
Additive and Non-Additive Effects on the Control of Key Agronomic Traits in Popcorn Lines under Contrasting Phosphorus Conditions

 ,
, 
 ,
, 
 ,
,  , ,
, ,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Genetic Variability for Agronomic Traits Evaluated under Contrasting Phosphorus Conditions in Both Environments
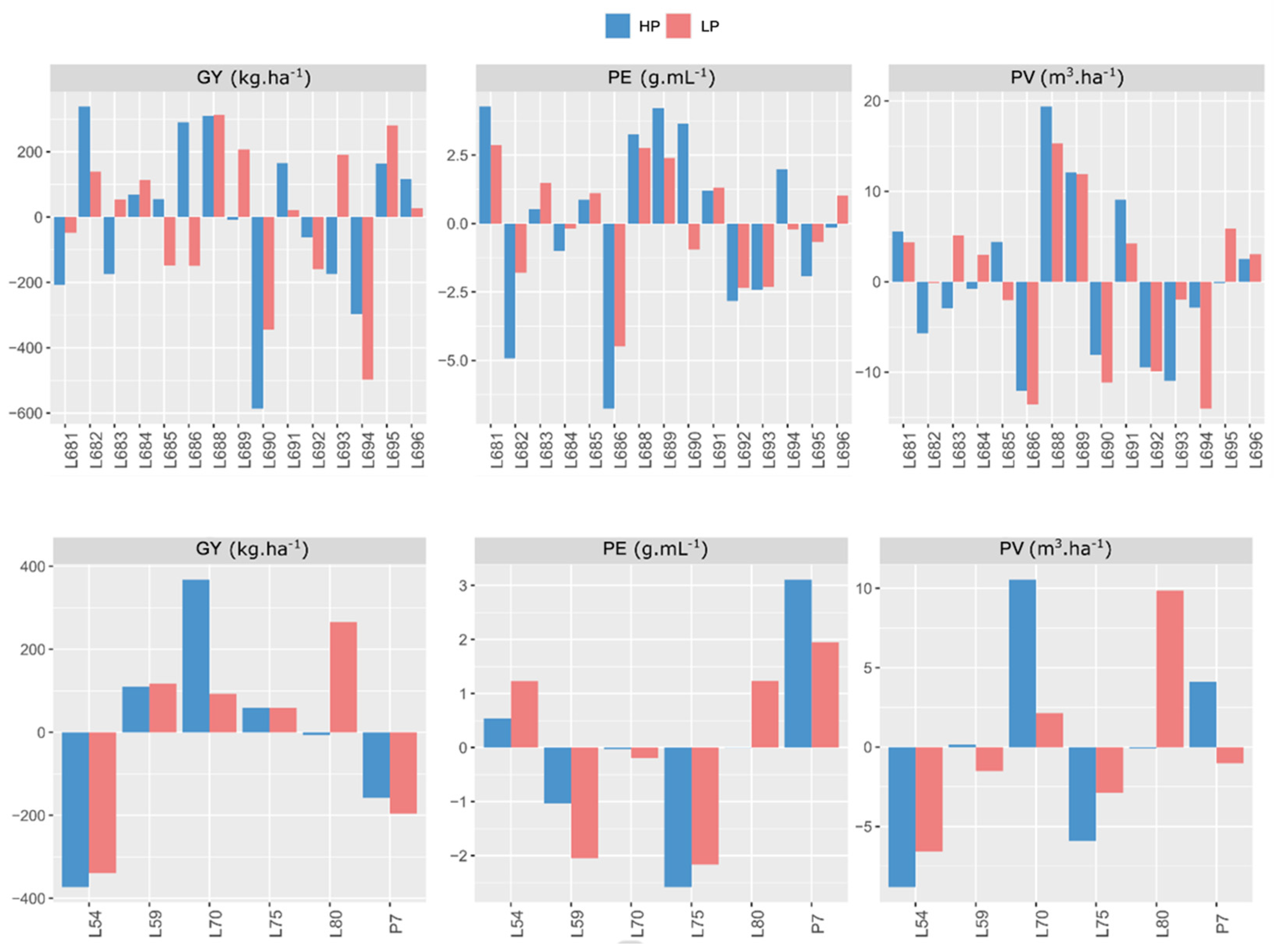
