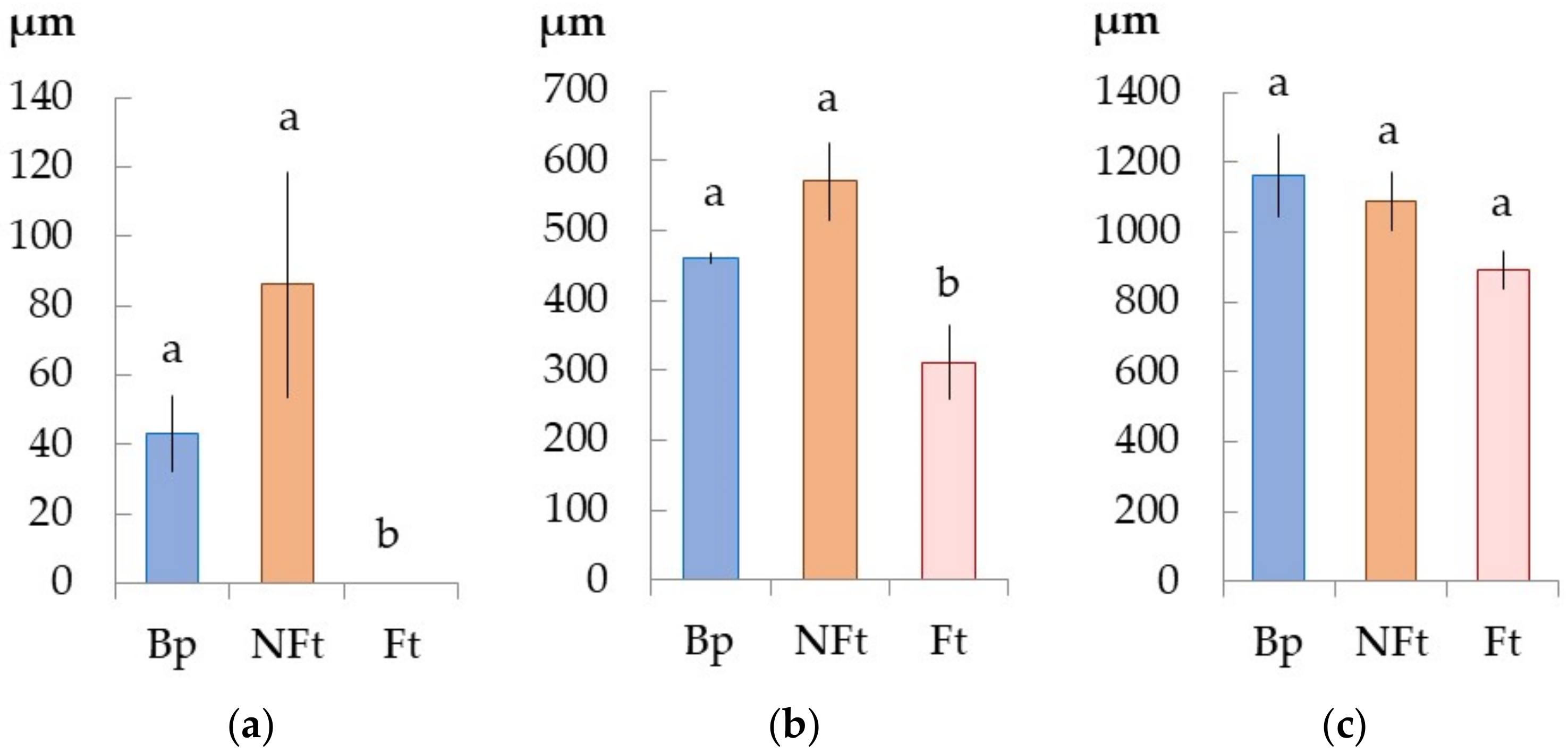
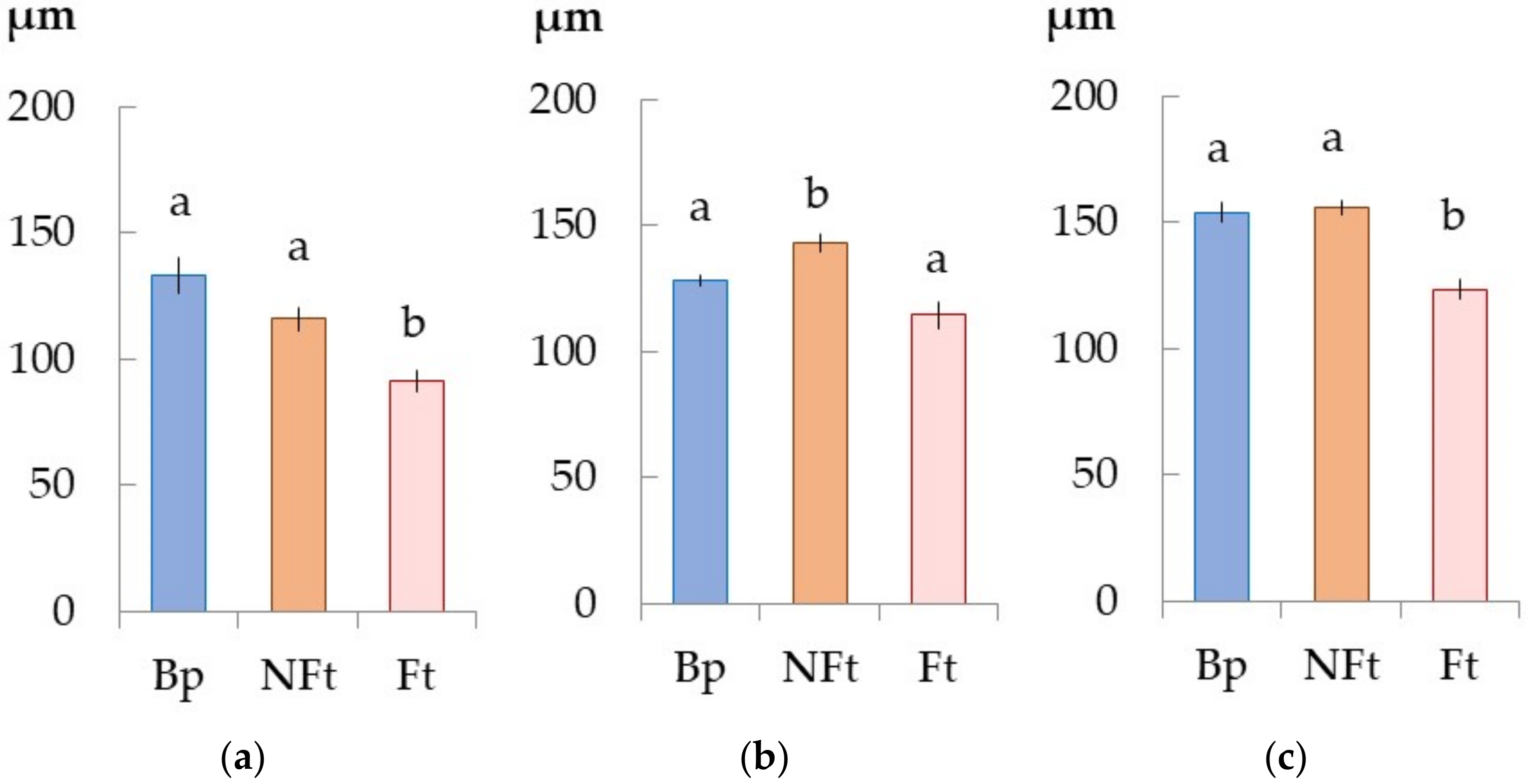
2.2. The Anatomical Features of the Trunk Tissues in Different Forms of Silver Birch
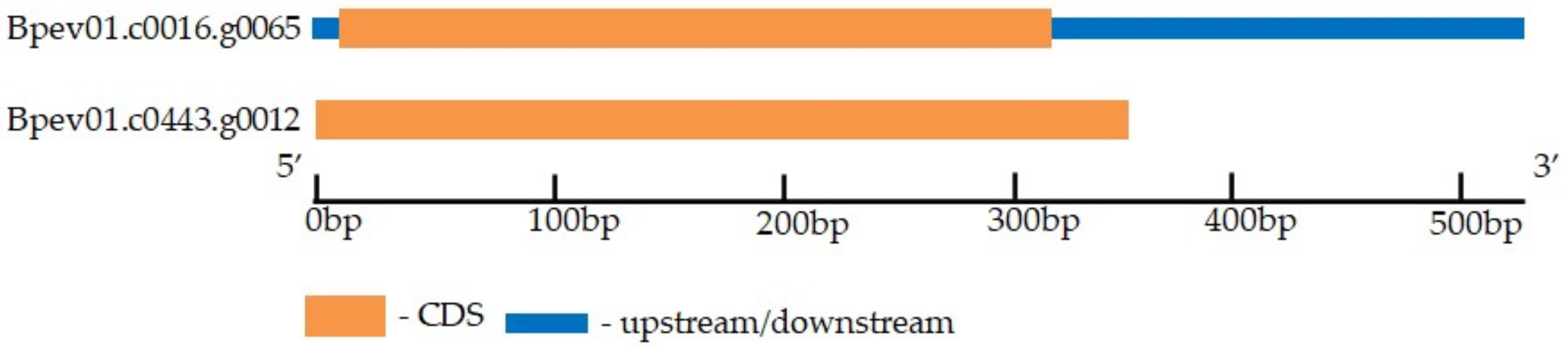
2.3. CLE41/44 Gene Identification in the Silver Birch Genome
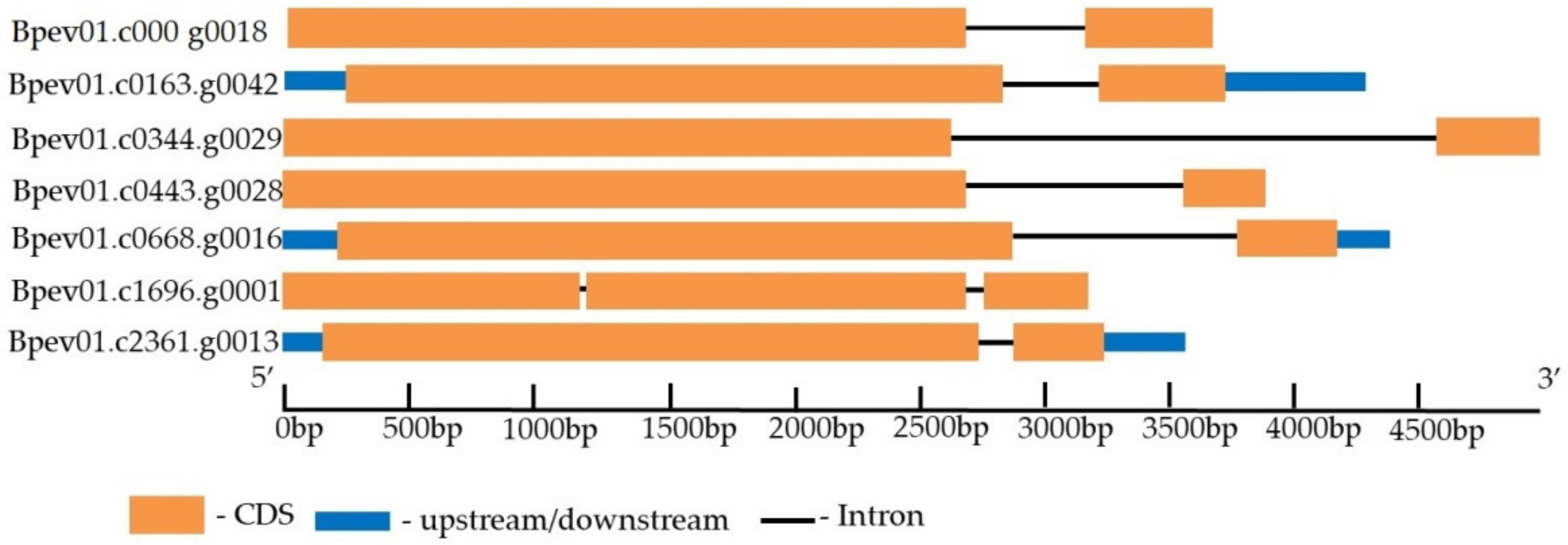
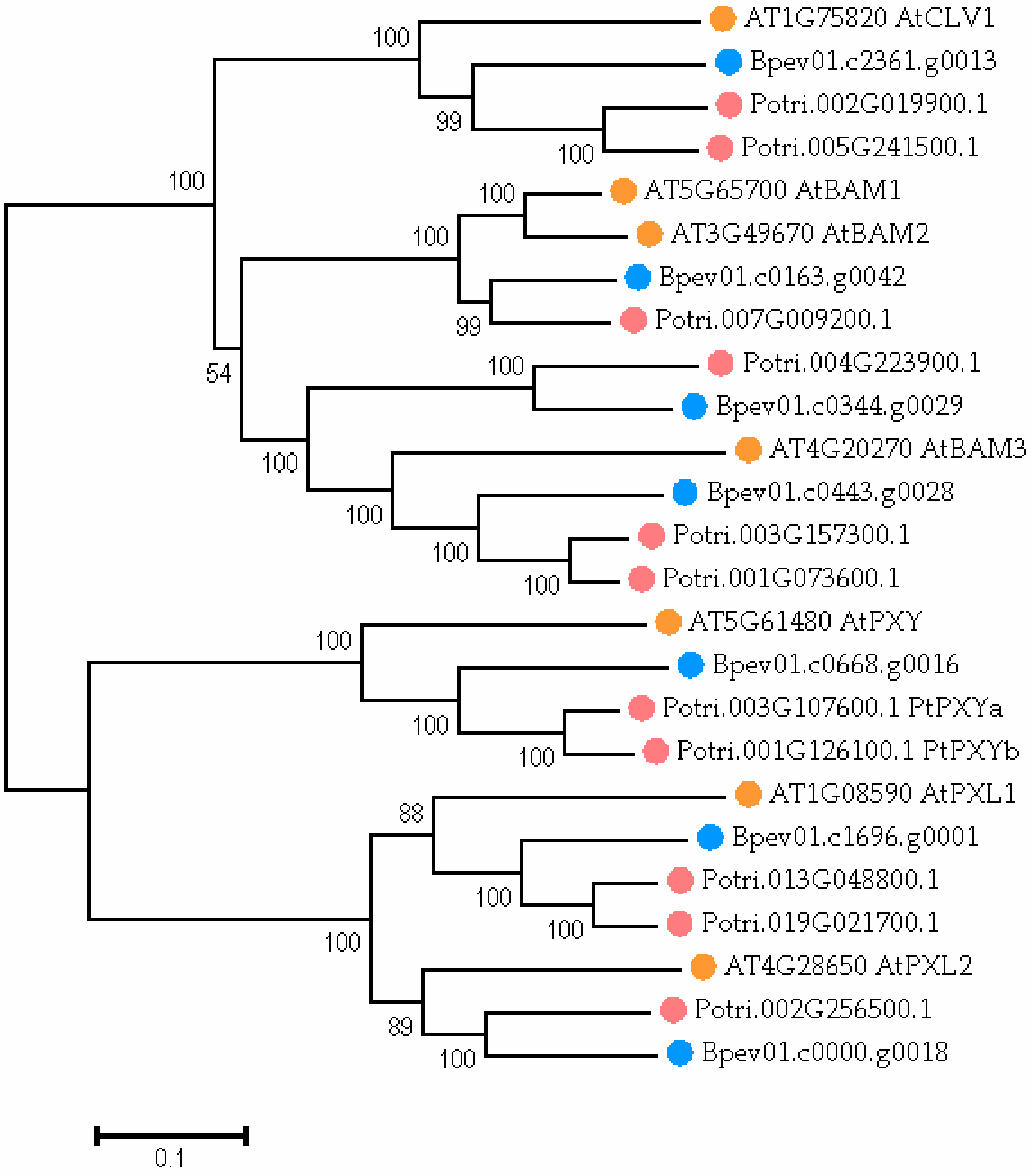
2.4. PXY Gene Identification in the Silver Birch Genome
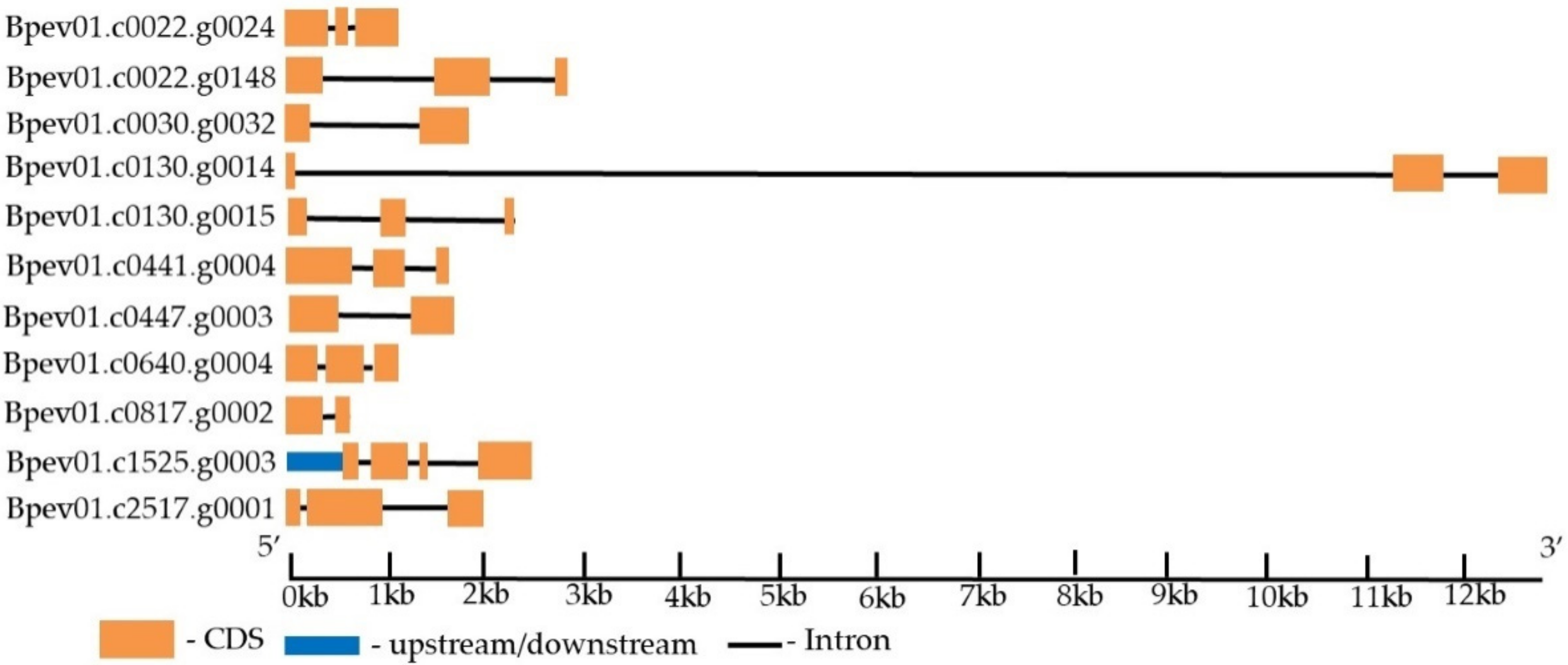
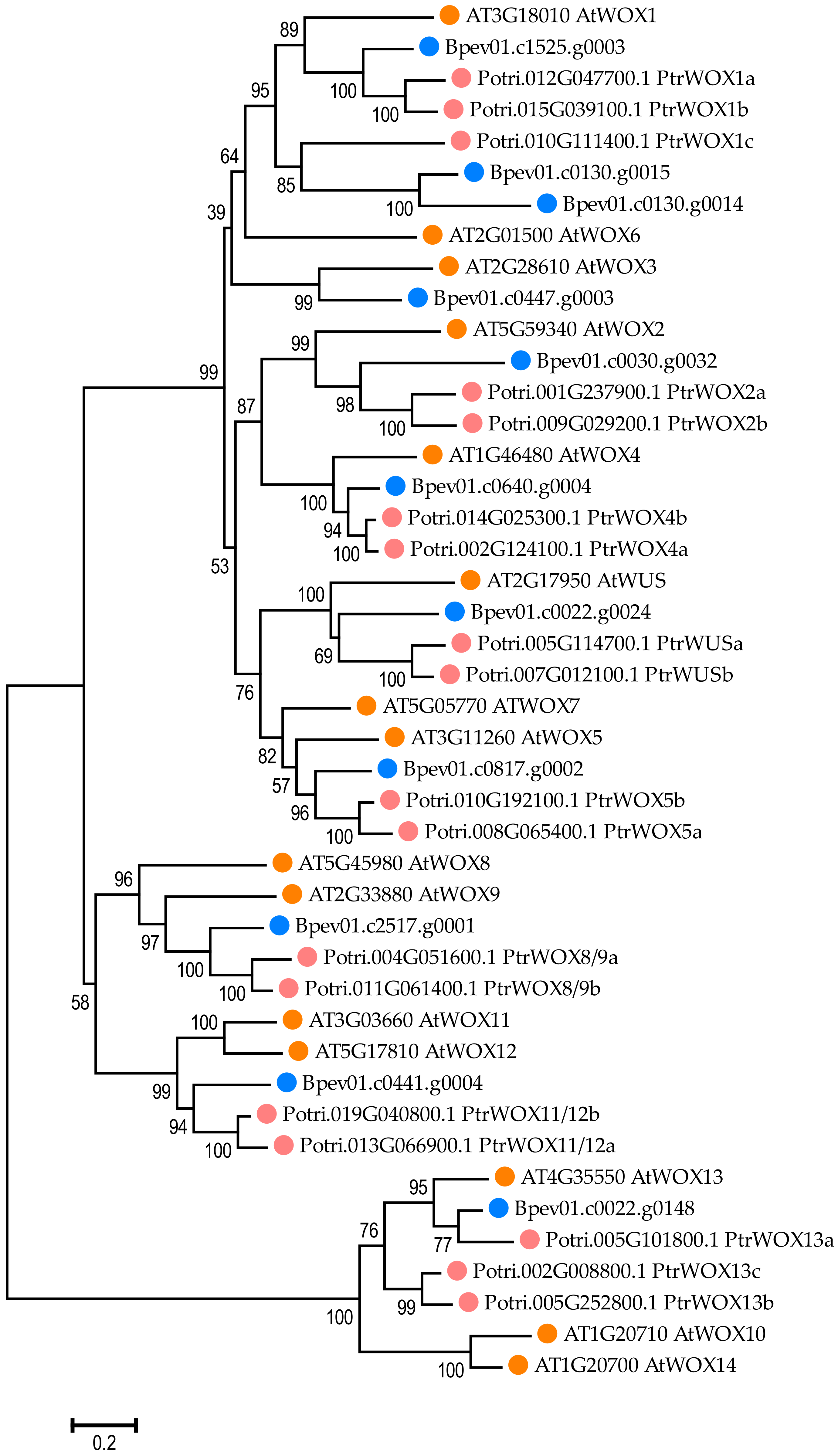
2.5. WOX Gene Identification in the Silver Birch Genome
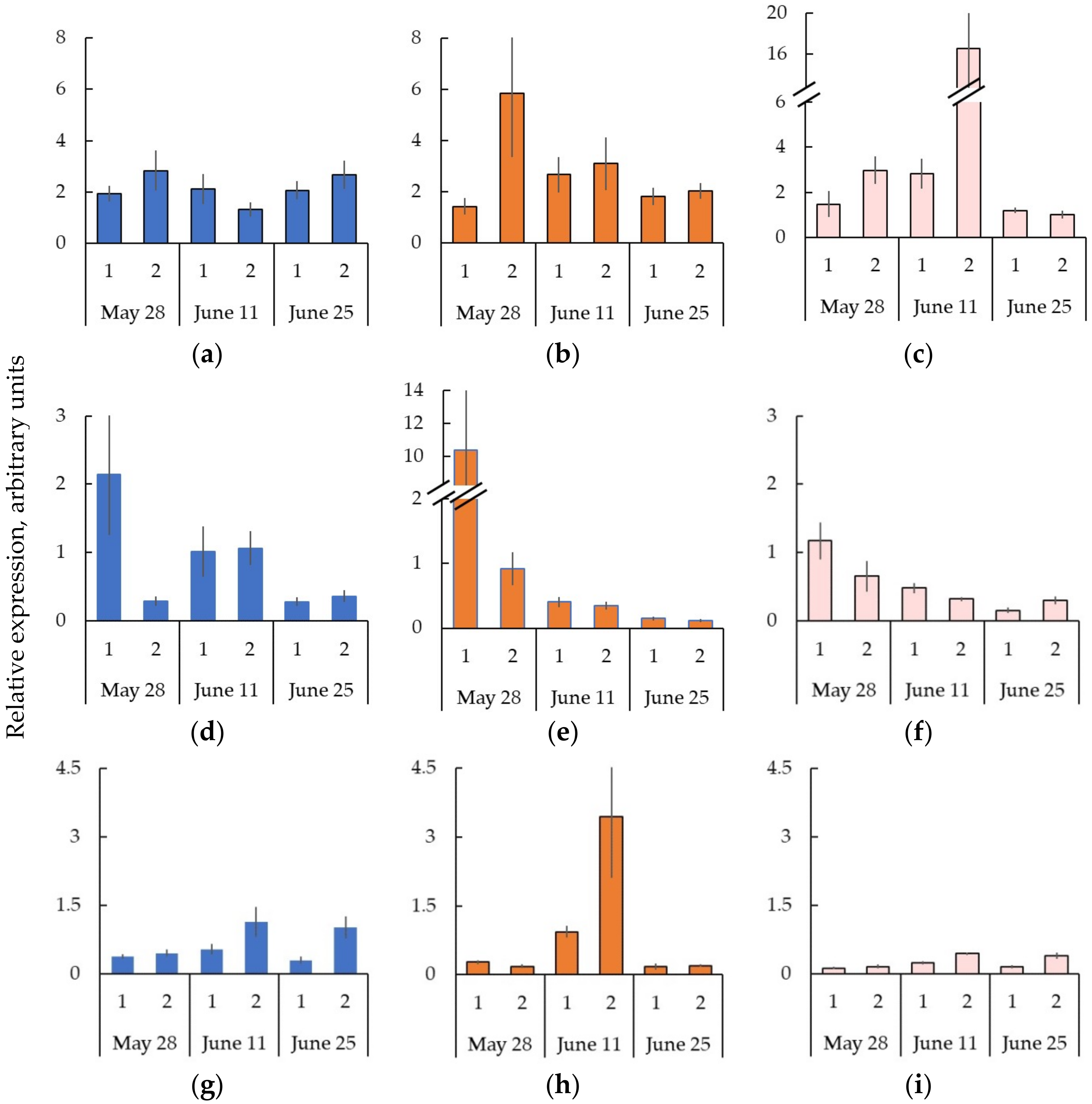
2.6. Expression of the Genes Encoding CLE-PXY-WOX Signaling Pathway in the Trunk Tissues of Bp Trees during Cambial Growth
2.7. The Formation of Straight-Grained Wood of Bc NF Trees Occurs against the Background of Greater Expression of the Genes CLE-PXY-WOX in the Cambial Zone and Differentiating Xylem Compared to the Bp Trees
2.8. Decreased Expression of the Genes CLE-PXY When Forming the Wood Figure in Bc FT Trees
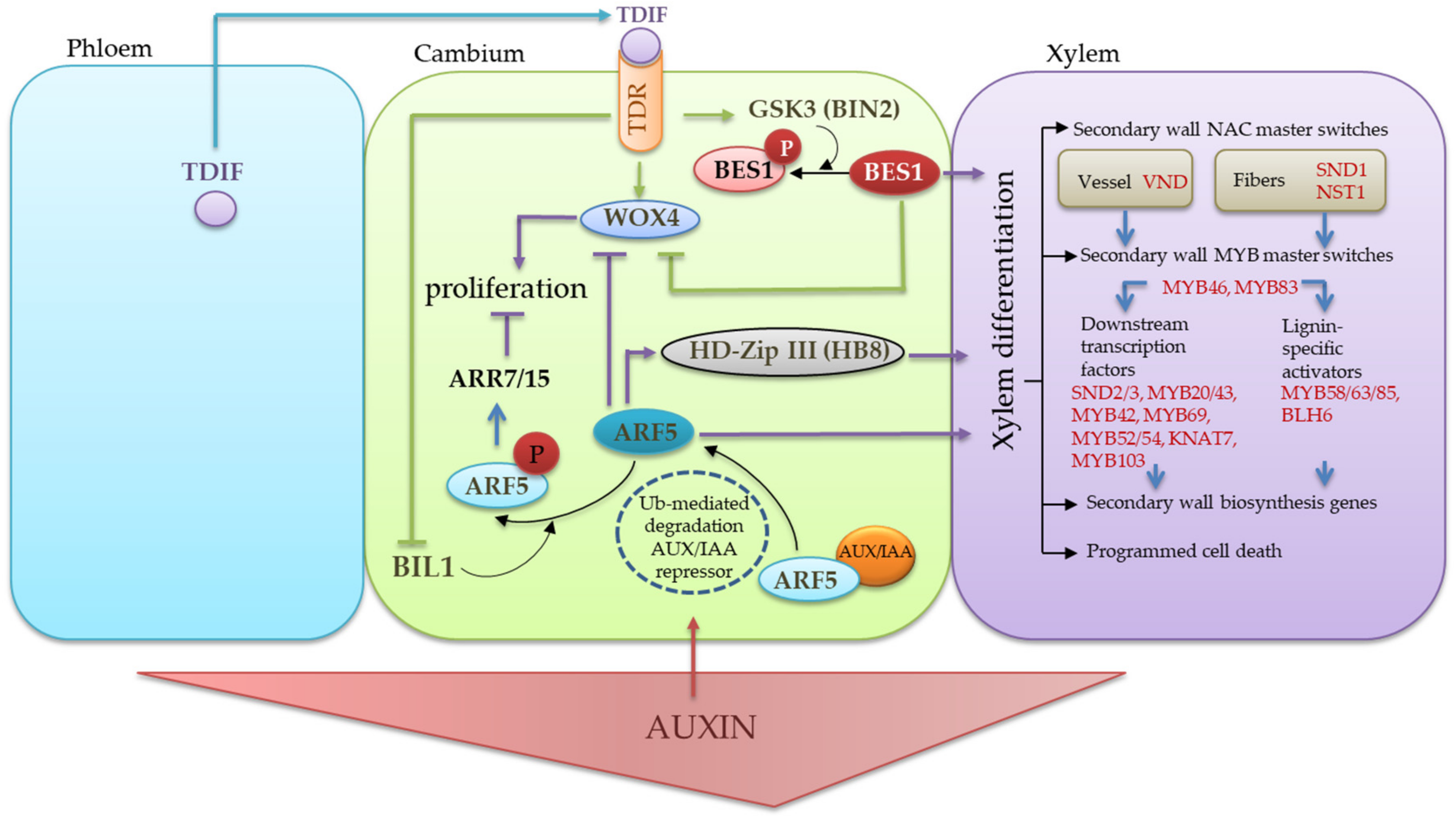
3. Discussion
4. Materials and Methods
4.1. Study Objects
4.2. Plant Sampling
4.3. Microscopy
4.4. Gene Retrieval from the Silver Birch Genome by Bioinformatics Methods
4.5. qRT-PCR
4.6. Statistical Data Processing
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Evert, R.F.; Eichhorn, S.E. Esau’s Plant Anatomy: Meristems, Cells, and Tissues of the Plant Body: Their Structure, Function, and Development, 3rd ed.; Wiley-Interscience: Hoboken, NJ, USA, 2006; ISBN 978-0-471-73843-5. [Google Scholar]
- Fischer, U.; Kucukoglu, M.; Helariutta, Y.; Bhalerao, R.P. The Dynamics of Cambial Stem Cell Activity. Annu. Rev. Plant Biol. 2019, 70, 293–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, D.; Lebovka, I.; López-Salmerón, V.; Sanchez, P.; Greb, T. Bifacial Cambium Stem Cells Generate Xylem and Phloem during Radial Plant Growth. Development 2019, 146, dev171355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smetana, O.; Mäkilä, R.; Lyu, M.; Amiryousefi, A.; Sánchez Rodríguez, F.; Wu, M.-F.; Solé-Gil, A.; Leal Gavarrón, M.; Siligato, R.; Miyashima, S.; et al. High Levels of Auxin Signalling Define the Stem-Cell Organizer of the Vascular Cambium. Nature 2019, 565, 485–489. [Google Scholar] [CrossRef] [PubMed]
