
Countering Triple Negative Breast Cancer via Impeding Wnt/β-Catenin Signaling, a Phytotherapeutic Approach


Abstract
:1. Introduction
2. Natural Derivatives Targeting TNBC via Impeding the Wnt/β-Catenin Pathway
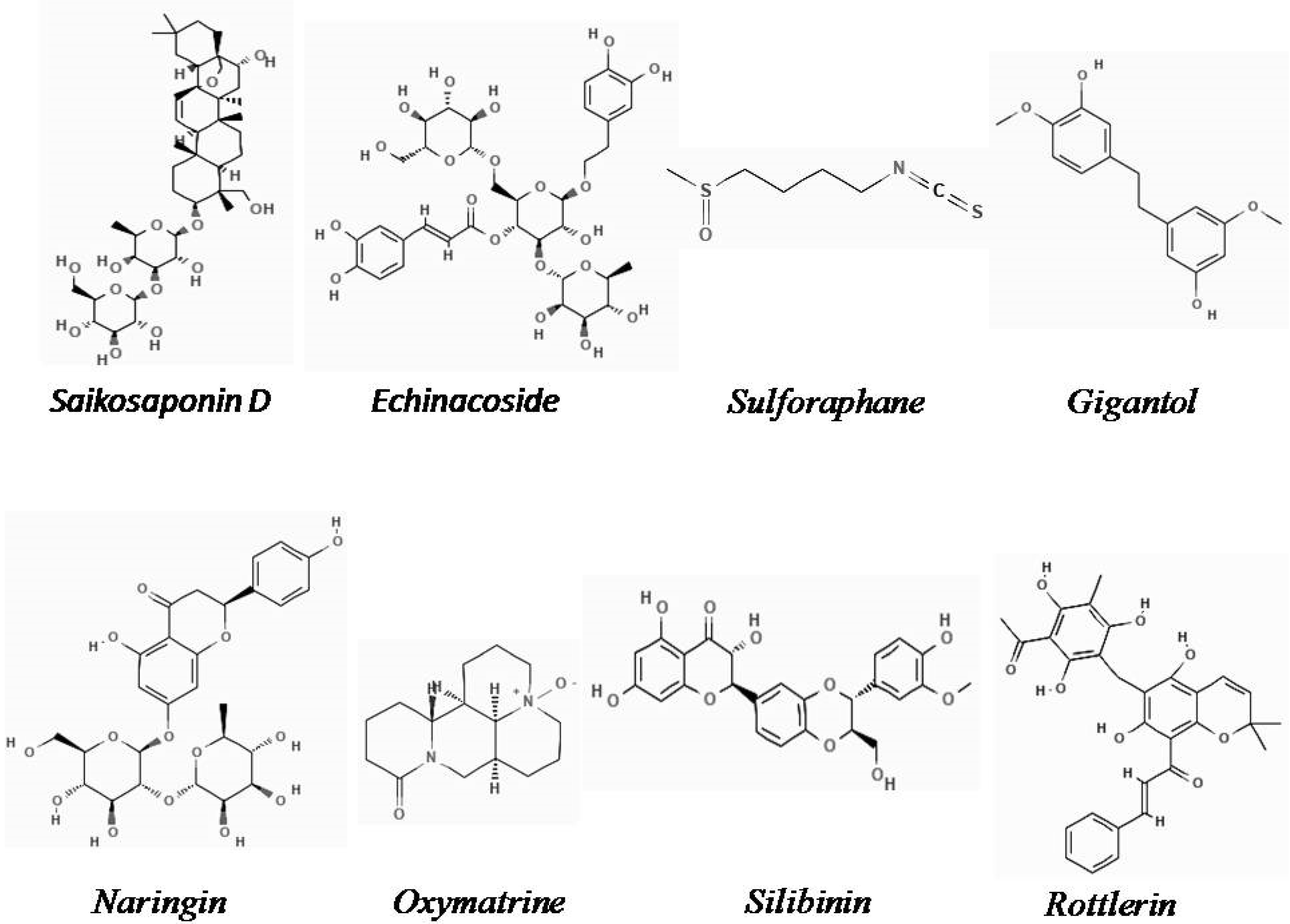
2.1. Saikosaponin D
2.2. Echinacoside
2.3. Sulforaphane
2.4. Gigantol
2.5. Naringin
2.6. Oxymatrine
2.7. Silibinin
2.8. Rottlerin
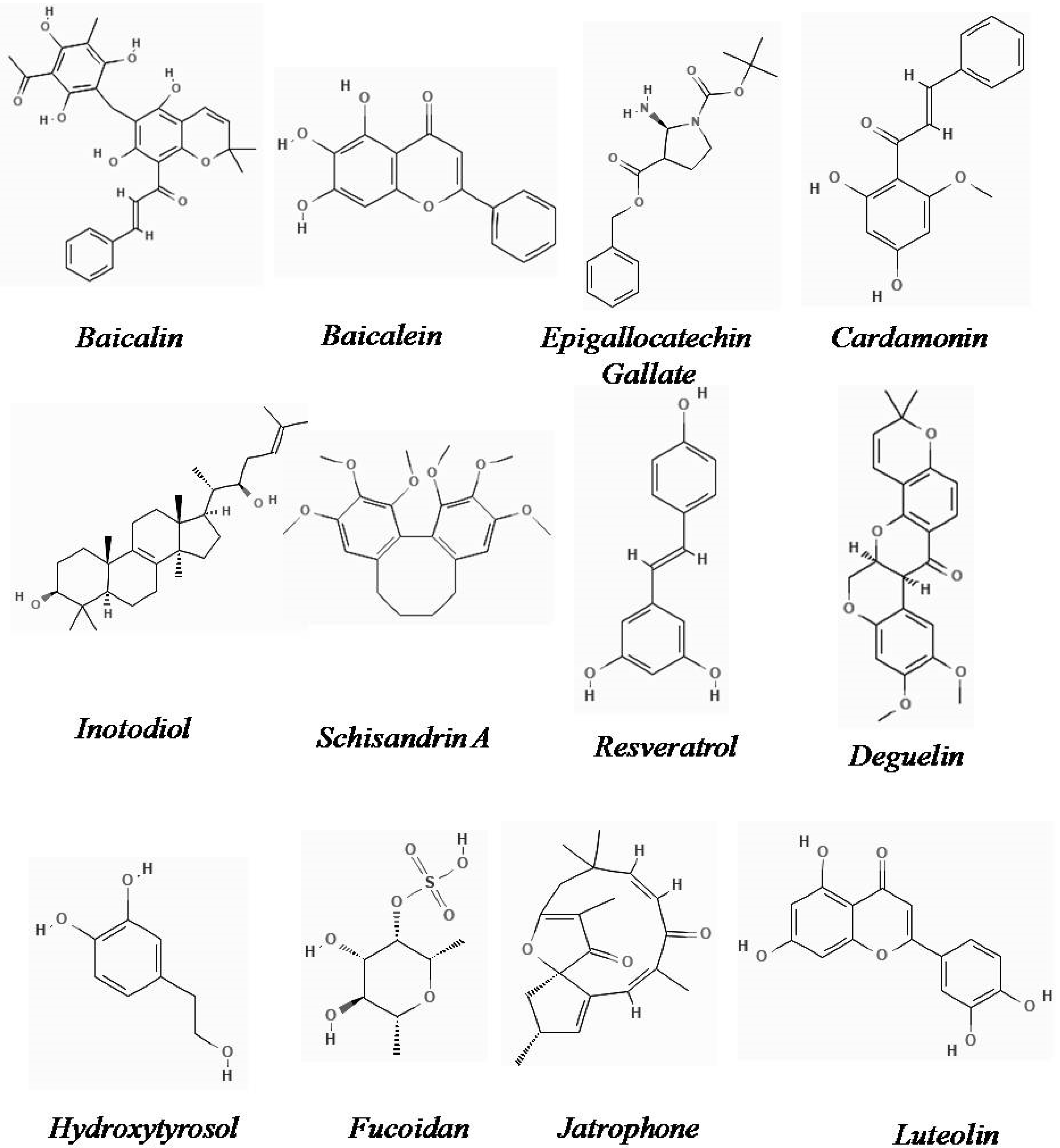
2.9. Baicalin
2.10. Baicalein
2.11. Epigallocatechin Gallate
2.12. Cardamonin
2.13. Inotodiol
2.14. Schisandrin A
2.15. Resveratrol
2.16. Deguelin
2.17. Hydroxytyrosol
2.18. Fucoidan
2.19. Jatrophone
2.20. Luteolin
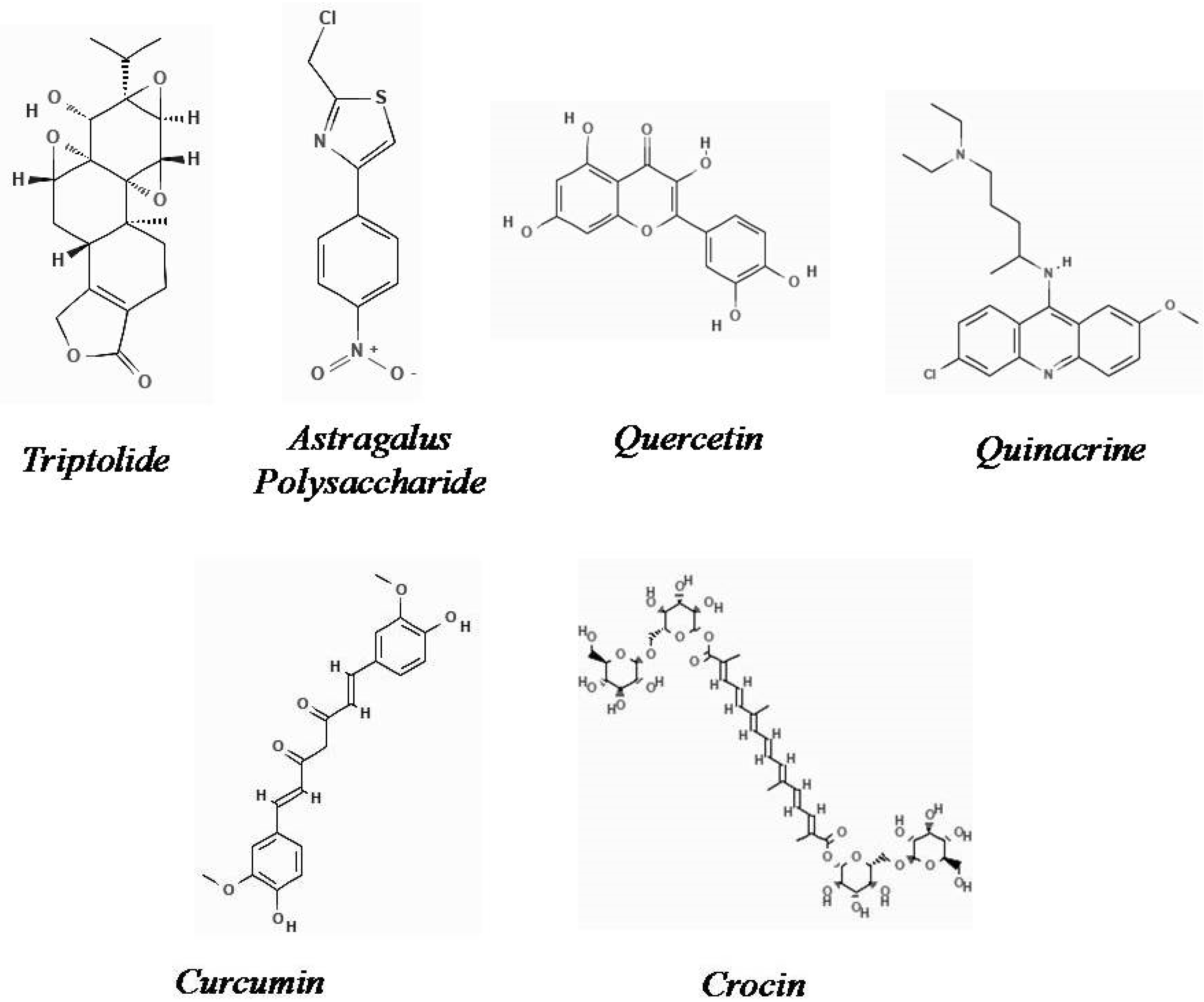
2.21. Triptolide
2.22. Astragalus Polysaccharide
2.23. Quercetin
2.24. Quinacrine
2.25. Curcumin
2.26. Crocin
3. Natural Pill or Extracts Targeting TNBC via Impeding the Wnt/β-Catenin Pathway
3.1. Liuwei Dihuang Pill
3.2. Syzygium guineense
3.3. Ganoderma lucidum
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Al-Thoubaity, F.K. Molecular classification of breast cancer: A retrospective cohort study. Ann. Med. Surg. 2020, 49, 44–48. [Google Scholar] [CrossRef]
- Yin, L.; Duan, J.J.; Bian, X.W.; Yu, S.C. Triple-negative breast cancer molecular subtyping and treatment progress. Breast Cancer Res. 2020, 22, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Foulkes, W.D.; Smith, I.E.; Reis-Filho, J.S. Triple-negative breast cancer. N. Engl. J. Med. 2010, 363, 1938–1948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jitariu, A.A.; Cîmpean, A.M.; Ribatti, D.; Raica, M. Triple negative breast cancer: The kiss of death. Oncotarget 2017, 8, 46652. [Google Scholar] [CrossRef] [Green Version]
- Fultang, N.; Chakraborty, M.; Peethambaran, B. Regulation of cancer stem cells in triple negative breast cancer. Cancer Drug Resist. 2021, 4, 321–342. [Google Scholar] [CrossRef]
- Pohl, S.G.; Brook, N.; Agostino, M.; Arfuso, F.; Kumar, A.P.; Dharmarajan, A. Wnt signaling in triple-negative breast cancer. Oncogenesis 2017, 6, e310. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, N.; Hossain, U.; Mandal, A.; Sil, P.C. The Wnt signaling pathway: A potential therapeutic target against cancer. Ann. N. Y. Acad. Sci. 2019, 1443, 54–74. [Google Scholar] [CrossRef]
- Harb, J.; Lin, P.J.; Hao, J. Recent development of Wnt signaling pathway inhibitors for cancer therapeutics. Curr. Oncol. Rep. 2019, 21, 12. [Google Scholar] [CrossRef]
- Merikhian, P.; Eisavand, M.R.; Farahmand, L. Triple-negative breast cancer: Understanding Wnt signaling in drug resistance. Cancer Cell Int. 2021, 21, 1–8. [Google Scholar] [CrossRef]
- Komiya, Y.; Habas, R. Wnt signal transduction pathways. Organogenesis 2008, 4, 68–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pai, S.G.; Carneiro, B.A.; Mota, J.M.; Costa, R.; Leite, C.A.; Barroso-Sousa, R.; Giles, F.J. Wnt/beta-catenin pathway: Modulating anticancer immune response. J. Hematol. Oncol. 2017, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Yin, S.Y.; Wei, W.C.; Jian, F.Y.; Yang, N.S. Therapeutic applications of herbal medicines for cancer patients. Evid. Based Complementary Altern. Med. 2013, 2013, 302426. [Google Scholar] [CrossRef] [PubMed]
- Tavakoli, J.; Miar, S.; Zadehzare, M.M.; Akbari, H. Evaluation of effectiveness of herbal medication in cancer care: A review study. Iran. J. Cancer Prev. 2012, 5, 144. [Google Scholar] [PubMed]
- Ghorbani-Abdi-Saedabad, A.; Hanafi-Bojd, M.Y.; Parsamanesh, N.; Tayarani-Najaran, Z.; Mollaei, H.; Hoshyar, R. Anticancer and apoptotic activities of parthenolide in combination with epirubicin in mda-mb-468 breast cancer cells. Mol. Biol. Rep. 2020, 47, 5807–5815. [Google Scholar] [CrossRef]
- Blagodatski, A.; Sova, V.; Gorovoy, P.; Usov, A.; Katanaev, V. Novel activators and inhibitors of the Wnt signaling pathway from medicinal plants. Planta Med. 2014, 80, SL29. [Google Scholar] [CrossRef]
- Richard, T.S.; Kamdje, A.H.N.; Mukhtar, F. Medicinal plants in breast cancer therapy. J. Dis. Med. Plants 2015, 1, 19–23. [Google Scholar]
- Yang, Z.; Zhang, Q.; Yu, L.; Zhu, J.; Cao, Y.; Gao, X. The signaling pathways and targets of traditional Chinese medicine and natural medicine in triple-negative breast cancer. J. Ethnopharmacol. 2021, 264, 113249. [Google Scholar] [CrossRef]
