
Quantifying the Variation in the Geometries of the Outer Rims of Corolla Tubes of Vinca major L.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
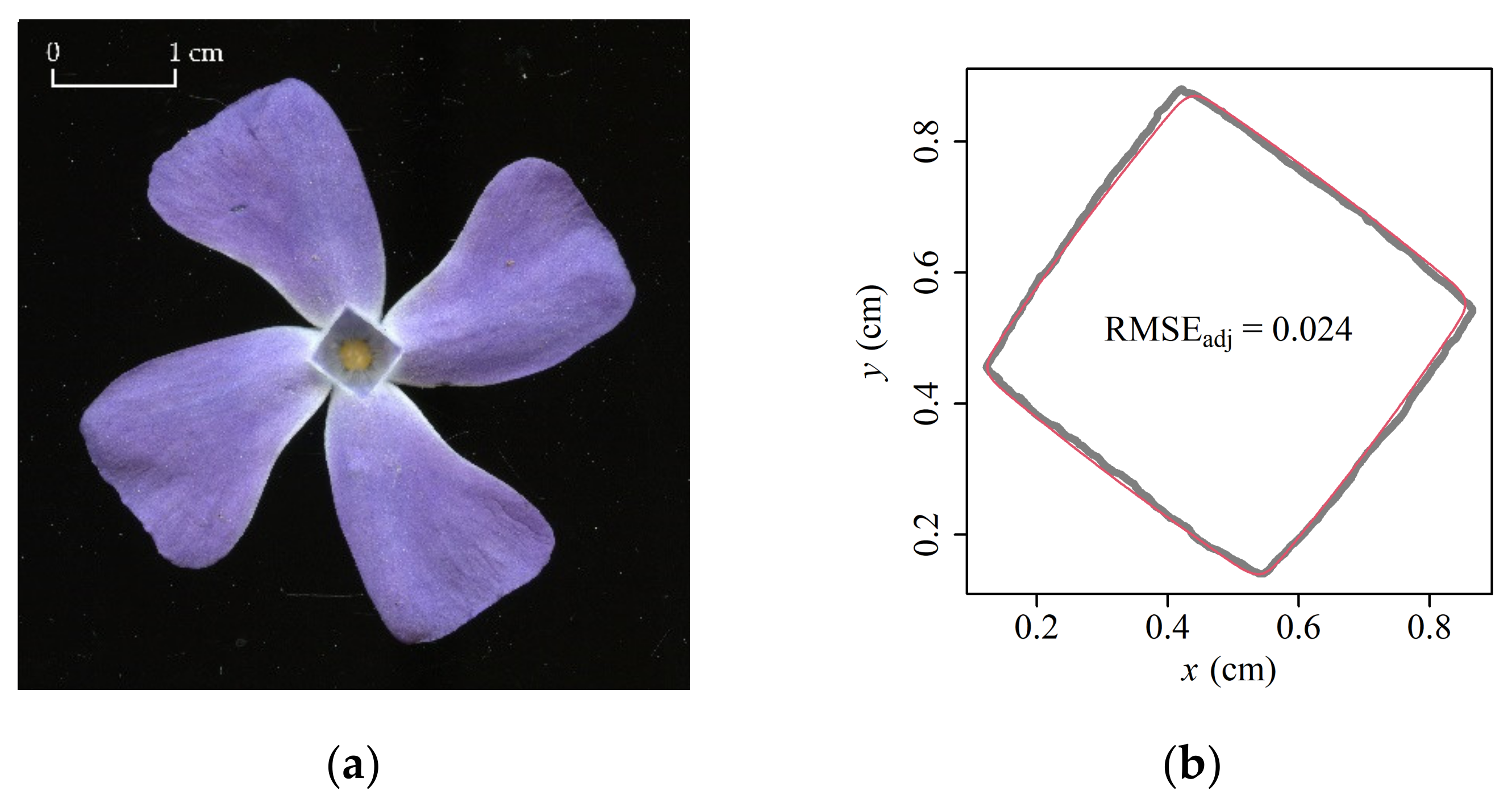
Abstract
:1. Introduction
2. Materials and Methods
2.1. Flower Sampling and Image Processing
2.2. Data Acquisition
2.3. Models
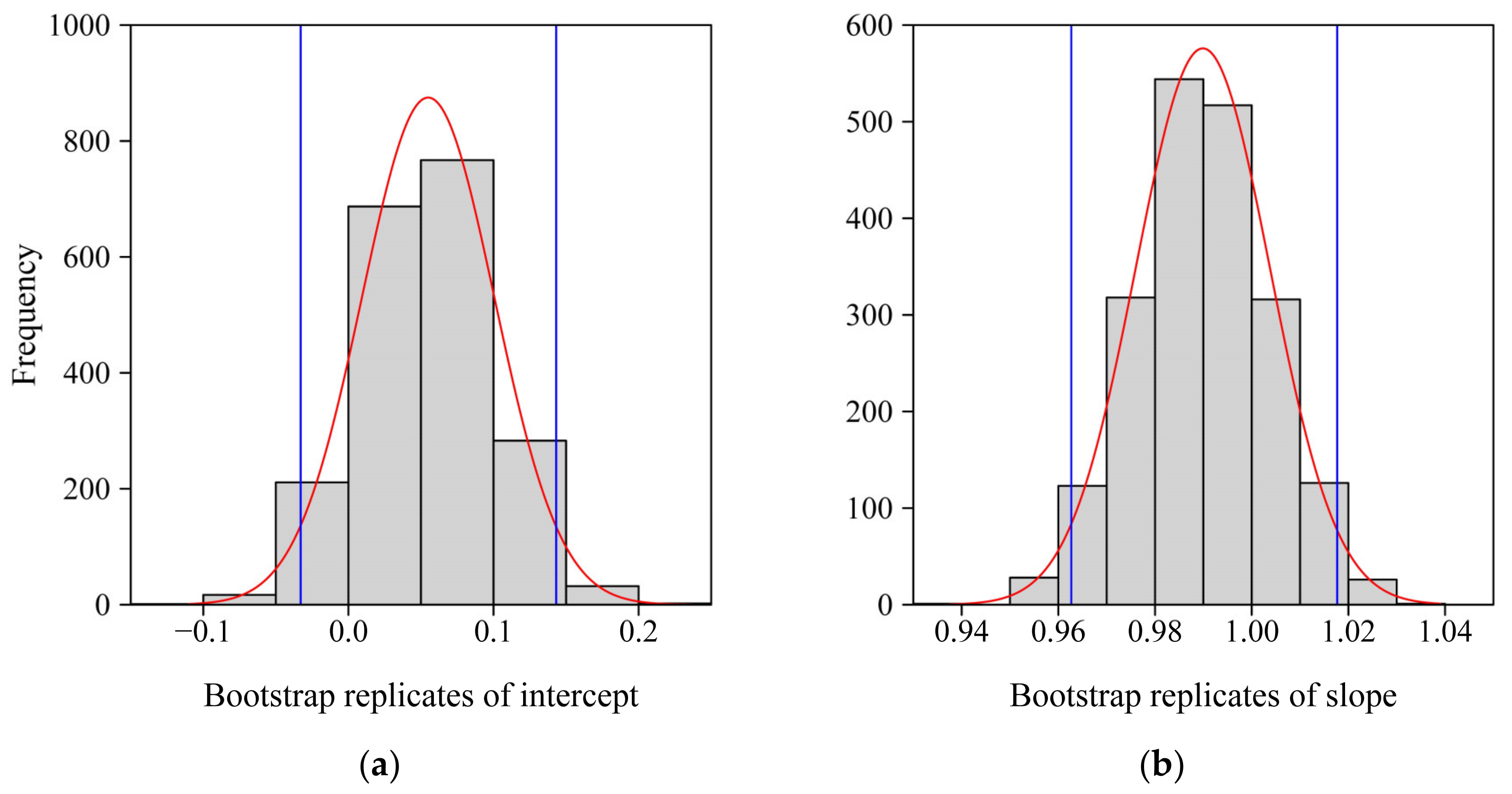
2.4. Model Fitting and Data Analysis
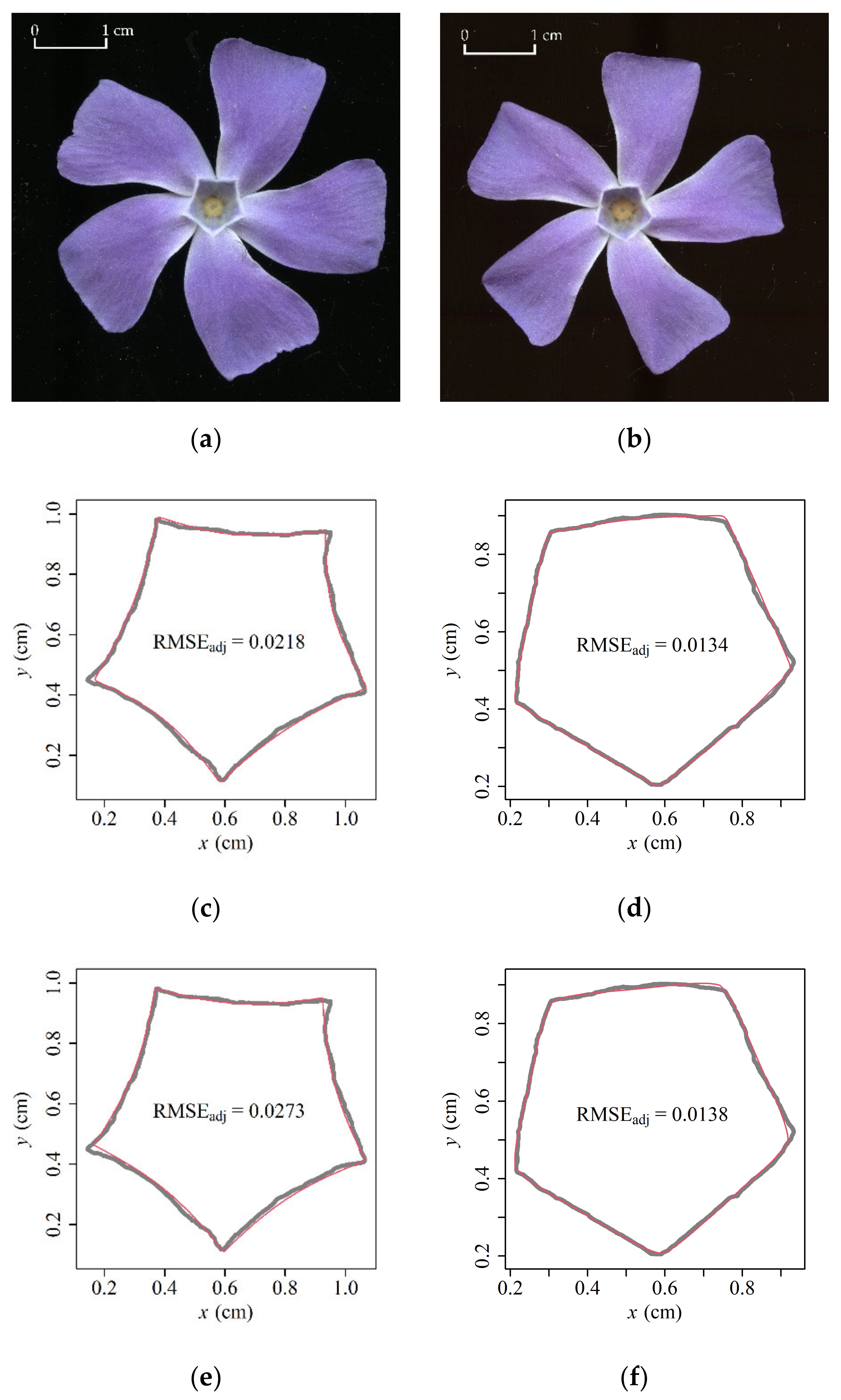
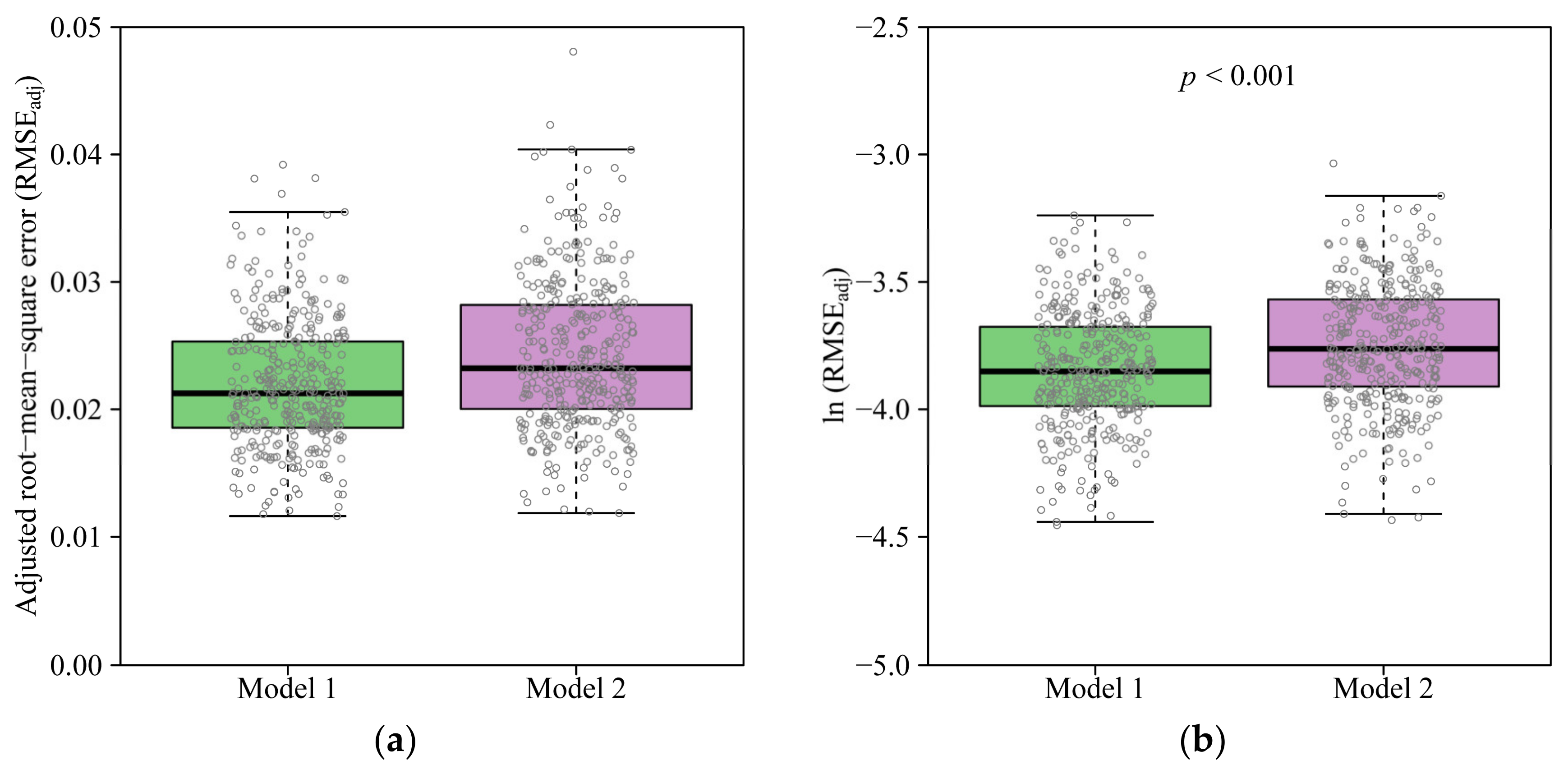
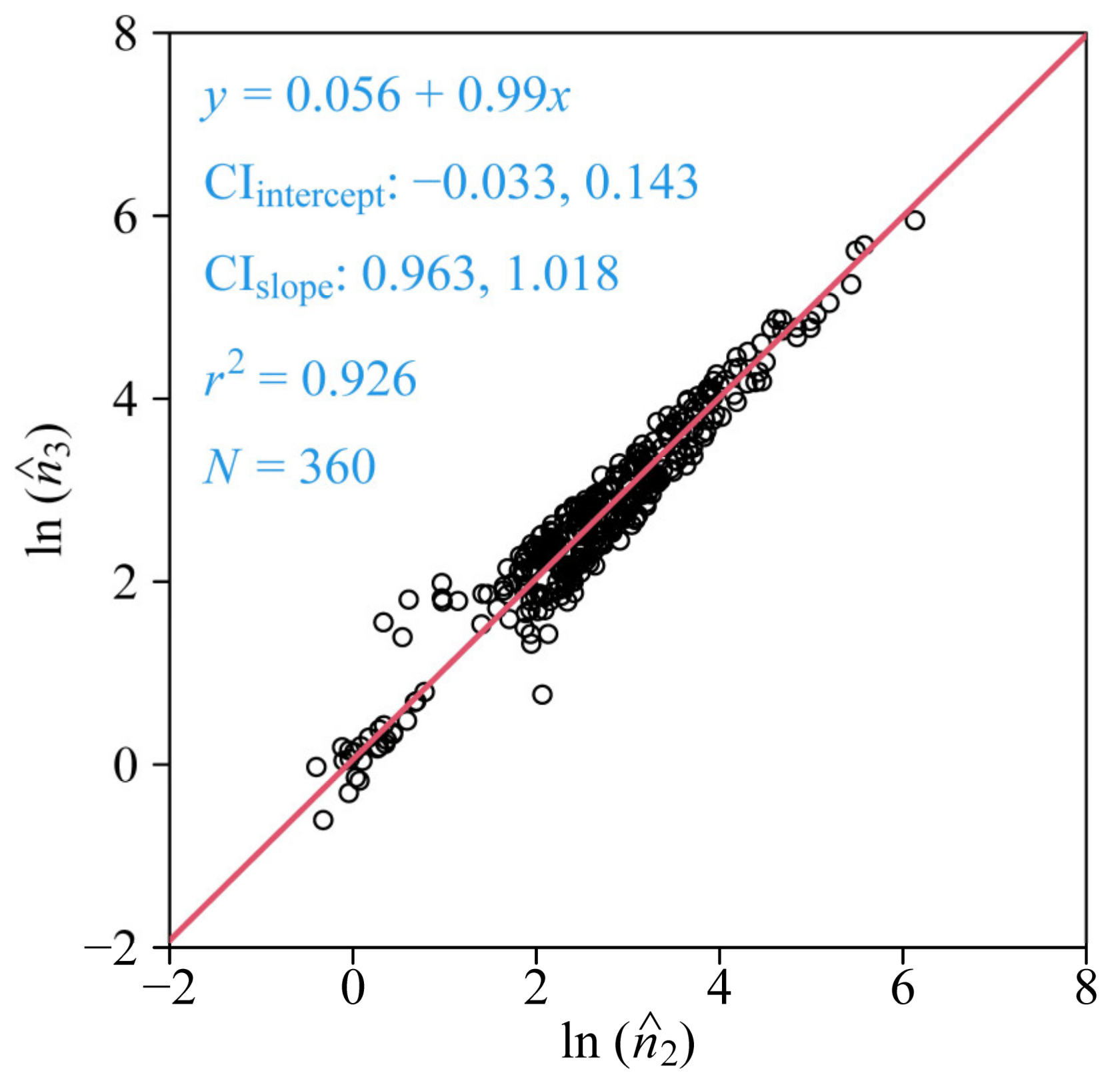
3. Results
4. Discussion
4.1. Analysis of the Prediction Errors
4.2. Comparison of the Two Models
4.3. Variation in the Number of Polygon Sides of the ORCT of V. major