- Nieminen, K.; Blomster, T.; Helariutta, Y.; Mähönen, A.P. Vascular Cambium Development. Arab. Book 2015, 13, e0177. [Google Scholar] [CrossRef] [Green Version]
- Schrader, J.; Baba, K.; May, S.T.; Palme, K.; Bennett, M.; Bhalerao, R.P.; Sandberg, G. Polar Auxin Transport in the Wood-Forming Tissues of Hybrid Aspen Is under Simultaneous Control of Developmental and Environmental Signals. Proc. Natl. Acad. Sci. USA 2003, 100, 10096–10101. [Google Scholar] [CrossRef] [Green Version]
- Etchells, J.P.; Turner, S.R. The PXY-CLE41 Receptor Ligand Pair Defines a Multifunctional Pathway That Controls the Rate and Orientation of Vascular Cell Division. Development 2010, 137, 767–774. [Google Scholar] [CrossRef] [Green Version]
- Hirakawa, Y.; Kondo, Y.; Fukuda, H. Regulation of Vascular Development by CLE Peptide-Receptor Systems. J. Integr. Plant Biol. 2010, 52, 8–16. [Google Scholar] [CrossRef]
- Hirakawa, Y.; Kondo, Y.; Fukuda, H. TDIF Peptide Signaling Regulates Vascular Stem Cell Proliferation via the WOX4 Homeobox Gene in Arabidopsis. Plant Cell 2010, 22, 2618–2629. [Google Scholar] [CrossRef] [Green Version]
- Betsuyaku, S.; Takahashi, F.; Kinoshita, A.; Miwa, H.; Shinozaki, K.; Fukuda, H.; Sawa, S. Mitogen-Activated Protein Kinase Regulated by the CLAVATA Receptors Contributes to Shoot Apical Meristem Homeostasis. Plant Cell Physiol. 2011, 52, 14–29. [Google Scholar] [CrossRef] [Green Version]
- Katsir, L.; Davies, K.A.; Bergmann, D.C.; Laux, T. Peptide Signaling in Plant Development. Curr. Biol. 2011, 21, R356–R364. [Google Scholar] [CrossRef] [Green Version]
- Etchells, J.P.; Provost, C.M.; Turner, S.R. Plant Vascular Cell Division Is Maintained by an Interaction between PXY and Ethylene Signalling. PLoS Genet. 2012, 8, e1002997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etchells, J.P.; Mishra, L.S.; Kumar, M.; Campbell, L.; Turner, S.R. Wood Formation in Trees Is Increased by Manipulating Pxy-Regulated Cell Division. Curr. Biol. 2015, 25, 1050–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dodueva, I.E.; Yurlova, E.V.; Osipova, M.A.; Lutova, L.A. CLE Peptides Are Universal Regulators of Meristem Development. Russ. J. Plant Physiol. 2012, 59, 14–27. [Google Scholar] [CrossRef]
