
Investigation of Morinda citrifolia Activities through Pinoresinol and α-EG Related Gene Expression



Abstract
:1. Introduction
2. Results
2.1. Macronutritional Components
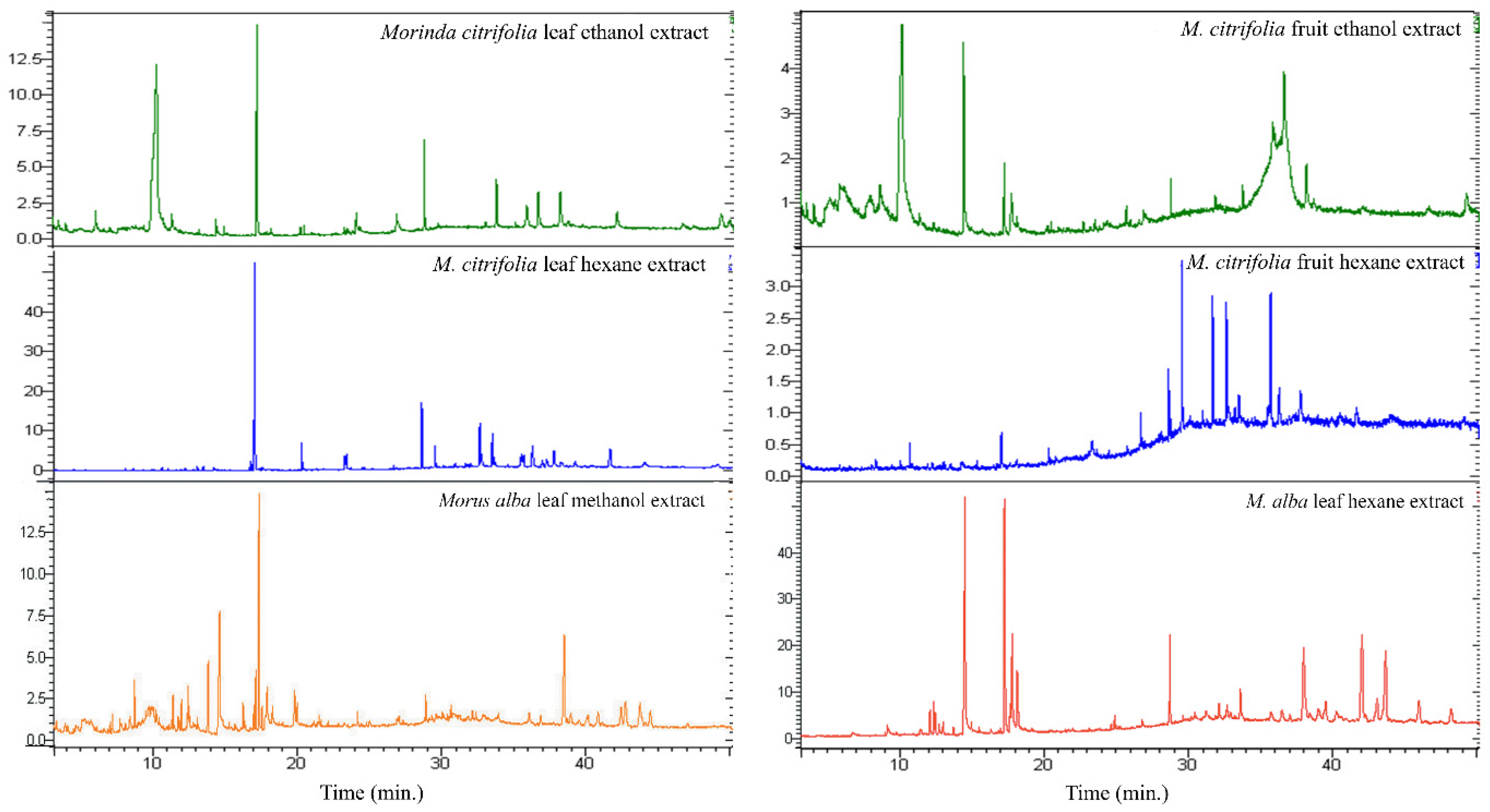
2.2. Phytochemical Constituent Evaluation by Gas Chromatography-Mass Spectrometry
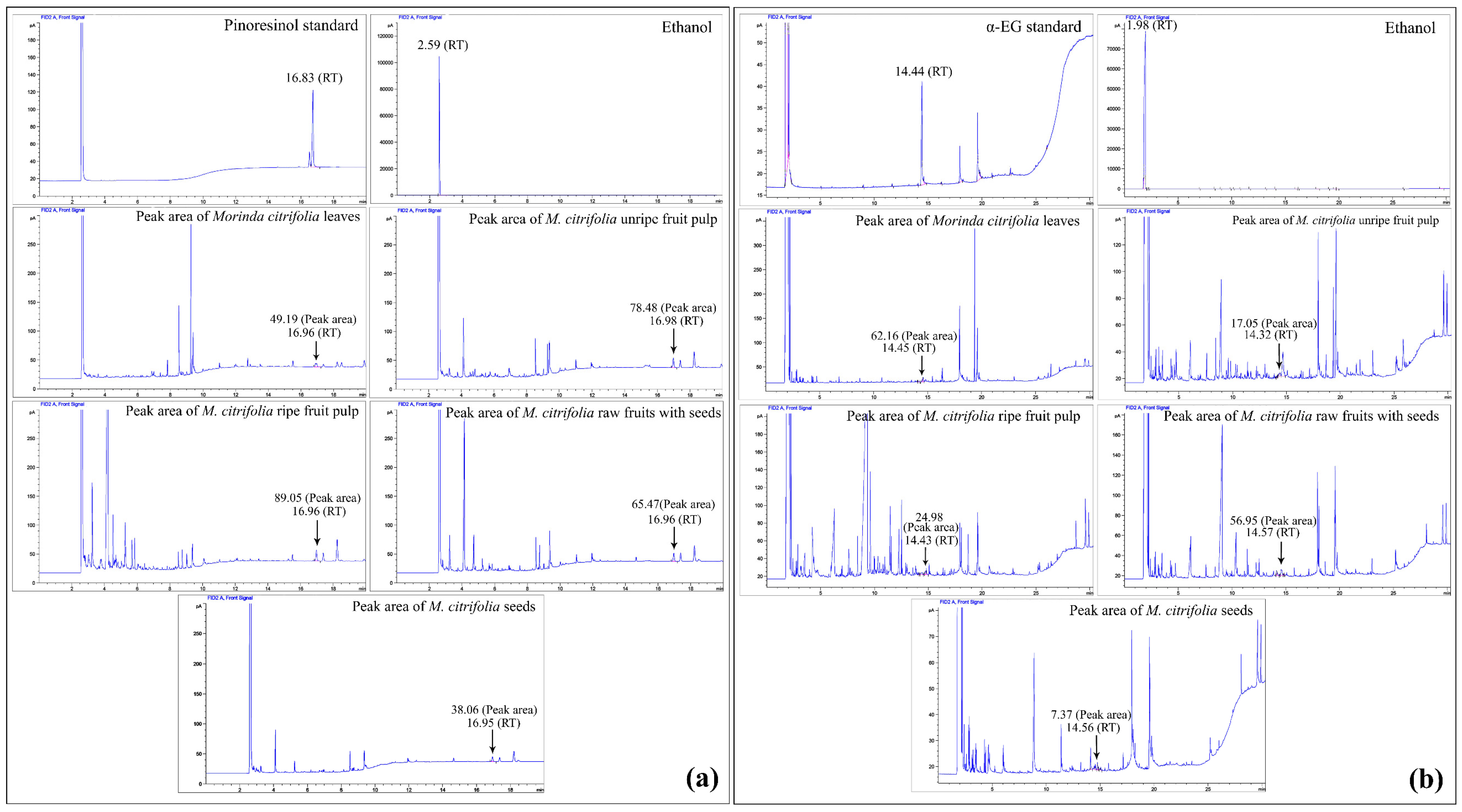
2.3. Gas Chromatography (GC) Compared to the Pinoresinol and α-EG Standards
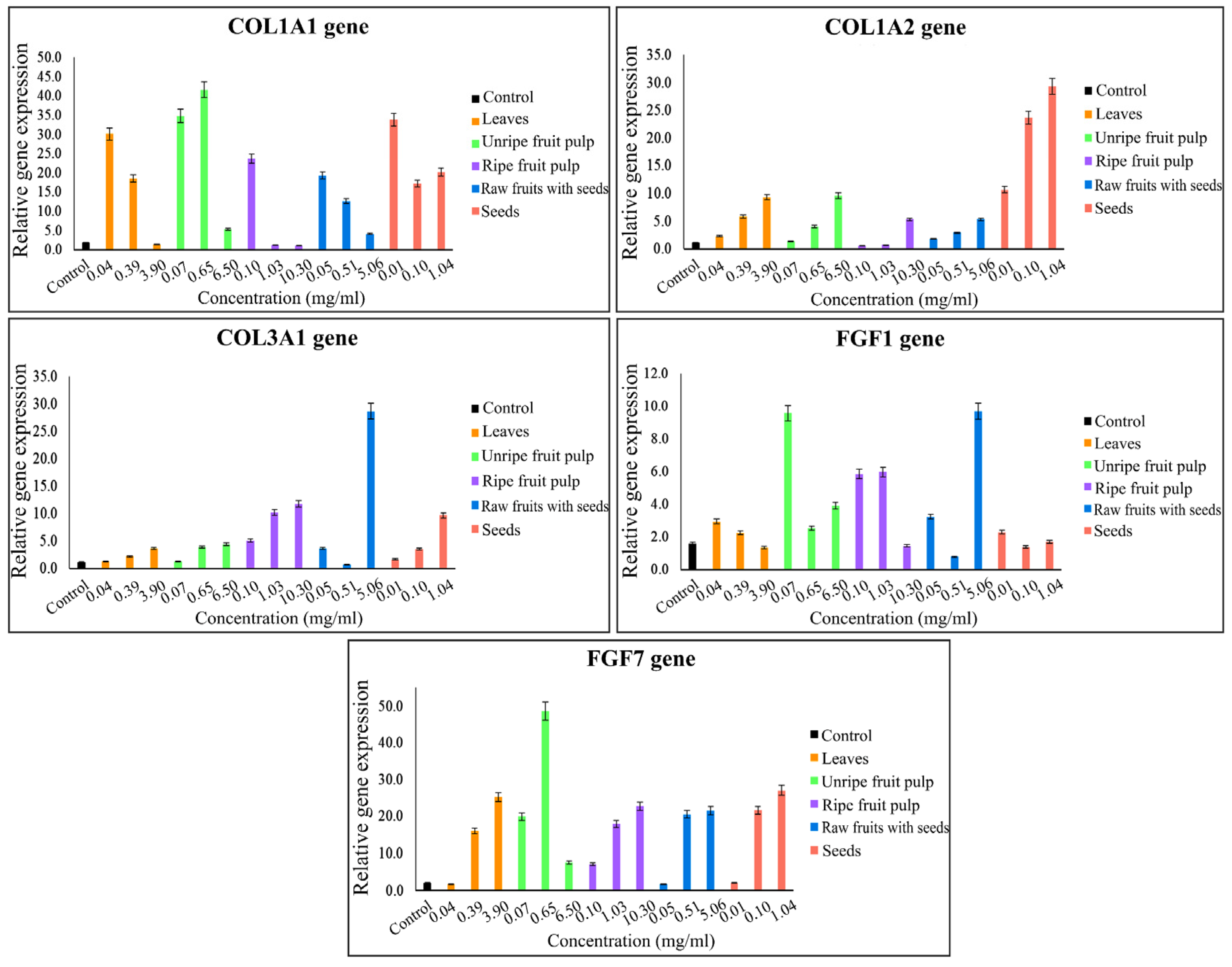
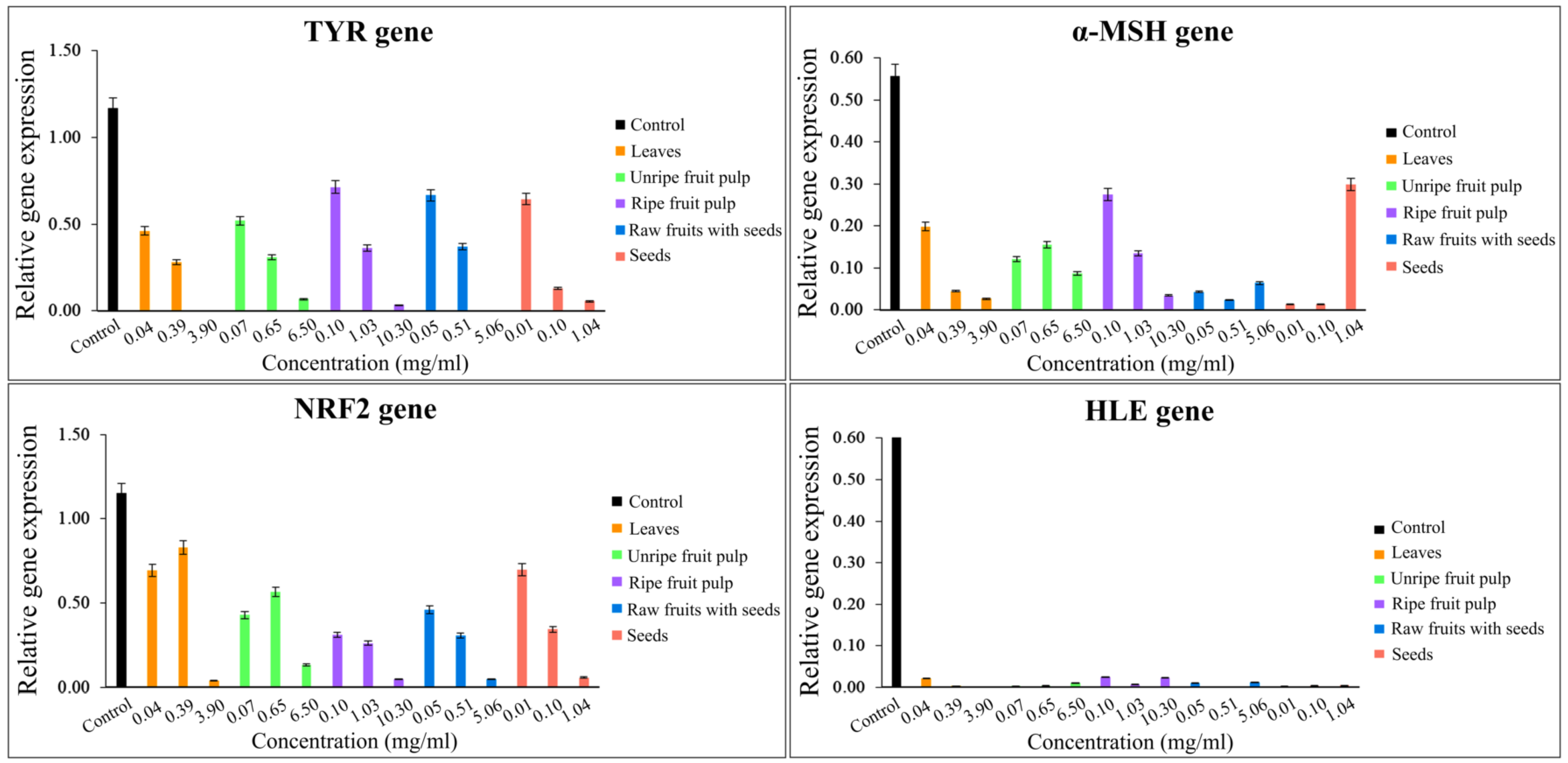
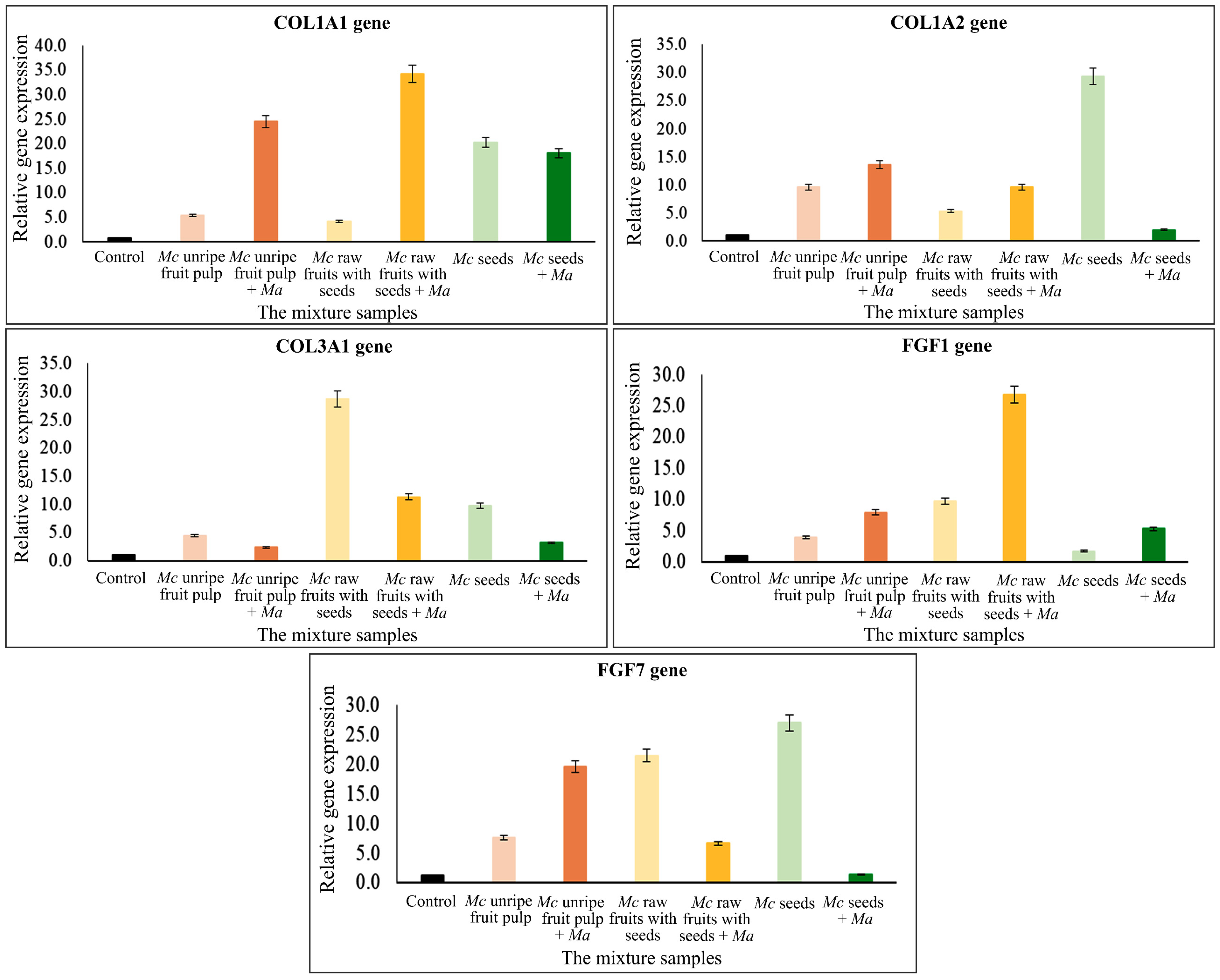
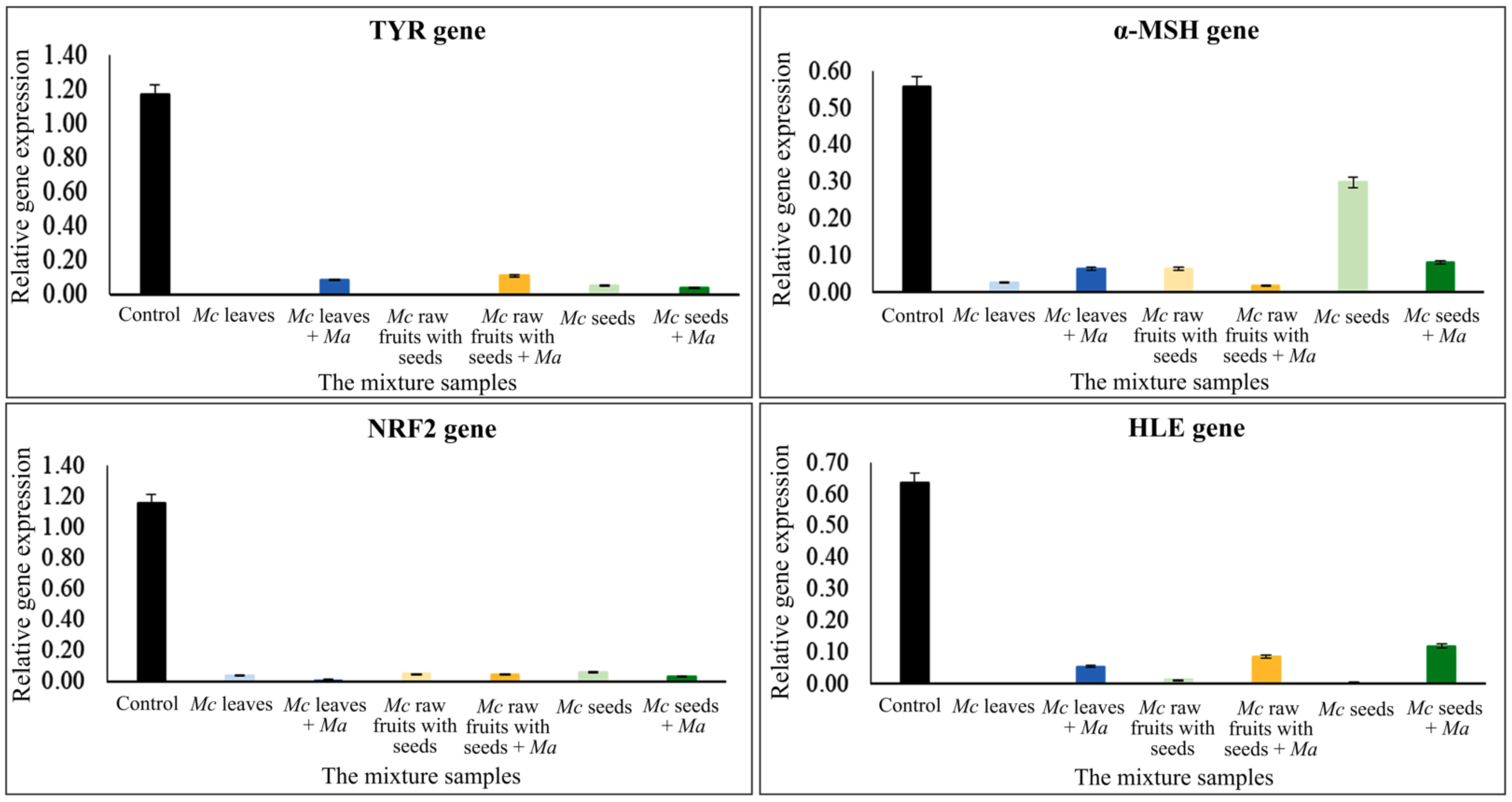
2.4. Gene Expression Quantification
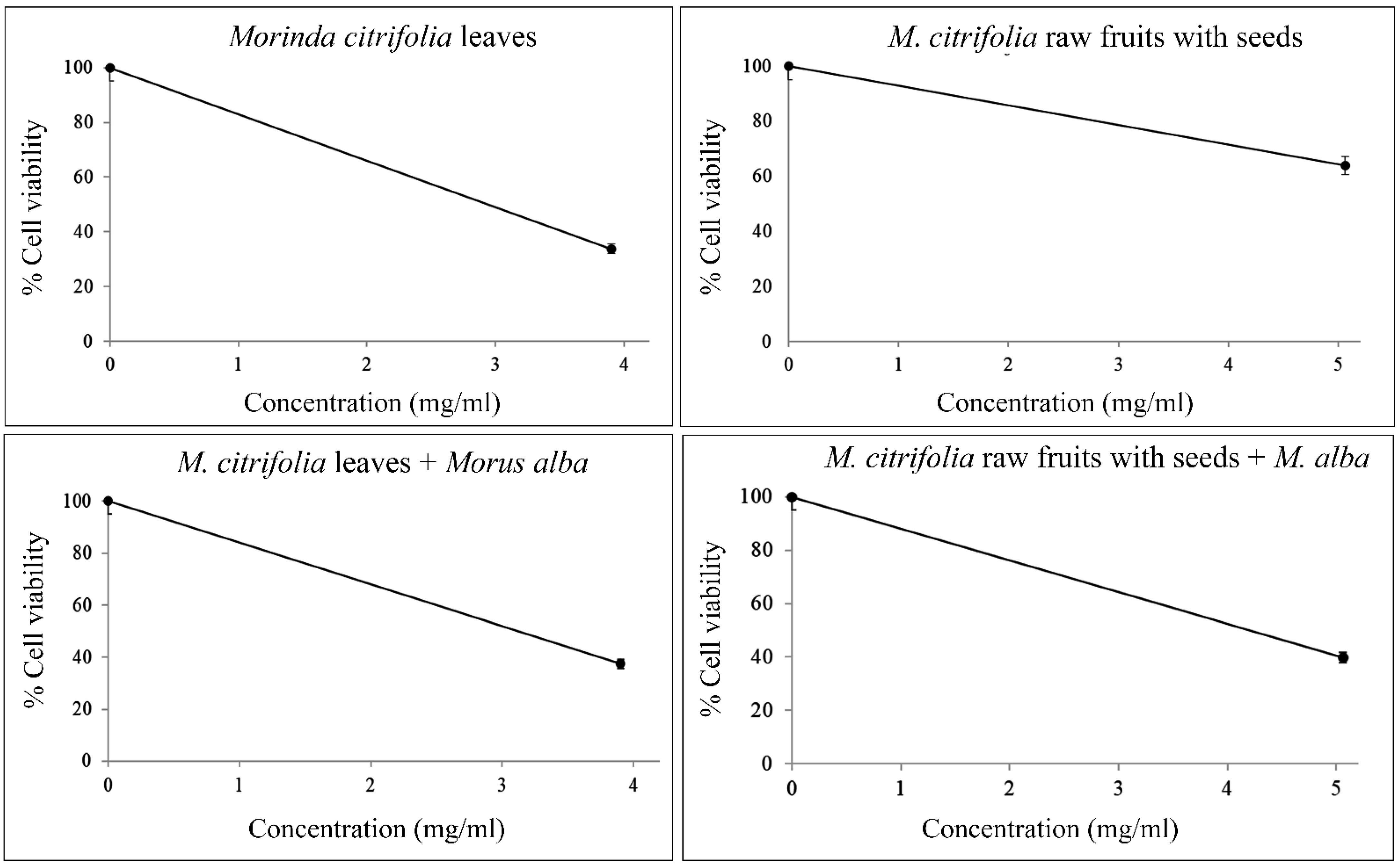
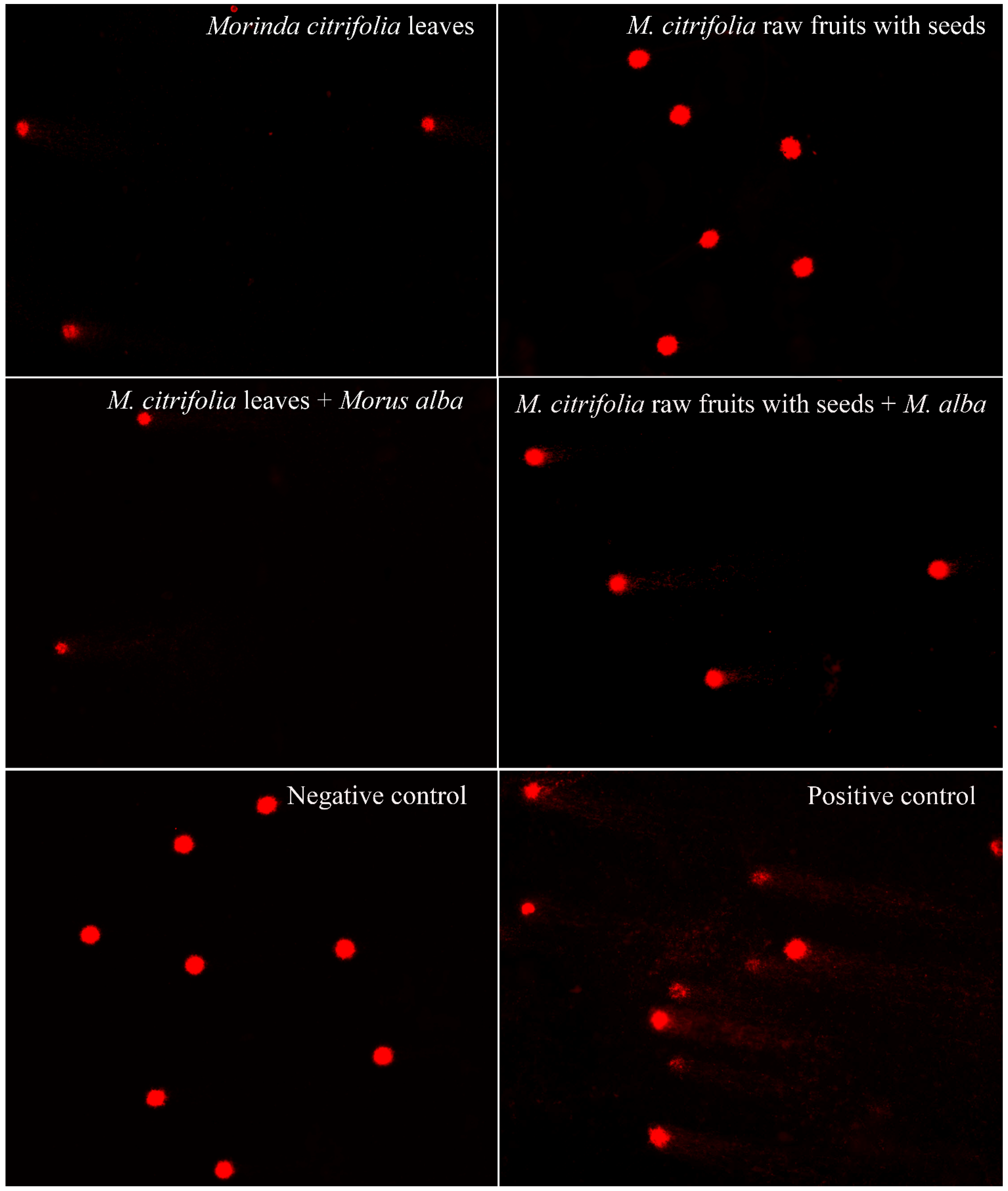
2.5. Cytotoxicity and Genotoxicity Testing via MTT and Comet Assays
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Extract Preparation
4.2. Proximate Analysis of Nutritional Content