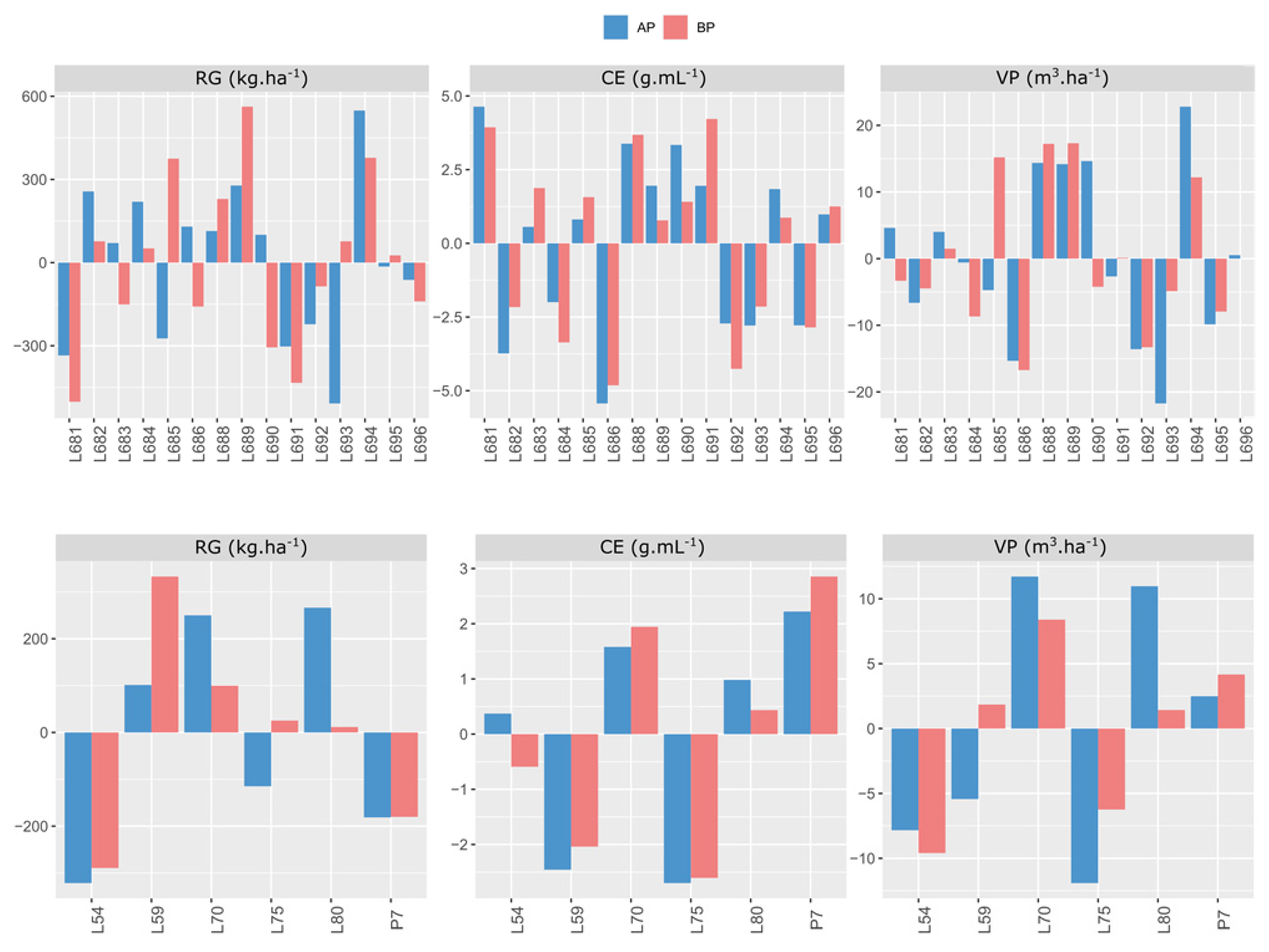
2.2. General Combining Ability Effects