- Dodueva, I.E.; Gancheva, M.S.; Osipova, M.A.; Tvorogova, V.E.; Lutova, L.A. Lateral Meristems of Higher Plants: Phytohormonal and Genetic Control. Russ. J. Plant Physiol. 2014, 61, 571–589. [Google Scholar] [CrossRef]
- Ito, Y.; Nakanomyo, I.; Motose, H.; Iwamoto, K.; Sawa, S.; Dohmae, N.; Fukuda, H. Dodeca-CLE Peptides as Suppressors of Plant Stem Cell Differentiation. Science 2006, 313, 842–845. [Google Scholar] [CrossRef] [PubMed]
- Hirakawa, Y.; Shinohara, H.; Kondo, Y.; Inoue, A.; Nakanomyo, I.; Ogawa, M.; Sawa, S.; Ohashi-Ito, K.; Matsubayashi, Y.; Fukuda, H. Non-Cell-Autonomous Control of Vascular Stem Cell Fate by a CLE Peptide/Receptor System. Proc. Natl. Acad. Sci. USA 2008, 105, 15208–15213. [Google Scholar] [CrossRef] [Green Version]
- Lehesranta, S.J.; Lichtenberger, R.; Helariutta, Y. Cell-to-Cell Communication in Vascular Morphogenesis. Curr. Opin. Plant Biol. 2010, 13, 59–65. [Google Scholar] [CrossRef]
- Schrader, J.; Moyle, R.; Bhalerao, R.; Hertzberg, M.; Lundeberg, J.; Nilsson, P.; Bhalerao, R.P. Cambial Meristem Dormancy in Trees Involves Extensive Remodelling of the Transcriptome. Plant J. 2004, 40, 173–187. [Google Scholar] [CrossRef]
- Suer, S.; Agusti, J.; Sanchez, P.; Schwarz, M.; Greb, T. WOX4 Imparts Auxin Responsiveness to Cambium Cells in Arabidopsis. Plant Cell 2011, 23, 3247–3259. [Google Scholar] [CrossRef] [Green Version]
- Ye, Z.-H.; Zhong, R. Molecular Control of Wood Formation in Trees. J. Exp. Bot. 2015, 66, 4119–4131. [Google Scholar] [CrossRef] [Green Version]
- Fisher, K.; Turner, S. PXY, a Receptor-like Kinase Essential for Maintaining Polarity during Plant Vascular-Tissue Development. Curr. Biol. 2007, 17, 1061–1066. [Google Scholar] [CrossRef] [PubMed]
- Kucukoglu, M.; Nilsson, J.; Zheng, B.; Chaabouni, S.; Nilsson, O. WUSCHEL-RELATED HOMEOBOX4 (WOX4)-like Genes Regulate Cambial Cell Division Activity and Secondary Growth in Populus Trees. New Phytol. 2017, 215, 642–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novitskaya, L.; Nikolaeva, N.; Tarelkina, T. Endogenous Variability of the Figured Wood of Karelian Birch. Wulfenia 2016, 23, 175–188. [Google Scholar]
- Novitskaya, L.L.; Shulyakovskaya, T.A.; Galibina, N.A.; Ilyinova, M.K. Membrane Lipid Composition upon Normal and Patterned Wood Formation in Betula Pendula Roth. J. Plant Growth Regul. 2018, 37, 958–970. [Google Scholar] [CrossRef]