4.3. Gas Chromatography-Mass Spectrometry (GC-MS)
4.4. Ethyl-α-d-Glucoside (α-EG) and 3,3′-Bisdemethylpinoresinol (Pinoresinol) Detection by GC Compared to the Standard
4.5. Gene Expression Analysis by qRT-PCR
4.5.1. Extract Preparation
4.5.2. Gene Expression Quantification
- Cell activation
- 2.
- Gene stimulation and inhibition
- 3.
- Protein plant extracts affect gene expression
4.6. Cytotoxicity and Genotoxicity Testing via MTT and Comet Assays
- Stock extract preparation. A powder sample was mixed with ethanol at a ratio of 1 g:5 mL and soaked for 72 h. Each solution was filtered through Whatman No.1 filter paper. The solvent of the filtrate was removed with a rotary evaporator (Rotavapor R-210, Buchi, Flawil, Switzerland) at 800–1000 mbar, 15 °C and 600 rpm for 2 h. Then, DMSO was added to the extracts until completely dissolved and maintained as stock extracts at −20 °C until the MTT and comet experiments were conducted.
- Human peripheral blood mononuclear cell (PBMCs) preparation. PBMCs were isolated from sodium heparin anticoagulated venous blood from a blood bank using FicollPaque Plus (GE Healthcare, Chicago, IL, USA). Freshly isolated PBMCs with viabilities of at least 98% were used for the toxicity testing. The cells were suspended at a concentration of 106 cells/mL for MTT assay and 4–6 × 105 cells/mL for comet assay in modified RPMI-1640 medium supplemented with 10% FBS and 1% antibiotic (streptomycin and penicillin).
- MTT assay. The stock extract was serially 10-fold diluted with water for five working concentrations. The prepared cells were seeded in 96-well plates with 125 µL per well. Then, 12.5 µL of the extract working concentrations were added to