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lamé, G. Examen des Différentes Méthodes Employées Pour Résoudre les Problèmes de Géométrie; V. Courcier: Paris, France, 1818. [Google Scholar]
- Gielis, J. A general geometric transformation that unifies a wide range of natural and abstract shapes. Am. J. Bot. 2003, 90, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Gielis, J. The Geometrical Beauty of Plants; Atlantis Press: Paris, France, 2017. [Google Scholar]
- Shi, P.; Huang, J.; Hui, C.; Grissino-Mayer, H.D.; Tardif, J.C.; Zhai, L.; Wang, F.; Li, B. Capturing spiral radial growth of conifers using the superellipse to model tree-ring geometric shape. Front. Plant Sci. 2015, 6, 856. [Google Scholar] [CrossRef] [Green Version]
- Shi, P.; Xu, Q.; Sandhu, H.S.; Gielis, J.; Ding, Y.; Li, H.; Dong, X. Comparison of dwarf bamboos (Indocalamus sp.) leaf parameters to determine relationship between spatial density of plants and total leaf area per plant. Ecol. Evol. 2015, 5, 4578–4589. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Zhang, L.; Reddy, G.V.P.; Hui, C.; Gielis, J.; Ding, Y.; Shi, P. A geometrical model for testing bilateral symmetry of bamboo leaf with a simplified Gielis equation. Ecol. Evol. 2016, 6, 6798–6806. [Google Scholar] [CrossRef] [PubMed]
- Shi, P.; Ratkowsky, D.A.; Gielis, J. The generalized Gielis geometric equation and its application. Symmetry 2020, 12, 645. [Google Scholar] [CrossRef] [Green Version]