- Novitskaya, L.L.; Tarelkina, T.V.; Galibina, N.A.; Moshchenskaya, Y.L.; Nikolaeva, N.N.; Nikerova, K.M.; Podgornaya, M.N.; Sofronova, I.N.; Semenova, L.I. The Formation of Structural Abnormalities in Karelian Birch Wood Is Associated with Auxin Inactivation and Disrupted Basipetal Auxin Transport. J. Plant Growth Regul. 2020, 39, 378–394. [Google Scholar] [CrossRef]
- Lyubavskaya, A.Y. Karelian Birch; Publishing House of Moscow State Forest University: Moscow, Russia, 2006. [Google Scholar]
- Novitskaya, L.; Nikolaeva, N.; Galibina, N.; Tarelkina, T.; Semenova, L. The Greatest Density of Parenchyma Inclusions in Karelian Birch Wood Occurs at Confluences of Phloem Flows. Silva Fenn. 2016, 50, 1461–1478. [Google Scholar] [CrossRef] [Green Version]
- Tarelkina, T.V.; Novitskaya, L.L.; Galibina, N.A.; Moshchenskaya, Y.L.; Nikerova, K.M.; Nikolaeva, N.N.; Sofronova, I.N.; Ivanova, D.S.; Semenova, L.I. Expression Analysis of Key Auxin Biosynthesis, Transport, and Metabolism Genes of Betula Pendula with Special Emphasis on Figured Wood Formation in Karelian Birch. Plants 2020, 9, 1406. [Google Scholar] [CrossRef]
- Galibina, N.A.; Tarelkina, T.V.; Chirva, O.V.; Moshchenskaya, Y.L.; Nikerova, K.M.; Ivanova, D.S.; Semenova, L.I.; Serkova, A.A.; Novitskaya, L.L. Molecular Genetic Characteristics of Different Scenarios of Xylogenesis on the Example of Two Forms of Silver Birch Differing in the Ratio of Structural Elements in the Xylem. Plants 2021, 10, 1593. [Google Scholar] [CrossRef]
- Alonso-Serra, J.; Safronov, O.; Lim, K.; Fraser-Miller, S.J.; Blokhina, O.B.; Campilho, A.; Chong, S.; Fagerstedt, K.; Haavikko, R.; Helariutta, Y.; et al. Tissue-specific Study across the Stem Reveals the Chemistry and Transcriptome Dynamics of Birch Bark. New Phytol. 2019, 222, 1816–1831. [Google Scholar] [CrossRef]
- Etchells, J.P.; Provost, C.M.; Mishra, L.; Turner, S.R. WOX4 and WOX14 Act Downstream of the PXY Receptor Kinase to Regulate Plant Vascular Proliferation Independently of Any Role in Vascular Organisation. Development 2013, 140, 2224–2234. [Google Scholar] [CrossRef] [Green Version]