- Yuan, B.; Yang, R.; Ma, Y.; Zhou, S.; Zhang, X.; Liu, Y. A systematic review of the active saikosaponins and extracts isolated from Radix Bupleuri and their applications. Pharm. Biol. 2017, 55, 620–635. [Google Scholar] [CrossRef] [Green Version]
- Wong, V.K.W.; Zhang, M.M.; Zhou, H.; Lam, K.Y.C.; Chan, P.L.; Law, C.K.M.; Liu, L. Saikosaponin-d enhances the anticancer potency of TNF-via overcoming its undesirable response of activating NF-Kappa B signalling in cancer cells. Evid. -Based Complementary Altern. Med. 2013, 2013, 745295. [Google Scholar]
- Wong, V.K.; Li, T.; Law, B.Y.; Ma, E.D.; Yip, N.C.; Michelangeli, F.; Liu, L. Saikosaponin-d, a novel SERCA inhibitor, induces autophagic cell death in apoptosis-defective cells. Cell Death Dis. 2013, 4, e720. [Google Scholar] [CrossRef] [PubMed]
- Zhong, D.; Zhang, H.J.; Jiang, Y.D.; Wu, P.; Qi, H.; Cai, C.; Dang, Q. Saikosaponin-d: A potential chemotherapeutics in castration resistant prostate cancer by suppressing cancer metastases and cancer stem cell phenotypes. Biochem. Biophys. Res. Commun. 2016, 474, 722–729. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Qi, H.; Zhang, X.; Si, W.; Xu, F.; Hou, T.; Liang, X. Saikosaponin D from Radix Bupleuri suppresses triple-negative breast cancer cell growth by targeting β-catenin signaling. Biomed. Pharmacother. 2018, 108, 724–733. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yang, L.; Dong, Y.; Zhang, B.; Ma, X. Echinacoside, an inestimable natural product in treatment of neurological and other disorders. Molecules 2018, 23, 1213. [Google Scholar] [CrossRef] [Green Version]
- Tang, C.; Gong, L.; Qiu, K.; Zhang, Z.; Wan, L. Echinacoside inhibits breast cancer cells by suppressing the Wnt/β-catenin signaling pathway. Biochem. Biophys. Res. Commun. 2020, 526, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Houghton, C.A. Sulforaphane: Its “coming of age” as a clinically relevant nutraceutical in the prevention and treatment of chronic disease. Oxidative Med. Cell. Longev. 2019, 2019, 2716870. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhang, T.; Korkaya, H.; Liu, S.; Lee, H.F.; Newman, B.; Sun, D. Sulforaphane, a dietary component of broccoli/broccoli sprouts, inhibits breast cancer stem cells. Clin. Cancer Res. 2010, 16, 2580–2590. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Lu, C.; Li, X.; Fang, H.; Wan, W.; Yang, Q.; Wei, X. Synthesis and biological evaluation of novel gigantol derivatives as potential agents in prevention of diabetic cataract. PLoS ONE 2015, 10, e0141092. [Google Scholar] [CrossRef]
- Yu, S.; Wang, Z.; Su, Z.; Song, J.; Zhou, L.; Sun, Q.; Lu, D. Gigantol inhibits Wnt/β-catenin signaling and exhibits anticancer activity in breast cancer cells. BMC Complementary Altern. Med. 2018, 18, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.; Qi, Q.L.; Wang, M.T.; Li, Q.Y. Therapeutic potential of naringin: An overview. Pharm. Biol. 2016, 54, 3203–3210. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Yang, B.; Huang, J.; Xiang, T.; Yin, X.; Wan, J.; Ren, G. Naringin inhibits growth potential of human triple-negative breast cancer cells by targeting β-catenin signaling pathway. Toxicol. Lett. 2013, 220, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Trejo-Solis, C.; Escamilla-Ramirez, A.; Jimenez-Farfan, D.; Castillo-Rodriguez, R.A.; Flores-Najera, A.; Cruz-Salgado, A. Crosstalk of the Wnt/β-catenin signaling pathway in the induction of apoptosis on cancer cells. Pharmaceuticals 2021, 14, 871. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.L.; Xiang, X.H.; Xia, S.H. Potential signaling pathways involved in the clinical application of oxymatrine. Phytother. Res. 2016, 30, 1104–1112. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Zhang, Y.; Zhang, S.; Wang, F.; Zhang, K.; Huang, Y.; Wang, J. Oxymatrine enhanced anti-tumor effects of Bevacizumab against triple-negative breast cancer via abating Wnt/β-Catenin signaling pathway. Am. J. Cancer Res. 2019, 9, 1796. [Google Scholar]