the corresponding wells and incubated for 24 h in a humidified CO2 incubator (Esco lifescience, Changi, Singapore) at 37 °C and 5% CO2. Corresponding DMSO concentrations were similarly prepared as vehicle controls. Untreated cells and UV-treated cells were the negative and positive controls, respectively. After this, the plates were centrifuged at 1500 rpm for 10 min and the medium was removed. MTT (Sigma, St. Louis, MO, USA) was added to a final concentration of 0.5 mg/mL and the plates were wrapped with aluminum foil and incubated for 4 h at 37 °C. Formazan crystals were solubilized by adding 100 µL DMSO to each well. The plates were kept in the dark for 2–4 h. The absorbance was read at 570 nm with a microtiter plate spectrophotometer (Multifunction microplate reader; Molecular devices, San Jose, CA, USA). Wells containing medium and MTT without cells were used as blanks. Each concentration treatment was performed in triplicate. All values were expressed as the mean ± SD. The cellular reduction of MTT formed violet crystal formazan through mitochondrial succinate dehydrogenase activity of the viable cells. The violet crystal formazan was quantified following the methods of Freshney et al. [29]. Percentages of cell viability were calculated using the equation cell viability (%) = average viable treated cells/average viable negative control cells × 100 to reveal the cytotoxicity of the plant extracts. Doses inducing 50% inhibition of cell viability (IC50 value) were determined by plotting a graph of the extract concentration against the cell viability. The IC50 value was used for the LD50 calculation [30] to infer hazardous levels, according to the World Health Organization guidelines [20].