- Etchells, J.P.; Smit, M.E.; Gaudinier, A.; Williams, C.J.; Brady, S.M. A Brief History of the TDIF-PXY Signalling Module: Balancing Meristem Identity and Differentiation during Vascular Development. New Phytol. 2016, 209, 474–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brackmann, K.; Qi, J.; Gebert, M.; Jouannet, V.; Schlamp, T.; Grünwald, K.; Wallner, E.-S.; Novikova, D.D.; Levitsky, V.G.; Agustí, J.; et al. Spatial Specificity of Auxin Responses Coordinates Wood Formation. Nat. Commun. 2018, 9, 875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Eswaran, G.; Alonso-Serra, J.; Kucukoglu, M.; Xiang, J.; Yang, W.; Elo, A.; Nieminen, K.; Damén, T.; Joung, J.-G.; et al. Transcriptional Regulatory Framework for Vascular Cambium Development in Arabidopsis Roots. Nat. Plants 2019, 5, 1033–1042. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Chen, Y.; Li, W.; Li, Q.; Lu, M.; Zhou, G.; Chai, G. Vascular Cambium: The Source of Wood Formation. Front. Plant Sci. 2021, 12, 700928. [Google Scholar] [CrossRef]
- Kondo, Y.; Ito, T.; Nakagami, H.; Hirakawa, Y.; Saito, M.; Tamaki, T.; Shirasu, K.; Fukuda, H. Plant GSK3 Proteins Regulate Xylem Cell Differentiation Downstream of TDIF–TDR Signalling. Nat. Commun. 2014, 5, 3504. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Hu, X.; Yang, Y.; He, C.; Hu, J.; Wang, X. Strigolactone Signaling Regulates Cambial Activity through Repression of WOX4 by Transcription Factor BES1. Plant Physiol. 2022, 188, 255–267. [Google Scholar] [CrossRef]
- Han, S.; Cho, H.; Noh, J.; Qi, J.; Jung, H.-J.; Nam, H.; Lee, S.; Hwang, D.; Greb, T.; Hwang, I. BIL1-Mediated MP Phosphorylation Integrates PXY and Cytokinin Signalling in Secondary Growth. Nat. Plants 2018, 4, 605–614. [Google Scholar] [CrossRef]
- Bagdassarian, K.S.; Brown, C.M.; Jones, E.T.; Etchells, P. Connections in the Cambium, Receptors in the Ring. Curr. Opin. Plant Biol. 2020, 57, 96–103. [Google Scholar] [CrossRef]
- Xu, C.; Shen, Y.; He, F.; Fu, X.; Yu, H.; Lu, W.; Li, Y.; Li, C.; Fan, D.; Wang, H.C.; et al. Auxin-mediated Aux/IAA-ARF-HB Signaling Cascade Regulates Secondary Xylem Development in Populus. New Phytol. 2019, 222, 752–767. [Google Scholar] [CrossRef]
- Mollenhauer, H.H. Plastic Embedding Mixtures for Use in Electron Microscopy. Stain Technol. 1964, 39, 111–114. [Google Scholar]
- IAWA List of Microscopic Features for Hardwood Identification. IAWA Bull. 1989, 10, 219–332.