- Tian, F.; Wang, Y.; Sandhu, H.S.; Gielis, J.; Shi, P. Comparison of seed morphology of two ginkgo cultivars. J. For. Res. 2020, 31, 751–758. [Google Scholar] [CrossRef]
- Li, Y.; Quinn, B.K.; Gielis, J.; Li, Y.; Shi, P. Evidence that supertriangles exist in nature from the vertical projections of Koelreuteria paniculata fruit. Symmetry 2022, 14, 23. [Google Scholar] [CrossRef]
- Shi, P.; Gielis, J.; Niklas, K.J. Comparison of a universal (but complex) model for avian egg shape with a simpler model. Ann. N. Y. Acad. Sci. 2022, in press. [CrossRef]
- Weng, Y. Growth and form in biology: Generation of the plant morphology by spontaneous symmetry breaking based on a pressure field. Chin. Phys. Lett. 2004, 21, 211–214. [Google Scholar]
- Gielis, J.; Caratelli, D.; Fougerolle, Y.; Ricci, P.E.; Gerats, T. A biogeometrical model for corolla fusion in asclepiad flowers. In Modeling in Mathematics; Gielis, J., Ricci, P.E., Tavkhelidze, I., Eds.; Atlantis Press: Paris, France, 2017; pp. 83–105. [Google Scholar]
- Spíchal, L. Superelipsa a superformule. Mat.–Fyzika–Inform. 2020, 29, 54–69. [Google Scholar]
- Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Bot. J. Linn. Soc. 2009, 161, 105–121. [CrossRef] [Green Version]
- Endress, M.E.; Liede-Schumann, S.; Meve, U. An updated classification for Apocynaceae. Phytotaxa 2014, 159, 175–194. [Google Scholar] [CrossRef]
- Ronse De Craene, L. Floral Diagrams: An Aid to Understanding Flower Morphology and Evolution; Cambridge University Press: Cambridge, UK, 2022. [Google Scholar]
- Shi, P.; Ratkowsky, D.A.; Li, Y.; Zhang, L.; Lin, S.; Gielis, J. A general leaf-area geometric formula exists for plants—Evidence from the simplified Gielis equation. Forests 2018, 9, 714. [Google Scholar] [CrossRef] [Green Version]
- Su, J.; Niklas, K.J.; Huang, W.; Yu, X.; Yang, Y.; Shi, P. Lamina shape does not correlate with lamina surface area: An analysis based on the simplified Gielis equation. Glob. Ecol. Conserv. 2019, 19, e00666. [Google Scholar] [CrossRef]
- Shi, P.; Gielis, J.; Quinn, B.K.; Niklas, K.J.; Ratkowsky, D.A.; Schrader, J.; Ruan, H.; Wang, L.; Niinemets, Ü. ‘biogeom’: An R package for simulating and fitting natural shapes. Ann. N. Y. Acad. Sci. 2022, in press. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 1 June 2022).