- Comet assay. The concentration at IC50 value or the maximum-treated concentration in the case of no IC50 value was used in the comet assay to assess the genotoxicity of plant extracts, following the method described by Singh et al. [31]. Briefly, 500 µL cells in media were added with 50 µL extracts in a 1.5 mL microtube and incubated for 24 h in a humidified CO2 incubator at 37 °C and 5% CO2. Then, the DNA was checked by electrophoresis. The electrophoresis buffer consisted of 0.3 M NaOH and 1 mM EDTA (pH = 10). Power was supplied at a constant of 3.4 v/cm, with an adjustment to 300 mA, for 25 min. To quantify the level of DNA damage, the extent of DNA migration was defined using the Olive Tail Moment (OTM), which is the relative amount of DNA in the tail of the comet multiplied by the median migration distance. The comets were observed at 200× magnifications and images were obtained using an image analysis system (Zeiss, Jena, Germany) attached to a fluorescence microscope (Nikon, Minato-ku, Japan), equipped with a 560 nm excitation filter, 590 nm barrier filter, and a CCD video camera PCO (Kelheim, Germany). At least 150 cells (50 cells for each of the triplicate slides) were examined for each experiment. The CASP software version 1.2.3 (CASPlab, Wroclaw, Poland) was used to analyze the OTM. The negative and positive controls were untreated cells and UV-treated cells, respectively. All experiments were performed in triplicate. The cultures were scored for the experiment. All values were expressed as the median ± S.D. The nonparametric Mann-Whitney U test was used for statistical analyses of the comet assay results; statistical significance was set at p < 0.05.
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Mc | Morinda citrifolia |
| α-EG | ethyl-α-d-glucoside |
| Pinoresinol | 3,3′-bisdemethylpinoresinol |
| ufp | unripe fruit pulp |
| rfp | ripe fruit pulp |
| rawfs | raw fruits with seeds |
| NHDF | normal human dermal fibroblast |
| qRT-PCR | quantitative reverse transcription-polymerase chain reaction |
| Ma | Morus alba |
| PBMCs | human peripheral blood mononuclear cells |
References
- Masuda, M.; Murata, K.; Fukuhama, A.; Naruto, S.; Fujita, T.; Uwaya, A.; Isami, F.; Matsuda, H. Inhibitory effects of constituents of Morinda citrifolia seeds on elastase and tyrosinase. J. Nat. Med. 2009, 63, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Sanni, D.M.; Fatoki, T.H.; Kolawole, A.O.; Akinmoladun, A.C. Xeronine structure and function: Computational comparative mastery of its mystery. In Silico Pharmacol. 2017, 5, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sousa, B.C.; Miguel, C.B.; Rodrigues, W.F.; Machado, J.R.; Silva, M.V.; Costa, T.A.; Lazo-Chica, J.M.; Degasperi, T.P.; Sales-Campos, H.; Bucek, E.U.; et al. Effects of short-term consumption of Morinda citrifolia (Noni) fruit juice on mice intestine, liver and kidney immune modulation. Food Agri. Immunol. 2017, 28, 528–542. [Google Scholar] [CrossRef] [Green Version]
- Sousa, B.C.; Machado, J.R.; Silva, M.V.; Costa, T.A.; Lazo-Chica, J.E.; Degasperi, T.P.; Junior, V.R.; Sales-Campos, H.; Bucek, E.U.; Oliveira, C.J.F. Morinda citrifolia (Noni) Fruit Juice Reduces Inflammatory Cytokines Expression and Contributes to the Maintenance of Intestinal Mucosal Integrity in DSS Experimental Colitis. Mediat. Inflam. 2017, 2017, 6567432. [Google Scholar]
- Soni, N.R. Novel Treatment of Cancer and its Potential in Obesity: “Xeronine” Found in Morinda Citrifolia (Noni Fruit). J. Nutr. Disord. Ther. 2018, 8, 1. [Google Scholar] [CrossRef]
- Zhang, K.; Meng, J.; Li, X.; Tang, X.; Ma, S.; Lv, Y.; Yang, S. Noni (Morinda citrifolia L.) wine prevents the oxidative stress and obesity in mice induced by high-fat diet. J. Food Biochem. 2020, 44, e13460. [Google Scholar] [CrossRef]
- Yang, X.; Lin, C.; Cai, S.; Li, W.; Tang, J.; Wu, X. Therapeutic effects of noni fruit water extract and polysaccharide on oxidative stress and inflammation in mice under high-fat diet. Food Funct. 2020, 11, 1133–1145. [Google Scholar] [CrossRef]
- Sina, H.; Dramane, G.; Tchekounou, P.; Assogba, M.F.; Chabi-Sika, K.; Boya, B.; Socohou, K.; Adjanohoun, A.; Baba-Moussa, L. Phytochemical composition and in vitro biological activities of Morinda citrifolia fruit juice. Saudi J. Biol. Sci. 2020, 28, 1331–1335. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, H.; Masuda, M.; Murata, K.; Abe, Y.; Uwaya., A. Study of the Anti-hotoaging effect of Noni (Morinda citrifolia). In Melanoma—From Early Detection to Treatment, 1st ed.; Duc, T.H., Ed.; IntechOpen: London, UK, 2013; pp. 629–648. [Google Scholar]
- Assi, R.A.; Darwis, Y.; Abdulbaqi, I.M.; Khan, A.A.; Vuanghao, L.; Laghari, M.H. Morinda citrifolia (Noni): A comprehensive review on its industrial uses, pharmacological activities, and clinical trials. Arab. J. Chem. 2017, 10, 691–707. [Google Scholar] [CrossRef]
- Inada, A.C.; Figueiredo, P.S.; Santos-Eichler, R.A.; Freitas, K.C.; Hiane, P.A.; Castro, A.P.; Guimarães, R.C.A. Morinda citrifolia Linn. (Noni) and Its Potential in Obesity-Related Metabolic Dysfunction. Nutrients 2017, 9, 540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, E.S.; Oliveira, D.; Hotza, D. Properties and Applications of Morinda citrifolia (Noni). Compr. Rev. Food Sci. Food Saf. 2019, 8, 883–909. [Google Scholar] [CrossRef] [Green Version]
- Sabsung, A.; Phornchirasilp, S.; Luanratana, O.; Sirikulchayanonta, V.; Tiengda, C. A toxicity study of Morus alba L. leaf extract. Thai J. Pharmacol. 2004, 26, 7. [Google Scholar]
- Polumackanycz, M.; Wesolowski, M.; Viapiana, A. Morus alba L. and Morus nigra L. leaves as a promising food source of phenolic compounds with antioxidant activity. Plant Food. Hum. Nutr. 2021, 76, 458–465. [Google Scholar] [CrossRef]
- Tominaga, N.; Nishiyama, T.; Hayashi, T. Collagens in skin. Sen-I Gakkaishi 1986, 42, 237–242. [Google Scholar] [CrossRef] [Green Version]
- Drąg-Zalesińska, M.; Rembiałkowska, N.; Borska, S.; Saczko, J.; Drąg, M.; Poręba, M.; Kulbacka, J. A New Betulin Derivative Stimulates the Synthesis of Collagen in Human Fibroblasts Stronger than its Precursor. In Vivo 2019, 33, 1087–1093. [Google Scholar] [CrossRef] [Green Version]
- Wang, H. A Review of the Effects of Collagen Treatment in Clinical Studies. Polymers 2021, 13, 3868. [Google Scholar] [CrossRef] [PubMed]
- Varani, J.; Dame, M.K.; Rittie, L.; Suzanne, E.G.; Fligiel, K.S.; Gary, J.; Fisher Voorhees, J.J. Decreased collagen production in chronologically aged skin: Roles of aged-dependent alteration in fibroblast function and defective mechanical stimulation. Am. J. Pathol. 2006, 168, 1861–1868. [Google Scholar] [CrossRef] [Green Version]
- Ma, Q. Role of Nrf2 in oxidative stress and toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 23, 401–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. The WHO Recommended Classification of Pesticides by Hazard and Guidelines to Classification. 2009. Available online: https://apps.who.int/iris/handle/10665/44271 (accessed on 21 September 2018).