- Scholz, A.; Klepsch, M.; Karimi, Z.; Jansen, S. How to Quantify Conduits in Wood? Front. Plant Sci. 2013, 4, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angyalossy, V.; Pace, M.R.; Evert, R.F.; Marcati, C.R.; Oskolski, A.A.; Terrazas, T.; Kotina, E.; Lens, F.; Mazzoni-Viveiros, S.C.; Angeles, G.; et al. IAWA List of Microscopic Bark Features. IAWA J. 2016, 37, 517–615. [Google Scholar] [CrossRef] [Green Version]
- Salojärvi, J.; Smolander, O.-P.; Nieminen, K.; Rajaraman, S.; Safronov, O.; Safdari, P.; Lamminmäki, A.; Immanen, J.; Lan, T.; Tanskanen, J.; et al. Genome Sequencing and Population Genomic Analyses Provide Insights into the Adaptive Landscape of Silver Birch. Nat. Genet. 2017, 49, 904–912. [Google Scholar] [CrossRef] [PubMed]
- Marchler-Bauer, A.; Bryant, S.H. CD-Search: Protein Domain Annotations on the Fly. Nucleic Acids Res. 2004, 32, W327–W331. [Google Scholar] [CrossRef] [PubMed]
- Thumuluri, V.; Almagro Armenteros, J.J.; Johansen, A.R.; Nielsen, H.; Winther, O. DeepLoc 2.0: Multi-Label Subcellular Localization Prediction Using Protein Language Models. Nucleic Acids Res. 2022, gkac278. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The Neighbor-Joining Method: A New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: Oxford, UK; New York, NY, USA, 2000; ISBN 978-0-19-513584-8. [Google Scholar]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An Upgraded Gene Feature Visualization Server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Zang, B.; Yao, H.S.; Huang, M.R. Isolation of High Quality RNA and Molecular Manipulations with Various Tissues of Populus. Russ. J. Plant Physiol. 2009, 56, 716–719. [Google Scholar] [CrossRef]
- Moshchenskaya, Y.L.; Galibina, N.A.; Tarelkina, T.V.; Nikerova, K.M.; Chirva, O.V.; Novitskaya, L.L. Selection of Reference Genes for Normalization of Quantitative PCR Data in Real Time in Two Forms of Silver Birch. Russ. J. Plant Physiol. 2021, 68, 214–221. [Google Scholar] [CrossRef]
- Chang, E.; Shi, S.; Liu, J.; Cheng, T.; Xue, L.; Yang, X.; Yang, W.; Lan, Q.; Jiang, Z. Selection of Reference Genes for Quantitative Gene Expression Studies in Platycladus Orientalis (Cupressaceae) Using Real-Time PCR. PLoS ONE 2012, 7, e33278. [Google Scholar] [CrossRef] [PubMed] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenotype | Number of Cells of Cambial Zone | Number of Differentiating Phloem Cells | Number of Sieve Tubes | |||||