- Nelder, J.A.; Mead, R. A simplex method for function minimization. Comput. J. 1965, 7, 308–313. [Google Scholar] [CrossRef]
- Wei, H.; Li, X.; Huang, H. Leaf shape simulation of castor bean and its application in nondestructive leaf area estimation. Int. J. Agric. Biol. Eng. 2019, 12, 135–140. [Google Scholar] [CrossRef]
- Milla, R.; Reich, P. The scaling of leaf area and mass: The cost of light interception increases with leaf size. Proc. R. Soc. B Biol. Sci. 2007, 274, 2109–2114. [Google Scholar] [CrossRef] [Green Version]
- Niklas, K.; Cobb, E.; Niinemets, Ü.; Reich, P.; Sellin, A.; Shipley, B.; Wright, I. “Diminishing returns” in the scaling of functional leaf traits across and within species groups. Proc. Natl. Acad. Sci. USA 2007, 104, 8891–8896. [Google Scholar] [CrossRef] [Green Version]
- Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press: Cambridge, UK, 2002; pp. 100–104. [Google Scholar]
- Smith, R. Use and misuse of the reduced major axis for line-fitting. Am. J. Phys. Anthropol. 2009, 140, 476–486. [Google Scholar] [CrossRef]
- Efron, B.; Tibshirani, R.J. An Introduction to the Bootstrap; Chapman and Hall/CRC: New York, NY, USA, 1993. [Google Scholar]
- Sandhu, H.S.; Shi, P.; Kuang, X.; Xue, F.; Ge, F. Applications of the bootstrap to insect physiology. Fla. Entomol. 2011, 94, 1036–1041. [Google Scholar] [CrossRef]
- Royston, P. Remark AS R94: A remark on Algorithm AS 181: The W test for normality. Appl. Statist. 1995, 44, 547–551. [Google Scholar] [CrossRef]
- Liao, F.; Xie, Y.; Jiang, H. The effect of water stress on the physiology of Vinca major ‘Variegata’. Appl. Mech. Mater. 2013, 409–410, 782–787. [Google Scholar] [CrossRef]
- Shen, J. Research on the shade tolerance of Vinca major. For. Sci. Technol. 2012, 37, 15–17. [Google Scholar]
- Wang, J.; Wang, J.; Zhou, J.; He, T.; Li, P. Recent progress of artificial lighting technique and effect of light on plant growth. J. Nanjing For. Univer. (Nat. Sci. Ed.) 2020, 44, 215–222. [Google Scholar]
- Zhuang, Y. Comparative study on the water physiological characteristics of Vinca major and Vinca major ‘Variegata’. Anhui For. Sci. Technol. 2013, 39, 19–24. [Google Scholar]
- Niinemets, Ü. Leaf trait plasticity and evolution in different plant functional types. Ann. Plant Rev. 2020, 3, 473–522. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Miao, Q.; Niinemets, Ü.; Gielis, J.; Shi, P. Quantifying the Variation in the Geometries of the Outer Rims of Corolla Tubes of Vinca major L. Plants 2022, 11, 1987. https://doi.org/10.3390/plants11151987
Wang L, Miao Q, Niinemets Ü, Gielis J, Shi P. Quantifying the Variation in the Geometries of the Outer Rims of Corolla Tubes of Vinca major L. Plants. 2022; 11(15):1987. https://doi.org/10.3390/plants11151987
Chicago/Turabian StyleWang, Lin, Qinyue Miao, Ülo Niinemets, Johan Gielis, and Peijian Shi. 2022. "Quantifying the Variation in the Geometries of the Outer Rims of Corolla Tubes of Vinca major L." Plants 11, no. 15: 1987. https://doi.org/10.3390/plants11151987
APA StyleWang, L., Miao, Q., Niinemets, Ü., Gielis, J., & Shi, P. (2022). Quantifying the Variation in the Geometries of the Outer Rims of Corolla Tubes of Vinca major L. Plants, 11(15), 1987. https://doi.org/10.3390/plants11151987