- Horwitz, W. Official Methods of Analysis of AOAC International, 17th ed.; Association of Official Analytical Chemists (AOAC): Gaithersburg, MD, USA, 2000. [Google Scholar]
- Yuan, J.S.; Reed, A.; Feng Chen, F.; Stewart, N.C. Statistical analysis of real-time PCR data. BMC Bioinform. 2006, 7, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogaki, T.; Mitani, K.; Oura, Y.; Ozeki, K. Effects of ethyl-α-d-glucoside on human dermal fibroblasts. Biosci. Biotechnol. Biochem. 2017, 81, 1706–1711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Praetorius, C.; Grill, C.; Stacey, S.N.; Metcalf, A.M.; Gorkin, D.U.; Robinson, K.C.; Van, O.E.; Kim, R.S.; Bergsteinsdottir, K.; Ogmundsdottir, M.H.; et al. A polymorphism in IRF4 affects human pigmentation through a tyrosinase-dependent MITF/TFAP2A pathway. Cell 2013, 155, 1022–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eerola, K.; Virtanen, S.; Vähätalo, L.; Ailanen, L.; Cai, M.; Hruby, V.; Savontaus, M.; Savontaus, E. Hypothalamic γ-melanocyte stimulating hormone gene delivery reduces fat mass in male mice. J. Endocrinol. 2018, 239, 19–31. [Google Scholar] [CrossRef]
- Kokot, A.; Metze, D.; Mouchet, N.; Galibert, M.D.; Schiller, M.; Luger, T.A.; Böhm, M. Alpha-melanocyte-stimulating hormone counteracts the suppressive effect of UVB on Nrf2 and Nrf-dependent gene expression in human skin. Endocrinology 2009, 150, 3197–3206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Talas, U.; Dunlop, J.; Khalaf, S.; Leigh, I.M.; Kelsell, D.P. Human elastase 1: Evidence for expression in the skin and the identification of a frequent frameshift polymorphism. J. Investig. Dermatol. 2000, 114, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Freshney, R.I. Culture of Animal Cells: A Manual of Basic Technique and Specialized Applications, 6th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2010; pp. 1–796. [Google Scholar]
- Walum, E. Acute oral toxicity. Environ. Health Perspect. 1998, 106, 497–503. [Google Scholar]
- Singh, N.P.; McCoy, M.T.; Tice, R.R.; Schneider, E.L. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp. Cell Res. 1988, 175, 184–191. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Name | INCI Name | CAS Number | Chemical Formula | Relative Content (%) | |||||
|---|---|---|---|---|---|---|---|---|---|
| Morinda citrifolia | Morus alba | ||||||||
| Fruits | Leaves | Leaves | |||||||
| Ethanol | Hexane | Ethanol | Hexane | Methanol | Hexane | ||||
| 3-Hydroxy-2,3-dihydromaltol | - | - | C6H8O4 | 0.89 | - | 0.51 | - | - | - |
| Ethyl-α-d-glucopyranoside | - | - | C8H16O6 | 54.12 | - | 52.33 | - | - | - |
| Palmitic acid | Palmitic acid | 57-10-3 | C16H32O2 | 11.82 | - | 1.33 | - | 14.22 | 19.32 |
| Phytol | Phytol | 150-86-7/7541-49-3 | C20H40O | 4.39 | 3.30 | 13.19 | 33.26 | 14.93 | 16.04 |
| Lanosteryl acetate | - | 2671-68-3 | C32H52O2 | - | - | - | - | - | 11.52 |
| Lupeol acetate | - | 1617-68-1 | C32H52O2 | - | - | - | - | - | 10.30 |
| dl-α-Tocopherol | Tocopherol | 1406-66-2 | C29H50O2 | 1.87 | 2.61 | 4.30 | - | - | - |
| Methyl oleanonate | - | - | C31H48O3 | 3.95 | - | 2.27 | - | - | - |
| 2-Methoxy-5-Vinylphenol | - | - | C9H10O2 | - | - | 1.56 | - | - | - |
| Ethyl palmitate | Ethyl palmitate | 628-97-7 | C18H36O2 | - | - | 0.43 | - | - | - |
| Chloropyrilene | - | - | C14H18ClN3S | - | - | 0.43 | - | - | - |
| Glycerol 1,3-dipalmitate | - | - | C35H68O5 | - | - | 0.52 | - | - | - |
| 2-Palmitoylglycerol | - | - | C19H38O4 | - | - | 1.43 | - | - | - |
| Glycerol 2-monooleate | - | - | C21H40O4 | - | - | 1.92 | - | - | - |
| Squalene | Squalene | 111-02-04 | C30H50 | - | - | 4.97 | - | - | - |
| Campesterol | - | - | C28H48O | - | - | 2.90 | 4.02 | - | 1.41 |
| γ-Sitosterol | Sitosterol | 83-46-5 | C29H50O | - | - | 4.22 | 6.24 | 13.80 | 10.32 |
| Methyl linolenate | - | 112-63-0 | C19H32O2 | - | - | - | - | 4.32 | - |
| Linolenic acid | Linolenic acid | 60-33-3 | C18H30O2 | - | - | - | - | 3.45 | - |
| Vitamin E | Tocopherol | 1406-66-2 | C29H50O2 | - | - | 1.80 | - | 2.83 | - |
| Methyl arjunolate | - | - | C31H50O5 | - | - | 0.86 | - | - | - |
| α-Tocopherolquinone | - | - | C29H50O3 | - | - | - | 1.91 | - | - |
| Stigmasterol | - | - | C29H48O | - | - | - | 5.97 | - | 1.39 |
| Benzyl beta-d-glucoside | - | 4304-12-5 | C13H16O7 | - | - | - | - | 4.18 | - |
| Aurantiamide | - | 58115-31-4 | C25H26N2O3 | - | - | - | - | 4.08 | - |
| Octadecanoic acid | - | 57-11-4 | C18H36O2 | - | - | - | - | 1.35 | 2.95 |
| Lupenone | - | 1617-70-5 | C30H48O | - | - | - | - | - | 2.84 |
| Simiarenol | - | 1615-94-7 | C30H50O | - | - | - | - | - | 2.83 |
| ethyl 4-ethoxybenzoate | - | 23676-09-7 | C11H14O3 | - | - | - | - | 2.05 | 0.98 |
| Lanosterol | - | 79-63-0 | C30H50O | - | - | - | - | - | 1.87 |
| Loliolide | - | 5989-02-6 | C11H16O3 | - | - | - | - | 1.79 | - |