|---|---|---|---|---|---|---|---|---|
| 11 June | 25 June | 28 May | 11 June | 25 June | 28 May | 11 June | 25 June | |
| Bp trees | 8.6 ± 0.3 | 7.9 ± 0.5 | 2.9 ± 0.1 | 2.9 ± 0.1 | 3.5 ± 0.1 | 3.8 ± 0.1 | 4.9 ± 0.1 | 4.9 ± 0.1 |
| BcNFt trees | 8.8 ± 0.3 | 6.3 ± 0.3 | 2.3 ± 0.2 | 3.0 ± 0.1 | 3.4 ± 0.1 | 3.6 ± 0.2 | 5.2 ± 0.2 | 5.2 ± 0.2 |
| Bc FT trees | 8.3 ± 0.3 | 8.3 ± 0.4 | 2.2 ± 0.2 | 3.3 ± 0.1 | 3.6 ± 0.2 | 3.2 ± 0.3 | 4.3 ± 0.2 | 3.1 ± 0.2 |
| Gene Name | Gene ID | Number of Amino Acid Residues in Prepropeptide | CLE-Motif Sequence | Subcellular Localization (Likelihood) |
|---|---|---|---|---|
| BpCLE41/44a | Bpev01.c0016.g0065 | 104 | HEVPSGPNPISN | Extracellular (0.8902) |
| BpCLE41/44b | Bpev01.c0443.g0012 | 117 | HEVPSGPNPISN | Extracellular (0.8565) |
| Gene Name | Gene ID | Number of Amino Acid Residues in Peptide | Location of Transmembrane Helices | Location of Protein Kinase Domain | Subcellular Localization (Likelihood) |
|---|---|---|---|---|---|
| BpPXL2 | Bpev01.c0000.g0018 | 1020 | 642–664 | 714–985 | Cell membrane (0.9555) |
| BpBAM1/2 | Bpev01.c0163.g0042 | 1022 | 638–657 | 696–1006 | Cell membrane (0.9967) |
| BpBAM3a | Bpev01.c0344.g0029 | 997 | 628–650 | 688–993 | Cell membrane (0.9942) |
| BpBAM3b | Bpev01.c0443.g0028 | 989 | 656–675 | 710–907 | Cell membrane (0.9888) |
| BpPXY | Bpev01.c0668.g0016 | 1028 | 654–676 | 720–987 | Cell membrane (0.9959) |
| BpPXL1 | Bpev01.c1696.g0001 | 1000 | 627–649 | 699–969 | Cell membrane (0.9977) |
| BpCLV1 | Bpev01.c2361.g0013 | 986 | 647–666 | 704–971 | Cell membrane (0.9936) |
| Gene Name | Gene ID | Number of Amino Acid Residues in Peptide | Location of Homeobox Domain | Subcellular Localization (Likelihood) |
|---|---|---|---|---|
| BpWUS | Bpev01.c0022.g0024 | 274 | 25–85 | Nucleus (0.9974) |
| BpWOX13 | Bpev01.c0022.g0148 | 299 | 121–179 | Nucleus (0.9933) |
| BpWOX2 | Bpev01.c0030.g0032 | 177 | 19–52 | Nucleus (0.9183) |
| BpWOX1c | Bpev01.c0130.g0014 | 240 | 31–66 | Nucleus (0.9899) |
| BpWOX1b | Bpev01.c0130.g0015 | 128 | 60–96 | Nucleus (0.8748) |
| BpWOX11/12 | Bpev01.c0441.g0004 | 261 | 28–80 | Nucleus (0.9984) |
| BpWOX3 | Bpev01.c0447.g0003 | 200 | 8–67 | Nucleus (0.9965) |