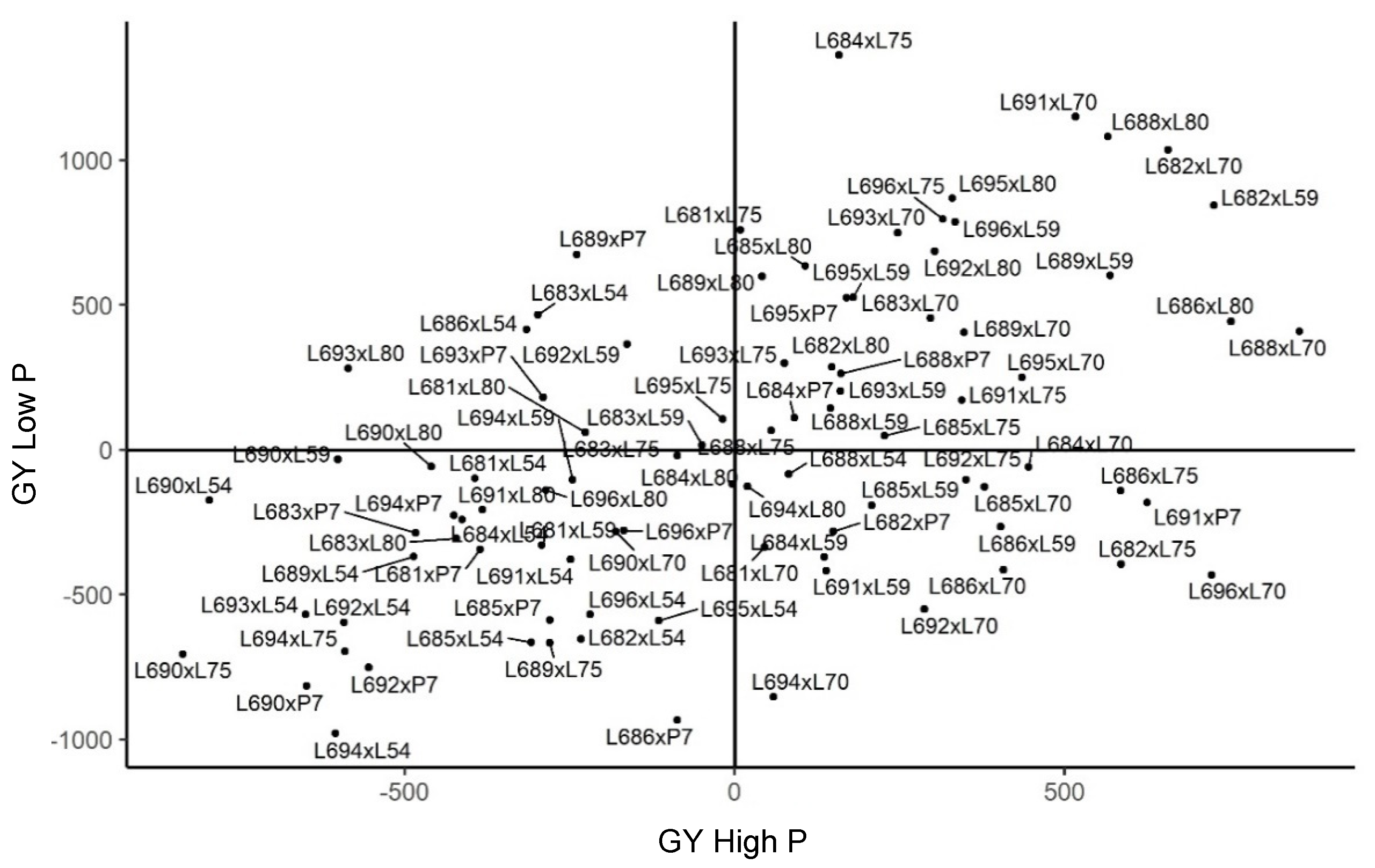
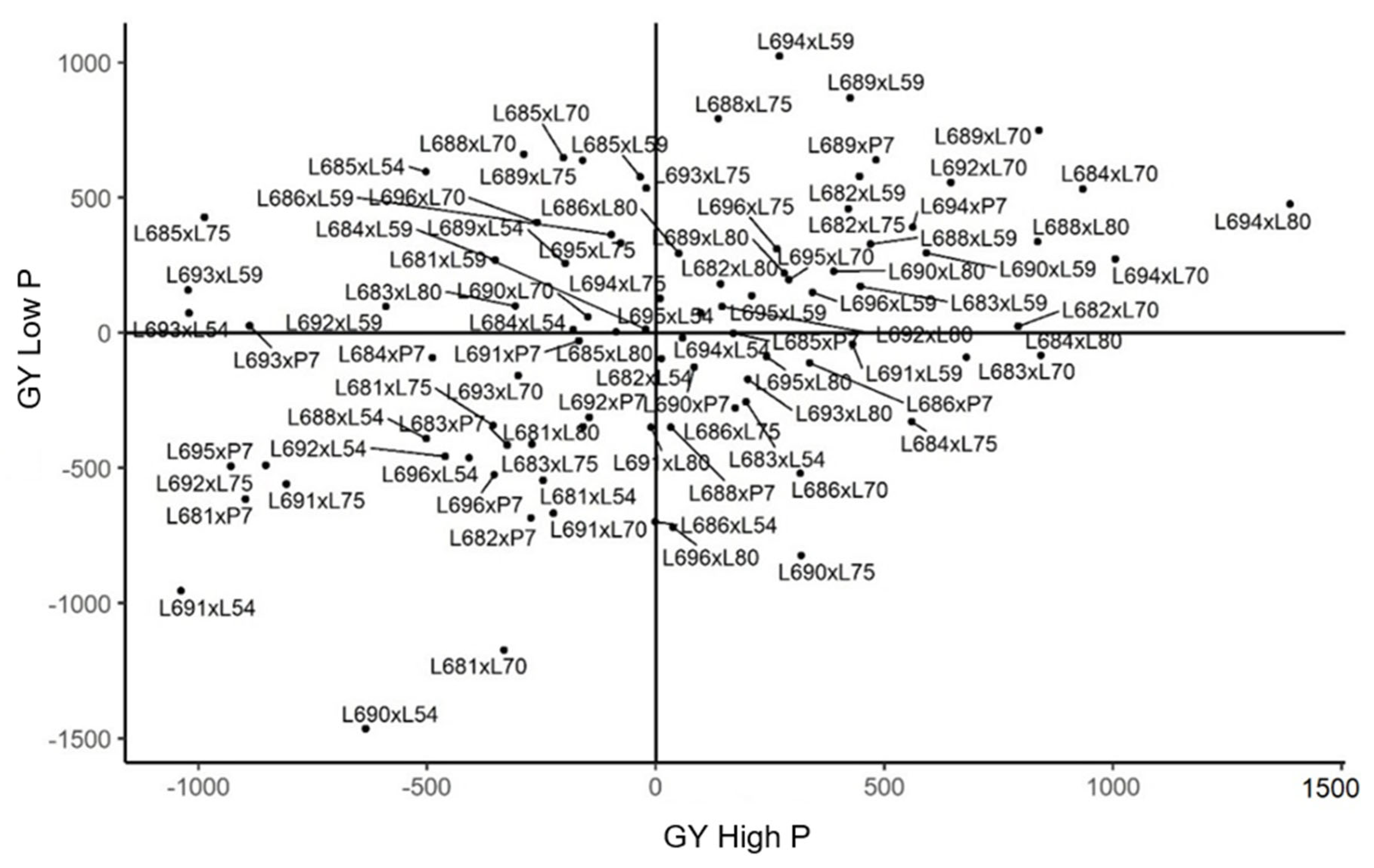
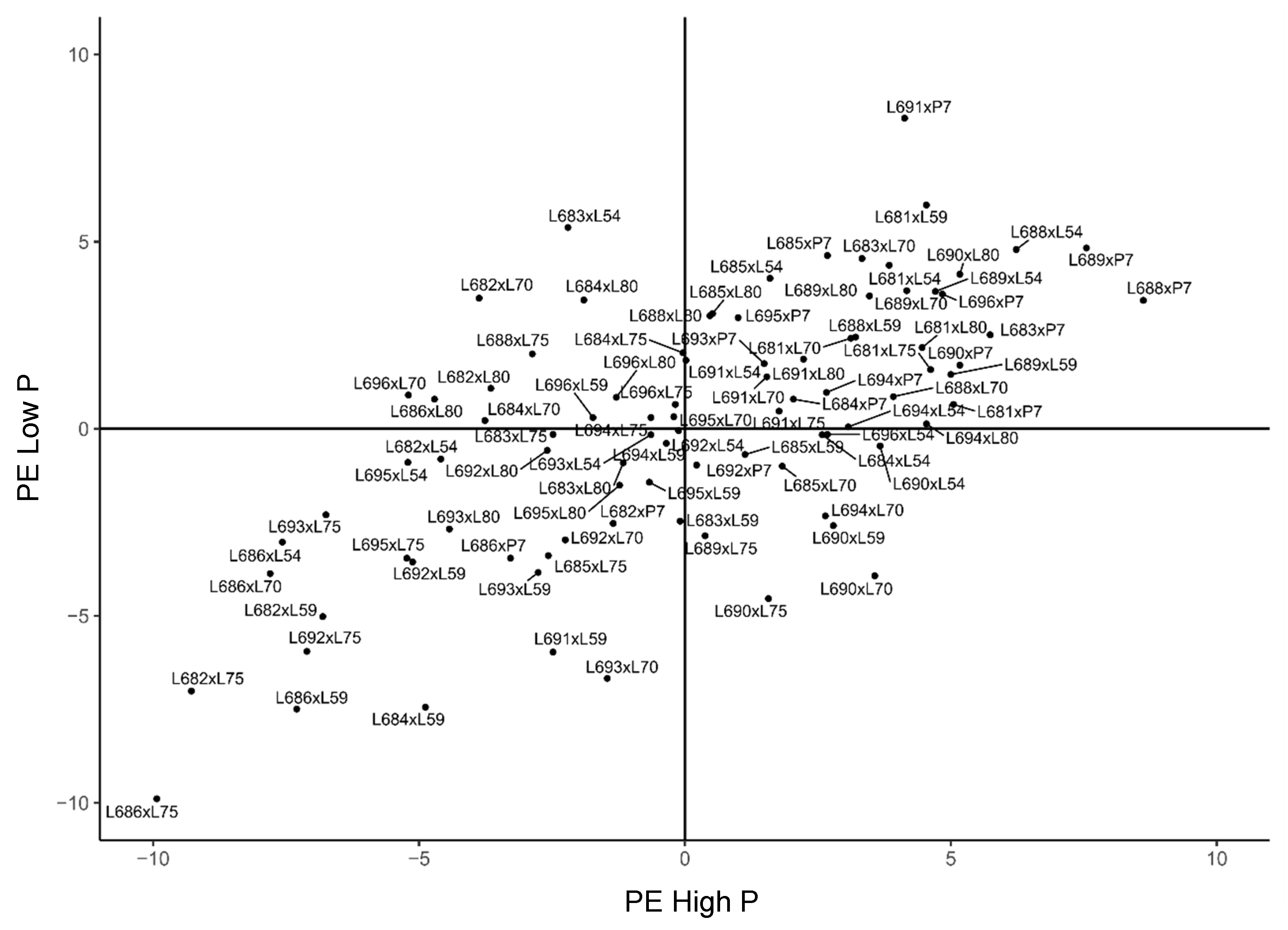