- Bear, H.D.; Tang, G.; Rastogi, P.; Geyer Jr, C.E.; Robidoux, A.; Atkins, J.N.; Wolmark, N. Bevacizumab added to neoadjuvant chemotherapy for breast cancer. N. Engl. J. Med. 2012, 366, 310–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.; Lan, X.; Xu, S.; Li, S.; Xi, Y. Efficacy of bevacizumab combined with chemotherapy in the treatment of HER2-negative metastatic breast cancer: A network meta-analysis. BMC Cancer 2020, 20, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Delmas, D.; Xiao, J.; Vejux, A.; Aires, V. Silymarin and cancer: A dual strategy in both in chemoprevention and chemosensitivity. Molecules 2020, 25, 2009. [Google Scholar] [CrossRef]
- Lu, W.; Lin, C.; King, T.D.; Chen, H.; Reynolds, R.C.; Li, Y. Silibinin inhibits Wnt/β-catenin signaling by suppressing Wnt co-receptor LRP6 expression in human prostate and breast cancer cells. Cell. Signal. 2012, 24, 2291–2296. [Google Scholar] [CrossRef] [Green Version]
- King, T.D.; Suto, M.J.; Li, Y. The wnt/β-catenin signaling pathway: A potential therapeutic target in the treatment of triple negative breast cancer. J. Cell. Biochem. 2012, 113, 13–18. [Google Scholar] [CrossRef]
- Ma, J.; Hou, Y.; Xia, J.; Zhu, X.; Wang, Z.P. Tumor suppressive role of rottlerin in cancer therapy. Am. J. Transl. Res. 2018, 10, 3345. [Google Scholar]
- Lu, W.; Lin, C.; Li, Y. Rottlerin induces Wnt co-receptor LRP6 degradation and suppresses both Wnt/β-catenin and mTORC1 signaling in prostate and breast cancer cells. Cell. Signal. 2014, 26, 1303–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, S.A.L.; Huang, S. Crosstalk of the Wnt/b-catenin pathway with other pathways in cancer cells. Genes Dis. 2015, 3, 41–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.; Liang, Y.; Cheng, A.; Wang, Q.; Li, Y.; Wei, H.; Wan, X. Antiviral properties of baicalin: A concise review. Rev. Bras. De Farmacogn. 2021, 31, 408–419. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Zhang, A.; Kuang, G.; Gong, X.; Jiang, R.; Lin, D.; Li, H. Baicalin inhibits the metastasis of highly aggressive breast cancer cells by reversing epithelial-to-mesenchymal transition by targeting β-catenin signaling. Oncol. Rep. 2017, 38, 3599–3607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Snyder, S.A.; Smith, J.N.; Chen, Y.C. Anticancer properties of baicalein: A review. Med. Chem. Res. 2016, 25, 1515–1523. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Yan, W.; Dai, Z.; Gao, X.; Ma, Y.; Xu, Q.; Zhang, S. Baicalein suppresses metastasis of breast cancer cells by inhibiting EMT via downregulation of SATB1 and Wnt/β-catenin pathway. Drug Des. Dev. Ther. 2016, 10, 1419. [Google Scholar] [CrossRef] [Green Version]
- Gan, R.Y.; Li, H.B.; Sui, Z.Q.; Corke, H. Absorption, metabolism, anti-cancer effect and molecular targets of epigallocatechin gallate (EGCG): An updated review. Crit. Rev. Food Sci. Nutr. 2018, 58, 924–941. [Google Scholar] [CrossRef]
- Hong, O.Y.; Noh, E.M.; Jang, H.Y.; Lee, Y.R.; Lee, B.K.; Jung, S.H.; Youn, H.J. Epigallocatechin gallate inhibits the growth of MDA-MB-231 breast cancer cells via inactivation of the β-catenin signaling pathway. Oncol. Lett. 2017, 14, 441–446. [Google Scholar] [CrossRef] [Green Version]
- Nelson, C.M.; Chen, C.S. Cell-cell signaling by direct contact increases cell proliferation via a PI3K-dependent signal. FEBS Lett. 2002, 514, 238–242. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Zhang, X.; Rieger-Christ, K.M.; Summerhayes, I.C.; Wazer, D.E.; Paulson, K.E.; Yee, A.S. Suppression of Wnt signaling by the green tea compound EGCG in breast cancer and the requirement for the HBP1 transcriptional repressor. Cancer Res. 2006, 66 (Suppl. 8), 30. [Google Scholar]
- Sampson, E.M.; Haque, Z.K.; Ku, M.C.; Tevosian, S.G.; Albanese, C.; Pestell, R.G.; Yee, A.S. Negative regulation of the Wnt–β-catenin pathway by the transcriptional repressor HBP1. EMBO J. 2001, 20, 4500–4511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawaz, J.; Rasul, A.; Shah, M.A.; Hussain, G.; Riaz, A.; Sarfraz, I.; Selamoglu, Z. Cardamonin: A new player to fight cancer via multiple cancer signaling pathways. Life Sci. 2020, 250, 117591. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, S.; Jeengar, M.K.; Thummuri, D.; Koval, A.; Katanaev, V.L.; Marepally, S.; Naidu, V.G.M. Cardamonin, a chalcone, inhibits human triple negative breast cancer cell invasiveness by downregulation of Wnt/β-catenin signaling cascades and reversal of epithelial–mesenchymal transition. Biofactors 2017, 43, 152–169. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.D.; Yu, L.; Wang, P.; Kou, P.; Li, J.; Wang, L.T.; Fu, Y.J. Inotodiol inhibits cells migration and invasion and induces apoptosis via p53-dependent pathway in HeLa cells. Phytomedicine 2019, 60, 152957. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.K.; Kang, J.H.; Song, K.H.; Park, M.S.; Kim, D.K.; Park, Y.J.; Oh, S.H. Inonotus obliquus suppresses proliferation of colorectal cancer cells and tumor growth in mice models by downregulation of β-catenin/NF-κB-signaling pathways. Eur. J. Inflamm. 2013, 11, 615–629. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Bao, C.; Zhang, J. Inotodiol suppresses proliferation of breast cancer in rat model of type 2 diabetes mellitus via downregulation of β-catenin signaling. Biomed. Pharmacother. 2018, 99, 142–150. [Google Scholar] [CrossRef]
- Kwon, Y.J.; Ye, D.J.; Baek, H.S.; Chun, Y.J. 7,12-Dimethylbenz [α] anthracene increases cell proliferation and invasion through induction of Wnt/β-catenin signaling and EMT process. Environ. Toxicol. 2018, 33, 729–742. [Google Scholar] [CrossRef]
- Garcia-Jimenez, C.; García-Martínez, J.M.; Chocarro-Calvo, A.; De la Vieja, A. A new link between diabetes and cancer: Enhanced WNT/β-catenin signaling by high glucose. J. Mol. Endocrinol. 2014, 52, R51–R66. [Google Scholar] [CrossRef] [Green Version]
- Kopustinskiene, D.M.; Bernatoniene, J. Antioxidant Effects of Schisandra chinensis Fruits and Their Active Constituents. Antioxidants 2021, 10, 620. [Google Scholar] [CrossRef]
- Xu, X.; Rajamanicham, V.; Xu, S.; Liu, Z.; Yan, T.; Liang, G.; Wang, Y. Schisandrin A inhibits triple negative breast cancer cells by regulating Wnt/ER stress signaling pathway. Biomed. Pharmacother. 2019, 115, 108922. [Google Scholar] [CrossRef]
- McGrath, E.P.; Logue, S.E.; Mnich, K.; Deegan, S.; Jäger, R.; Gorman, A.M.; Samali, A. The unfolded protein response in breast cancer. Cancers 2018, 10, 344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berman, A.Y.; Motechin, R.A.; Wiesenfeld, M.Y.; Holz, M.K. The therapeutic potential of resveratrol: A review of clinical trials. NPJ Precis. Oncol. 2017, 1, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuthapisith, S.; Eremin, J.; El-Sheemey, M.; Eremin, O. Breast cancer chemoresistance: Emerging importance of cancer stem cells. Surg. Oncol. 2010, 19, 27–32. [Google Scholar] [CrossRef]
- Fu, Y.; Chang, H.; Peng, X.; Bai, Q.; Yi, L.; Zhou, Y.; Mi, M. Resveratrol inhibits breast cancer stem-like cells and induces autophagy via suppressing Wnt/β-catenin signaling pathway. PLoS ONE 2014, 9, e102535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindeman, G.J.; Visvader, J.E. Insights into the cell of origin in breast cancer and breast cancer stem cells. Asia-Pac. J. Clin. Oncol. 2010, 6, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Boyd, J.; Han, A. Deguelin and its role in chronic diseases. Drug Discov. Mother Nat. 2016, 929, 363–375. [Google Scholar]
- Murillo, G.; Peng, X.; Torres, K.E.; Mehta, R.G. Deguelin inhibits growth of breast cancer cells by modulating the expression of key members of the Wnt signaling pathway. Cancer Prev. Res. 2009, 2, 942–950. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, R.; Bahamonde, C.; Sanz, C.; Pérez, A.G. Identification and Functional Characterization of Genes Encoding Phenylacetaldehyde Reductases That Catalyze the Last Step in the Biosynthesis of Hydroxytyrosol in Olive. Plants 2021, 10, 1268. [Google Scholar] [CrossRef]
- Cruz-Lozano, M.; González-González, A.; Marchal, J.A.; Muñoz-Muela, E.; Molina, M.P.; Cara, F.E.; Granados-Principal, S. Hydroxytyrosol inhibits cancer stem cells and the metastatic capacity of triple-negative breast cancer cell lines by the simultaneous targeting of epithelial-to-mesenchymal transition, Wnt/β-catenin and TGFβ signaling pathways. Eur. J. Nutr. 2019, 58, 3207–3219. [Google Scholar] [CrossRef]
- Vikram, R.; Chou, W.C.; Hung, S.C.; Shen, C.Y. Tumorigenic and metastatic role of CD44−/low/CD24−/low cells in luminal breast cancer. Cancers 2020, 12, 1239. [Google Scholar] [CrossRef]
- Li, B.; Lu, F.; Wei, X.; Zhao, R. Fucoidan: Structure and bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar] [CrossRef] [Green Version]