| Vomifoliol | - | 23526-45-6 | C13H20O3 | - | - | - | - | 1.68 | - |
| Phytol acetate | - | 10236-16-5 | C22H42O2 | - | - | - | - | - | 1.48 |
| 6,10,14-Trimethylpentadecan-2-one | - | 502-69-2 | C18H36O | - | - | - | - | 1.42 | - |
| Dodecanoic acid | Lauric Acid | 143-07-7 | C12H24O2 | - | - | - | - | - | 1.33 |
| Glycerol-beta-palmitate | - | 23470-00-0 | C19H38O4 | - | - | - | - | 1.11 | - |
| Dotriacontane | - | 544-85-4 | C32H66 | - | - | - | - | - | 0.94 |
| γ-Tocopherol | - | 54-28-4 | C28H48O2 | - | - | - | - | 0.56 | 0.88 |
| 2-Pentadecanone, 6,10,14-trimethyl- | Hexahydrofarnesyl acetone | 502-69-2 | C18H36O | - | - | - | - | - | 0.82 |
| delta-Tocopherol | - | 119-13-1 | C27H46O2 | - | - | - | - | 0.75 | - |
| (2,3-Diphenylcyclopropyl)methyl phenyl sulfoxide, trans- | - | 131758-71-9 | C22H20OS | - | - | - | - | - | 0.63 |
| Unknown | - | - | - | 22.96 | 94.09 | 5.03 | 48.60 | 27.48 | 12.15 |
| Plant Samples | Filtrate Volume (mL) | Peak Area (pA × s) | Concentration (mg/mL) | Amounts (mg/g Sample) | ||||
|---|---|---|---|---|---|---|---|---|
| Pinoresinol | α-EG | Pinoresinol | α-EG | Pinoresinol | α-EG | Pinoresinol | α-EG | |
| Leaves | 4.50 | 10.00 | 49.1908 | 62.1647 | 0.09 | 0.30 | 0.20 | 1.51 |
| Unripe fruit pulp | 5.20 | 31.00 | 78.4832 | 17.0516 | 0.13 | 0.11 | 0.34 | 1.75 |
| Ripe fruit pulp | 5.30 | 34.00 | 89.0466 | 24.9789 | 0.14 | 0.15 | 0.38 | 2.48 |
| Raw fruit with seeds | 5.00 | 27.00 | 65.4722 | 56.9494 | 0.11 | 0.28 | 0.28 | 3.77 |
| Seeds | 5.20 | 26.00 | 38.0631 | 7.3699 | 0.08 | 0.07 | 0.20 | 0.94 |
| Treatment and Control | Conc. (mg/mL) | Human Dermal Fibroblast Cells | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ct Value of GADPH | Gene Expression Value (%) | |||||||||||
| COL1A1 | COL1A2 | COL3A1 | FGF1 | FGF7 | ||||||||
| − | +Ma | − | +Ma | − | +Ma | − | +Ma | − | +Ma | |||
| Control | - | 18.70 | 190 | 70 | 110 | 100 | 120 | 100 | 160 | 90 | 210 | 110 |
| Mc unripe fruit pulp | 6.50, 3.11 | 34.45 | 540 | 2450 | 960 | 1360 | 440 | 240 | 390 | 790 | 750 | 1960 |
| Mc raw fruits with seeds | 5.06, 3.11 | 37.19 | 410 | 3200 | 530 | 960 | 2870 | 1130 | 970 | 2670 | 2150 | 660 |
| Mc seeds | 1.04, 3.11 | 34.88 | 2030 | 1810 | 2930 | 200 | 970 | 320 | 170 | 530 | 2700 | 140 |
| The Mixture Samples at a Ratio 6:4 | Conc. (mg/mL) | Human Dermal Fibroblast Cells | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Ct Value of GADPH | Gene Expression Value (%) | |||||||||
| TYR | α-MSH | NRF2 | HLE | |||||||
| − | +Ma | − | +Ma | − | +Ma | − | +Ma | |||
| Control | - | 21.20 | 116.90 | 53.20 | 55.70 | 106.20 | 115.40 | 109.40 | 10.50 | 66.00 |
| Mc leaves + Ma leaves | 3.90, 3.11 | 36.90 | 0.20 | 8.70 | 2.60 | 6.40 | 3.80 | 1.40 | 0.20 | 5.30 |
| Mc raw fruits with seeds + Ma leaves | 5.06, 3.11 | 37.20 | 0.10 | 11.00 | 6.40 | 1.90 | 4.90 | 4.80 | 1.20 | 8.40 |
| Mc seeds + Ma leaves | 1.04, 3.11 | 35.70 | 5.40 | 3.90 | 29.80 | 8.20 | 5.80 | 3.40 | 0.50 | 11.80 |
| Plant Samples | Working Conc. (mg/mL) | IC50 (mg/mL) | LD50 (mg/kg rat) | % Cell Viability (Mean ± S.D.) | Olive Tail Moment (Median ± S.D.) | p-Values |
|---|---|---|---|---|---|---|
| Negative control | - | - | - | - | 0.0972 ± 0.0897 | - |
| Mc leaves | 3.90 | 3.00 | 2077.25 | 33.82 ± 0.04 | 9.9719 ± 0.6977 | <0.0001 |
| Mc raw fruits with seeds | 5.06 | - | - | 62.17 ± 0.04 | 0.1093 ± 0.0891 | 0.1652 |
| Mc leaves + Ma leaves | 3.90, 3.11 | 3.10 | 2102.74 | 37.44 ± 0.08 | 7.1208 ± 0.5955 | <0.0001 |
| Mc raw fruits with seeds + Ma leaves | 5.06, 3.11 | 4.20 | 2354.23 | 39.74 ± 0.04 | 3.2936 ± 0.1640 | <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sudmoon, R.; Kaewdaungdee, S.; Ameamsri, U.; Tanee, T.; Siripiyasing, P.; Wonok, W.; Chaveerach, A. Investigation of Morinda citrifolia Activities through Pinoresinol and α-EG Related Gene Expression. Plants 2022, 11, 1985. https://doi.org/10.3390/plants11151985
Sudmoon R, Kaewdaungdee S, Ameamsri U, Tanee T, Siripiyasing P, Wonok W, Chaveerach A. Investigation of Morinda citrifolia Activities through Pinoresinol and α-EG Related Gene Expression. Plants. 2022; 11(15):1985. https://doi.org/10.3390/plants11151985
Chicago/Turabian StyleSudmoon, Runglawan, Sanit Kaewdaungdee, Unchaleeporn Ameamsri, Tawatchai Tanee, Pornnarong Siripiyasing, Warin Wonok, and Arunrat Chaveerach. 2022. "Investigation of Morinda citrifolia Activities through Pinoresinol and α-EG Related Gene Expression" Plants 11, no. 15: 1985. https://doi.org/10.3390/plants11151985
APA StyleSudmoon, R., Kaewdaungdee, S., Ameamsri, U., Tanee, T., Siripiyasing, P., Wonok, W., & Chaveerach, A. (2022). Investigation of Morinda citrifolia Activities through Pinoresinol and α-EG Related Gene Expression. Plants, 11(15), 1985. https://doi.org/10.3390/plants11151985