| BpWOX4 | Bpev01.c0640.g0004 | 219 | 83–142 | Nucleus (0.9907) |
| BpWOX5 | Bpev01.c0817.g0002 | 168 | 30–88 | Nucleus (0.9987) |
| BpWOX1a | Bpev01.c1525.g0003 | 334 | 65–124 | Nucleus (0.9971) |
| BpWOX8/9 | Bpev01.c2517.g0001 | 349 | 46–106 | Nucleus (0.999) |
| Gene Name | Gene ID | Forward Primer (5′→3′) | Reverse Primer (5′→3′) | Ta, °C |
|---|---|---|---|---|
| Ef1a | Bpev01.c0437.g0013 | TCCTTGAGGCTCTTGACTTG | ATACCAGGCTTGATGACACC | 54 |
| GAPDH | Bpev01.c1040.g0016 | AGAATACAAGCCAGAACTCAAC | CTCTACCACCTCTCCAATCC | 54 |
| BpCLE41/44a | Bpev01.c0016.g0065 | TGCTCCTCTTGCTTGTTACTC | GCCGATTGTTGTGTTGAAGG | 55 |
| BpCLE41/44b | Bpev01.c0443.g0012 | TGGCGGCGTTGGCAAGTCC | TGGCAATGGCGGTTTACCTCTCC | 60 |
| BpPXY | Bpev01.c0668.g0016 | GTCCTATACCGCCGAGATATG | GCACGCTACCTTCCAACG | 55 |
| BpWOX4 | Bpev01.c0640.g0004 | ACACCACCGACACTTCTTC | CCTTATACAGCATCTCCAATATCC | 55 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galibina, N.A.; Moshchenskaya, Y.L.; Tarelkina, T.V.; Chirva, O.V.; Nikerova, K.M.; Serkova, A.A.; Semenova, L.I.; Ivanova, D.S. Changes in the Activity of the CLE41/PXY/WOX Signaling Pathway in the Birch Cambial Zone under Different Xylogenesis Patterns. Plants 2022, 11, 1727. https://doi.org/10.3390/plants11131727
Galibina NA, Moshchenskaya YL, Tarelkina TV, Chirva OV, Nikerova KM, Serkova AA, Semenova LI, Ivanova DS. Changes in the Activity of the CLE41/PXY/WOX Signaling Pathway in the Birch Cambial Zone under Different Xylogenesis Patterns. Plants. 2022; 11(13):1727. https://doi.org/10.3390/plants11131727
Chicago/Turabian StyleGalibina, Natalia A., Yulia L. Moshchenskaya, Tatiana V. Tarelkina, Olga V. Chirva, Kseniya M. Nikerova, Aleksandra A. Serkova, Ludmila I. Semenova, and Diana S. Ivanova. 2022. "Changes in the Activity of the CLE41/PXY/WOX Signaling Pathway in the Birch Cambial Zone under Different Xylogenesis Patterns" Plants 11, no. 13: 1727. https://doi.org/10.3390/plants11131727
APA StyleGalibina, N. A., Moshchenskaya, Y. L., Tarelkina, T. V., Chirva, O. V., Nikerova, K. M., Serkova, A. A., Semenova, L. I., & Ivanova, D. S. (2022). Changes in the Activity of the CLE41/PXY/WOX Signaling Pathway in the Birch Cambial Zone under Different Xylogenesis Patterns. Plants, 11(13), 1727. https://doi.org/10.3390/plants11131727