- Xue, M.; Ge, Y.; Zhang, J.; Liu, Y.; Wang, Q.; Hou, L.; Zheng, Z. Fucoidan inhibited 4T1 mouse breast cancer cell growth in vivo and in vitro via downregulation of Wnt/β-catenin signaling. Nutr. Cancer 2013, 65, 460–468. [Google Scholar] [CrossRef]
- Zhurinsky, J.; Shtutman, M.; Ben-Ze’ev, A. Plakoglobin and beta-catenin: Protein interactions, regulation and biological roles. J. Cell Sci. 2000, 113, 3127–3139. [Google Scholar] [CrossRef]
- Devappa, R.K.; Makkar, H.P.; Becker, K. Nutritional, biochemical, and pharmaceutical potential of proteins and peptides from Jatropha. J. Agric. Food Chem. 2010, 58, 6543–6555. [Google Scholar] [CrossRef]
- Fatima, I.; El-Ayachi, I.; Taotao, L.; Lillo, M.A.; Krutilina, R.; Seagroves, T.N.; Miranda-Carboni, G.A. The natural compound Jatrophone interferes with Wnt/β-catenin signaling and inhibits proliferation and EMT in human triple-negative breast cancer. PLoS ONE 2017, 12, e0189864. [Google Scholar] [CrossRef] [Green Version]
- Imran, M.; Rauf, A.; Abu-Izneid, T.; Nadeem, M.; Shariati, M.A.; Khan, I.A.; Mubarak, M.S. Luteolin, a flavonoid, as an anticancer agent: A review. Biomed. Pharmacother. 2019, 112, 108612. [Google Scholar] [CrossRef]
- Lin, D.; Kuang, G.; Wan, J.; Zhang, X.; Li, H.; Gong, X.; Li, H. Luteolin suppresses the metastasis of triple-negative breast cancer by reversing epithelial-to-mesenchymal transition via downregulation of β-catenin expression. Oncol. Rep. 2017, 37, 895–902. [Google Scholar] [CrossRef] [Green Version]
- Tong, L.; Zhao, Q.; Datan, E.; Lin, G.Q.; Minn, I.; Pomper, M.G.; Liu, J.O. Triptolide: Reflections on two decades of research and prospects for the future. Nat. Prod. Rep. 2021, 38, 843–860. [Google Scholar] [CrossRef]
- Shao, H.; Ma, J.; Guo, T.; Hu, R. Triptolide induces apoptosis of breast cancer cells via a mechanism associated with the Wnt/β catenin signaling pathway. Exp. Ther. Med. 2014, 8, 505–508. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Ren, W.; Zhang, L.; Zhang, Y.; Liu, D.; Liu, Y. A review of the pharmacological action of Astragalus polysaccharide. Front. Pharmacol. 2020, 11, 349. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Sun, S.; Xu, W.; Yu, B.; Wang, G.; Wang, H. Astragalus polysaccharide inhibits breast cancer cell migration and invasion by regulating epithelial-mesenchymal transition via the Wnt/β-catenin signaling pathway. Mol. Med. Rep. 2020, 21, 1819–1832. [Google Scholar] [CrossRef] [Green Version]
- David, A.V.A.; Arulmoli, R.; Parasuraman, S. Overviews of biological importance of quercetin: A bioactive flavonoid. Pharmacogn. Rev. 2016, 10, 84. [Google Scholar]
- Srinivasan, A.; Thangavel, C.; Liu, Y.; Shoyele, S.; Den, R.B.; Selvakumar, P.; Lakshmikuttyamma, A. Quercetin regulates β-catenin signaling and reduces the migration of triple negative breast cancer. Mol. Carcinog. 2016, 55, 743–756. [Google Scholar] [CrossRef]
- Curtin, J.C.; Lorenzi, M.V. Drug discovery approaches to target Wnt signaling in cancer stem cells. Oncotarget 2010, 1, 563. [Google Scholar] [CrossRef] [Green Version]
- Oien, D.B.; Pathoulas, C.L.; Ray, U.; Thirusangu, P.; Kalogera, E.; Shridhar, V. Repurposing quinacrine for treatment-refractory cancer. Semin. Cancer Biol. 2021, 68, 21–30. [Google Scholar] [CrossRef]
- Preet, R.; Mohapatra, P.; Mohanty, S.; Sahu, S.K.; Choudhuri, T.; Wyatt, M.D.; Kundu, C.N. Quinacrine has anticancer activity in breast cancer cells through inhibition of topoisomerase activity. Int. J. Cancer 2012, 130, 1660–1670. [Google Scholar] [CrossRef]
- Preet, R.; Mohapatra, P.; Das, D.; Satapathy, S.R.; Choudhuri, T.; Wyatt, M.D.; Kundu, C.N. Lycopene synergistically enhances quinacrine action to inhibit Wnt-TCF signaling in breast cancer cells through APC. Carcinogenesis 2013, 34, 277–286. [Google Scholar] [CrossRef] [Green Version]
- Vallée, A.; Lecarpentier, Y.; Vallée, J.N. Curcumin: A therapeutic strategy in cancers by inhibiting the canonical WNT/β-catenin pathway. J. Exp. Clin. Cancer Res. 2019, 38, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Prasad, C.P.; Rath, G.; Mathur, S.; Bhatnagar, D.; Ralhan, R. Potent growth suppressive activity of curcumin in human breast cancer cells: Modulation of Wnt/β-catenin signaling. Chem. -Biol. Interact. 2009, 181, 263–271. [Google Scholar] [CrossRef]
- Li, X.; Wang, X.; Xie, C.; Zhu, J.; Meng, Y.; Chen, Y.; Zhao, Y. Sonic hedgehog and Wnt/β-catenin pathways mediate curcumin inhibition of breast cancer stem cells. Anti-Cancer Drugs 2018, 29, 208–215. [Google Scholar] [CrossRef]
- Kakarala, M.; Brenner, D.E.; Korkaya, H.; Cheng, C.; Tazi, K.; Ginestier, C.; Wicha, M.S. Targeting breast stem cells with the cancer preventive compounds curcumin and piperine. Breast Cancer Res. Treat. 2010, 122, 777–785. [Google Scholar] [CrossRef] [Green Version]
- Hoshyar, R.; Mollaei, H. A comprehensive review on anticancer mechanisms of the main carotenoid of saffron, crocin. J. Pharm. Pharmacol. 2017, 69, 1419–1427. [Google Scholar] [CrossRef] [Green Version]
- Arzi, L.; Hoshyar, R. Saffron anti-metastatic properties, ancient spice novel application. Crit. Rev. Food Sci. Nutr. 2021, 62, 1–12. [Google Scholar] [CrossRef]
- Arzi, L.; Riazi, G.; Sadeghizadeh, M.; Hoshyar, R.; Jafarzadeh, N. A comparative study on anti-invasion, antimigration, and antiadhesion effects of the bioactive carotenoids of saffron on 4T1 breast cancer cells through their effects on Wnt/β-catenin pathway genes. DNA Cell Biol. 2018, 37, 697–707. [Google Scholar] [CrossRef]
- Arzi, L.; Farahi, A.; Jafarzadeh, N.; Riazi, G.; Sadeghizadeh, M.; Hoshyar, R. Inhibitory effect of crocin on metastasis of triple-negative breast cancer by interfering with Wnt/β-catenin pathway in murine model. DNA Cell Biol. 2018, 37, 1068–1075. [Google Scholar] [CrossRef]
- Arzi, L.; Hoshyar, R.; Jafarzadeh, N.; Riazi, G.; Sadeghizadeh, M. Anti-metastatic properties of a potent herbal combination in cell and mice models of triple negative breast cancer. Life Sci. 2020, 243, 117245. [Google Scholar] [CrossRef]
- Feng, P.; Che, Y.; Chen, D.Q. Molecular mechanism of action of LiuweiDihuang pill for the treatment of osteoporosis based on network pharmacology and molecular docking. Eur. J. Integr. Med. 2020, 33, 101009. [Google Scholar] [CrossRef]
- Limopasmanee, W.; Chansakaow, S.; Rojanasthien, N.; Manorot, M.; Sangdee, C.; Teekachunhatean, S. Effects of the Chinese herbal formulation (Liu Wei Di Huang Wan) on the pharmacokinetics of isoflavones in postmenopausal women. BioMed Res. Int. 2015, 2015, 902702. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Yao, K.; Yang, X.; Liu, W.; Feng, B.; Ma, J.; Xiong, X. Chinese patent medicine liuwei di huang wan combined with antihypertensive drugs, a new integrative medicine therapy, for the treatment of essential hypertension: A systematic review of randomized controlled trials. Evid. -Based Complementary Altern. Med. 2012, 2012, 714805. [Google Scholar]
- Lixiang, Z.; Hongning, L.; Yan, G.; Xianming, M.; Runde, J.; Xiaomin, W.; Yue, W. Effect of Liuweidihuang pill and Jinkuishenqi pill on inhibition of spontaneous breast carcinoma growth in mice. J. Tradit. Chin. Med. 2015, 35, 453–459. [Google Scholar] [CrossRef] [Green Version]
- Lixiang, Z.; Qing, Z.; Zhipeng, Y.; Jian, W.; Xiaoying, R.; Yan, G.; Haizhou, L. LiuweiDihuang pill suppresses metastasis by regulating the wnt pathway and disrupting β-catenin/T cell factor interactions in a murine model of triple-negative breast cancer. J. Tradit. Chin. Med. 2019, 39, 826–832. [Google Scholar]
- Tadesse, S.A.; Wubneh, Z.B. Antimalarial activity of Syzygiumguineense during early and established Plasmodium infection in rodent models. BMC Complementary Altern. Med. 2017, 17, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koval, A.; Pieme, C.A.; Queiroz, E.F.; Ragusa, S.; Ahmed, K.; Blagodatski, A.; Katanaev, V.L. Tannins from Syzygium guineense suppress Wnt signaling and proliferation of Wnt-dependent tumors through a direct effect on secreted Wnts. Cancer Lett. 2018, 435, 110–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kladar, N.V.; Gavarić, N.S.; Božin, B.N. Ganoderma: Insights into anticancer effects. Eur. J. Cancer Prev. 2016, 25, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Suarez-Arroyo, I.J.; Rosario-Acevedo, R.; Aguilar-Perez, A.; Clemente, P.L.; Cubano, L.A.; Serrano, J.; Martínez-Montemayor, M.M. Anti-tumor effects of Ganoderma lucidum (reishi) in inflammatory breast cancer in in vivo and in vitro models. PLoS ONE 2013, 8, e57431. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y. Ganoderma lucidum (Reishi) suppresses proliferation and migration of breast cancer cells via inhibiting Wnt/β-catenin signaling. Biochem. Biophys. Res. Commun. 2017, 488, 679–684. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Active Pharmaceutical Ingredient | Plant | Model of Study | Dose | Targeted Genes | Impact on TNBC | Reference |
|---|---|---|---|---|---|---|
| Saikosaponin D | Radix Bupleuri | HCC1937 MDA-MB-468 MDA-MB-231 | 10, 15 and 20 µM | β-Catenin↓ Cyclin D1↓ c-Myc↓ | Proliferation↓ Apoptosis↑ | [23] |
| Echinacoside | Cistanche and Echinacea | MDA-MB-468 MDA-MB-231 | 25–100µM | p-LRP6↓ total LRP6↓ p-Dvl2↓ active β-Catenin↓ total β-Catenin↓ CD44↓ LEF1↓ Cyclin D↓ | Proliferation↓ Migration↓ Invasion↓ Tumor sizes↓ Tumor weights↓ | [25] |
| Nude mice | 10 mg/kg | |||||
| Sulforaphane | Brassica | SUM159 | 0.5, 1 and 5 µM | β-Catenin↓ Cyclin D1↓ | Proliferation↓ Apoptosis↑ Mammosphere formation↓ BCSC activity↓ Tumor sizes↓ | [27] |
| NOD/SCID mice | 50 mg/kg | |||||
| Gigantol | Orchidaceae | MDA-MB-231 MDA-MB-468 | 0–100 µM | p-LRP6,↓ total LRP6↓ Cytosolic β-catenin↓ Axin2↓ Survivin↓ | Proliferation↓ Migration↓ | [29] |
| Naringin | Drynaria fortunei Citrus aurantium Citrus medicaand citrus fruit | MDA-MB-231 MDA-MB-468 BT-549 | 0–200 µM | Survivin↓ P21↓ β-Catenin ↓ Cyclin E↓ Rb ↓ p-Rb↓ | Proliferation↓ Apoptosis↑ Arrest cell cycle in G1 Tumor sizes↓ Tumor weights↓ | [31] |
| SCID hairless mice | 100 mg/kg | |||||
| Oxymatrine | Sophora Flavescens Ait | MDA-MB-231 MDA-MB-468 | 0, 1, 2 and 4 mM Bevacizumab 200 nM | N-Cad↓ Vim↓ ZEB↓ Snail↓ Slug↓ E-Cad↑ β-Catenin↓ c-Myc↓ Cyclin D1↓ CD44↓ VegfA↓ | Migration↓ Invasion↓ EMT↓ BCSC self-renewal↓ Tumor size↓ Angiogenesis↓ | [34] |
| BALB/c nude mice | 25 mg/kg + 5 mg/kg Bevacizumab | |||||
| Silibinin | Silybum marianum. Gaertn | MDA-MB-231 | 0–200 µM | LRP6↓ p-LRP6↓ Axin2↓ | Proliferation↓ | [38] |
| Rottlerin | Mallotus philippinensis | MDA-MB-231 | 0.1–31.6 μM | cytosolicβ-catenin↓ totalβ-Catenin↓ LRP6↓ p--LRP6↓ Axin2↓ P70-S6K↓ p-P70-S6K↓ S6↓ p-S6↓ CyclinD1↓ Survivin↓ | Proliferation↓ | [41] |
| Baicalin | Scutellaria baicalensis Georgi | MDA-MB-231 4T1 | 10, 30 and 100 μM | β-Catenin↓ E-Cad↑ Claudin↑ N-Cad↓ VIM↓ Snail↓ Slug↓ | Proliferation↓ Migration↓ Invasion↓ EMT↓ Metastatic colonies in liver and lung↓ | [44] |
| BALB/c mice | 100 mg/kg | |||||
| Baicalein | Scutellaria baicalensis Georgi | MDA-MB-231 | 10, 20, and 40 μM/L | E-Cad↑ Vim↓ Snail↓ Wnt1↓ β-Catenin↓ SATB1↓ Cyclin D1↓ Axin2↓ | Proliferation↓ Migration↓ Invasion↓ EMT↓ Metastatic colonies in liver and lung↓ | [46] |
| BALB/c nude mice | 50 or 100 mg/kg | |||||
| Epigallocatechin gallate | Green tea | MDA-MB-231 | 25, 50, 75, 100 and 200 μM | β-Catenin↓ p-AKT↓ Cyclin D1↓ | Proliferation↓ | [48] |
| MDA-MB-231 | 25–100 μM | HBP1↓ β-Catenin↓ c-Myc↓ | Proliferation↓ Migration↓ Invasion↓ | [50] | ||
| Cardamonin | Zingiberaceae | MDA-MB-231 BT-549 | 0–100 μM | Bax↑ Bcl-2↓ Cyt-C↑ Caspase-3↑ PARP↑ β-Catenin↓ E Cad↑ N-Cad↓ Vim↓ Slug↓ Snail↓ Cyclin D1↓ c-Myc↓ Vegf↓ CDK4↓ | Proliferation↓ Migration↓ Invasion↓ EMT↓ Apoptosis↑ Arrest cell cycle in G2/M Tumor size↓ | [53] |
| BALB/c mice | 2.5 and 5 mg/kg | |||||
| Inotodiol | Inonotus obliquus | Sprague-Dawley rats | 10 mg/kg | PCNA↓ β-Catenin↓ c-Myc↓ Cyclin D1↓ Caspase-3↑ PARP↑ | Proliferation↓ Apoptosis↑ Body weight↑ Antioxidant enzyme activities↑ Abnormal histological signs of pancreas↓ Glucose↓ Cholesterol↓ Triglyceride↓ HDL↓ | [56] |
| Schisandrin | Schisandra chinensis Baill | MDA-MB-231 BT-549 | 25,50 and 100 µM | ATF4↑ CHOP ↑ p-Elf2α↑ β-Catenin↓ p-GSK3β↓ Bax↑ P53↑ Bcl2↓ | Proliferation↓ Arrest cell cycle in G1 Apoptosis↑ ER stress↑ Tumor size↓ | [60] |
| BALB/c mice | 25 mg/kg | |||||
| Resveratrol | Grapes, Berries and Peanuts | SUM159 | 10, 20 and 40µM | LC3-II↑ Beclin1↑ Atg 7↑ β-Catenin↓ Cyclin D1↓ | Proliferation↓ BCSC activity↓ BCSC self-renewal↓ Autophagy↑ Tumor size↓ | [64] |
| NOD/SCID mice | 100 mg/kg | |||||
| Deguelin | Mundulea sericea | MDA-MB-231 | 0.1–10 μM/L | WIF-1↑ DDK4↑ CDH3↑ CDH7↑ CDH9↑ Wnt14↓ Wnt 2B↓ Wnt 3↓ Snail↓ p-GSK-3β↓ β-Catenin↓ Cyclin D1↓ | Proliferation↓ Apoptosis↑ Arrest cell cycle in S | [67] |
| Hydroxytyrosol | Olive oil | SUM159PT BT549 MDA-MB-231 Hs578T | 0–100 μM | Zeb↓ Slug↓ Vim↓ Zo-1↑ p-LRP6↓ LRP6↓ β-Catenin↓ Cyclin D1↓ | BCSC self-renewal↓ Migration↓ Invasion↓ EMT↓ BCSC activity↓ | [69] |
| Fucoidan | Brown seaweed and Marine invertebrates | 4T1 | 50, 100 and 200 µg/mL | β-Catenin↓ c-Myc↓ Cyclin D1↓ Survivin↓ TCF/LEF↓ | Proliferation↓ Apoptosis↑ Arrest cell cycle in G1 Tumor size↓ Tumor weight↓ | [72] |
| BALB/c mice | 5, 10 mg/kg | |||||
| Jatrophone | Euphorbiaceae | MDA-MB-231 MDA-MB-157 HCC38 MDA-MB-468 Patient-derived xenograft | 100 nM- 30 μM | BIRC5↓ Axin2↓ HMGA2↓ Myc↓ PCNA↓ CCND1↓ Cyclin D1↓ β-Catenin↓ Slug↓ Fibronectin↓ Vim↓ | Proliferation↓ Apoptosis↑ Arrest cell cycle in S-phase EMT↓ Migration↓ | [75] |
| Luteolin | Celery Sweet bell peppers, Carrots Broccoli Onion leaves Parsley | MDA-MB-231 BT5–49 | 10, 30 and 100 μM | N-Cad↓ Vim↓ Snail↓ Slug↓ E-Cad↑ Claudin↑ β-Catenin↓ | Migration↓ Invasion↓ EMT↓ Metastatic colonies↓ | [77] |
| Nude mice | 100 mg/kg | |||||
| Triptolide | Tripterygium wilfordii Hook F | MDA-MB-231 | 10, 25 and 50 nM | β-Catenin↓ | Proliferation↓ Apoptosis↑ | [79] |
| Astragalus polysaccharide | Astragalus membranaceus | MDA-MB-231 | 25, 50, 100, 200, 400, 800 and 1600 μg/mL | Vim↓ Snail↓ E-Cad↑ β-Catenin↓ c-Myc↓ Cycline D1↓ | Proliferation↓ Migration↓ Invasion↓ EMT↓ | [81] |
| Quercetin | Fruits, Vegetables, Seeds, Nuts, Green tea, and Red wine | MDA-MB-231 MDA-MB-468 | 10 and 50 μM | Vim↓ E-Cad↑ Cyclin D1↓ c-Myc↓ p-AKT↓ | Proliferation↓ Survival rate↓ Migration↓ Invasion↓ Reshape mesenchymal to epithelial shape | [83] |
| Quinacrine | Cinchona tree | MDA-MB-231 | 5, 10, 15 and 20 μM | β-Catenin↓ Cyclin D1↓ APC↑ | Proliferation↓ Survival rate↓ Apoptosis↑ DNA damage↑ Topoisomerase activity↓ | [86,87] |
| Quinacrine + Lycopen | Cinchona tree Tomato | 5 μM + 2, 4, 8, 10, 12, 14 and 16 μM | β-Catenin↓ Cyclin D1↓ APC↑ | Proliferation↓ Survival ↓ | ||
| Curcumin | Curcuma longa | MDA-MB-231 | 20 μM | Dsh↓ β-Catenin↓, Cyclin D1↓ Slug↓ | Proliferation↓ Apoptosis↑ Arrest cell cycle in G2/M | [89] |
| SUM159 | 10, 20 and 40 μM | CD44↓ ALDH1A1↓ Nanog↓ OCT4↓ PCNA↓ Cyclin D1↓ Bcl2↓ Bax↑. Caspase 8↑ Caspase9↑ cleaved Caspase 3↑ p-GSK3β↓ β-Catenin↓ c-Myc↓ | Proliferation↓ Apoptosis↑ Mammosphere formation↓ BCSC activity↓ | [90] | ||
| Crocin | Crocus sativus | 4T1 | 2.5 and 3 mM | FzD7↓ Nedd9↓ Vegf-α↓ Mmp9↓ Vim↓ E-Cad↑ | Proliferation↓ Invasion↓ Migration↓ Cell–ECM adhesion↓ Tumor size↓ Metastatic colonies↓ | [94] |
| BALB/c mice | 200 mg/kg | [95] | ||||
| Crocin + Crocetin | Crocus sativus | 4T1 | Crocin 2.5 mM + Crocetin 0.05 mM & Crocetin 0.1 mM + Crocin 2 mM | Fzd7↓ Nedd 9↓ Vegf-α↓ Mmp9↓ Vim↓ | Proliferation↓ Invasion↓ Migration↓ Tumor size↓ Cell–ECM adhesion↓ Metastatic colonies↓ | [96] |
| BALB/c mice | Crocin 200 mg/kg + Crocetin 5 mg/kg | |||||
| Liuwei Dihuang pill | Rehmannia glutinosa Dioscorea opposita Cornus officinalis Poria cocos Alisma orientalis Paeonia suffruticosa | Kunming mice | 2.3, 4.6 and 9.2 g/kg | β-Catenin↓ Cyclin D1↓ TCF-1↓ Vegf↓ | Tumor sizes↓ Tumor weights↓ Survival time↑ Metastatic colonies in liver and lung↓ | [101] |
| Tannins | Syzygium guineense Wall | BT-20 HCC38 MDA-MB-231, HCC1806 HCC1395 MDA-MB-468 | 0–100 μg/mL | Wnt3a↓ β-Catenin↓ LRP6↓ | Proliferation↓ | [103] |
| Ganoderma Lucidum | _ | MDA-MB-231 4T1 | 0–200 µg/mL | β-Catenin↓ p-LRP6↓ p-Dvl2↓ Axin2↓ | Proliferation ↓ Migration↓ | [106] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arzi, L.; Mollaei, H.; Hoshyar, R. Countering Triple Negative Breast Cancer via Impeding Wnt/β-Catenin Signaling, a Phytotherapeutic Approach. Plants 2022, 11, 2191. https://doi.org/10.3390/plants11172191
Arzi L, Mollaei H, Hoshyar R. Countering Triple Negative Breast Cancer via Impeding Wnt/β-Catenin Signaling, a Phytotherapeutic Approach. Plants. 2022; 11(17):2191. https://doi.org/10.3390/plants11172191
Chicago/Turabian StyleArzi, Laleh, Homa Mollaei, and Reyhane Hoshyar. 2022. "Countering Triple Negative Breast Cancer via Impeding Wnt/β-Catenin Signaling, a Phytotherapeutic Approach" Plants 11, no. 17: 2191. https://doi.org/10.3390/plants11172191
APA StyleArzi, L., Mollaei, H., & Hoshyar, R. (2022). Countering Triple Negative Breast Cancer via Impeding Wnt/β-Catenin Signaling, a Phytotherapeutic Approach. Plants, 11(17), 2191. https://doi.org/10.3390/plants11172191