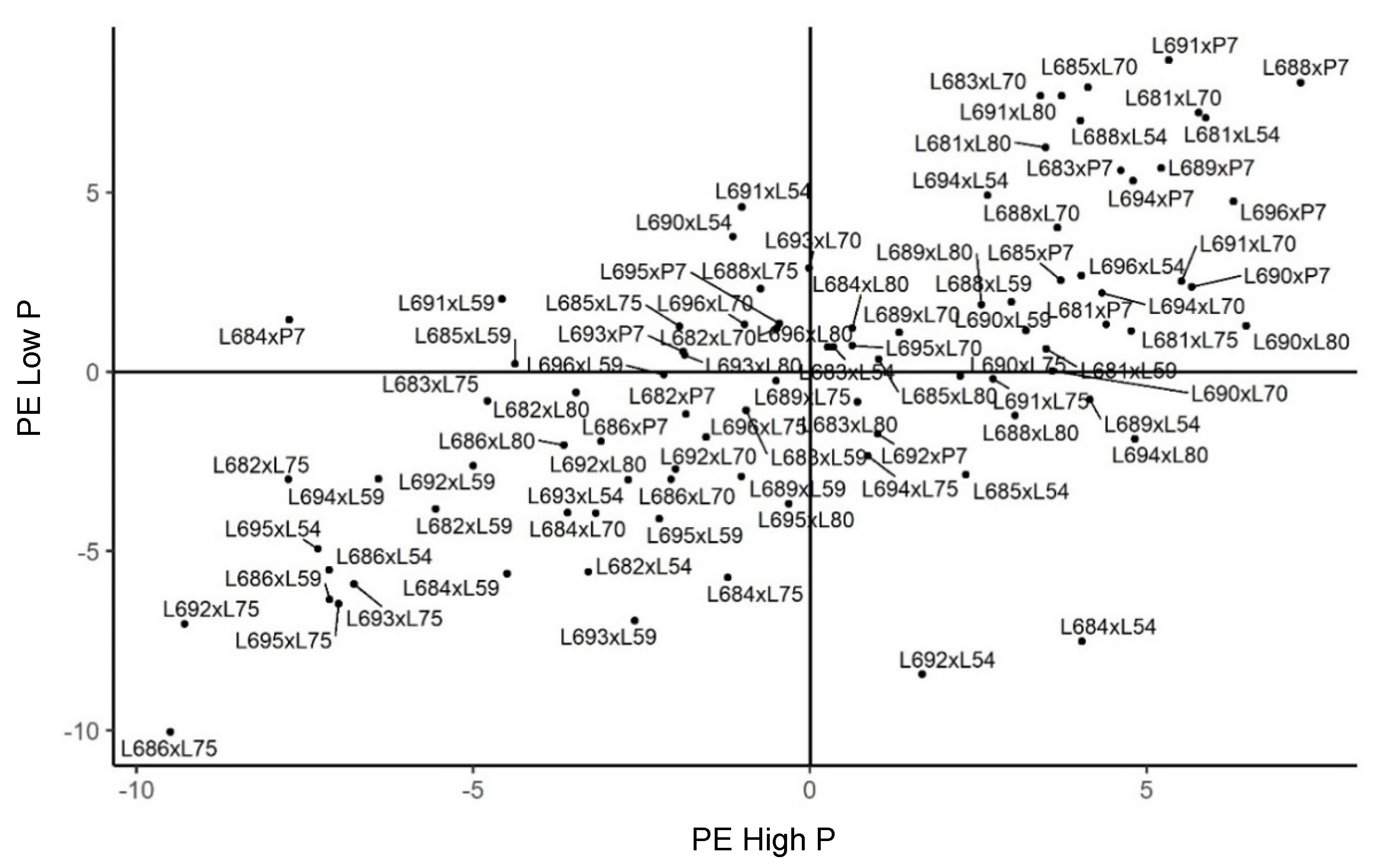
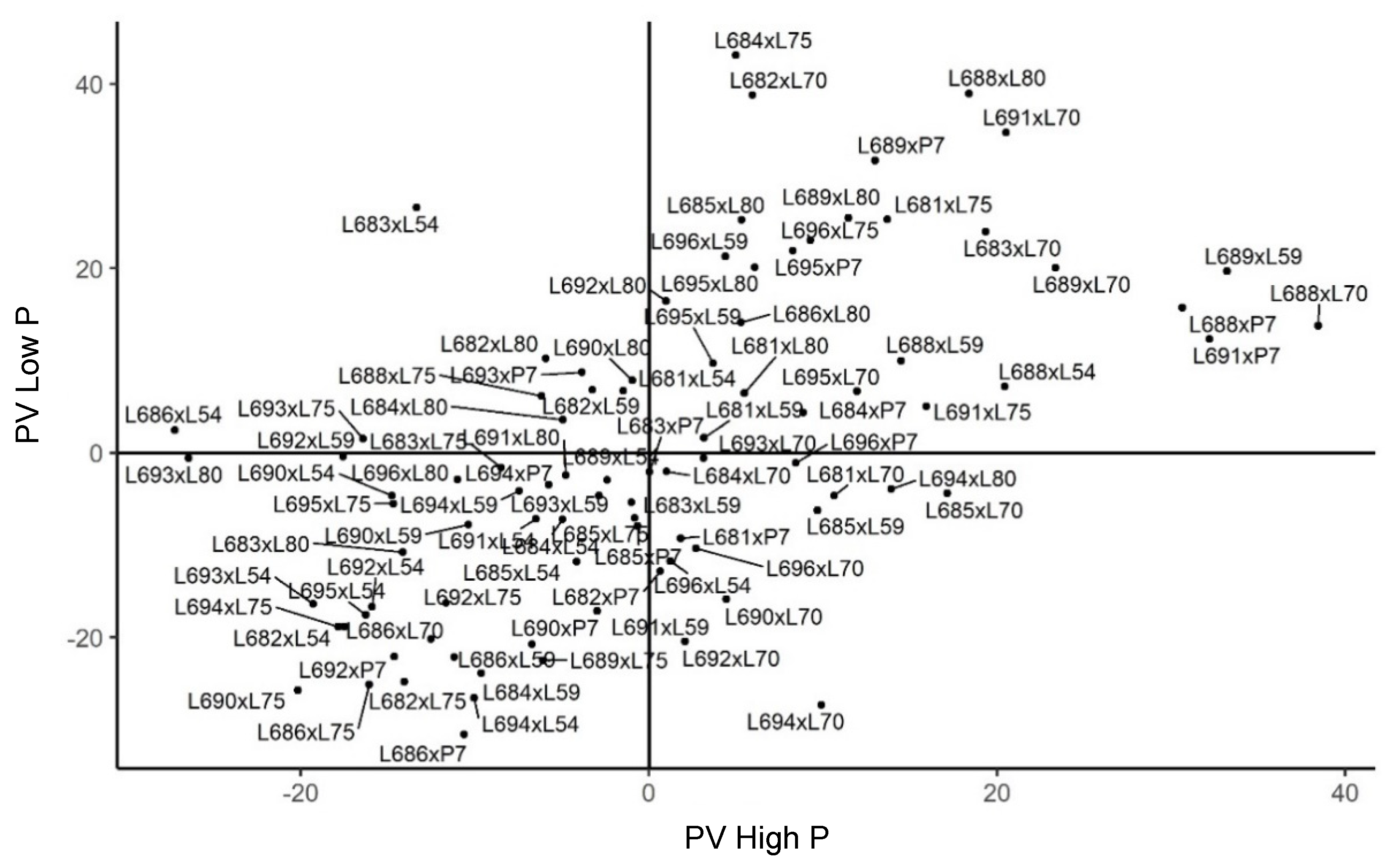
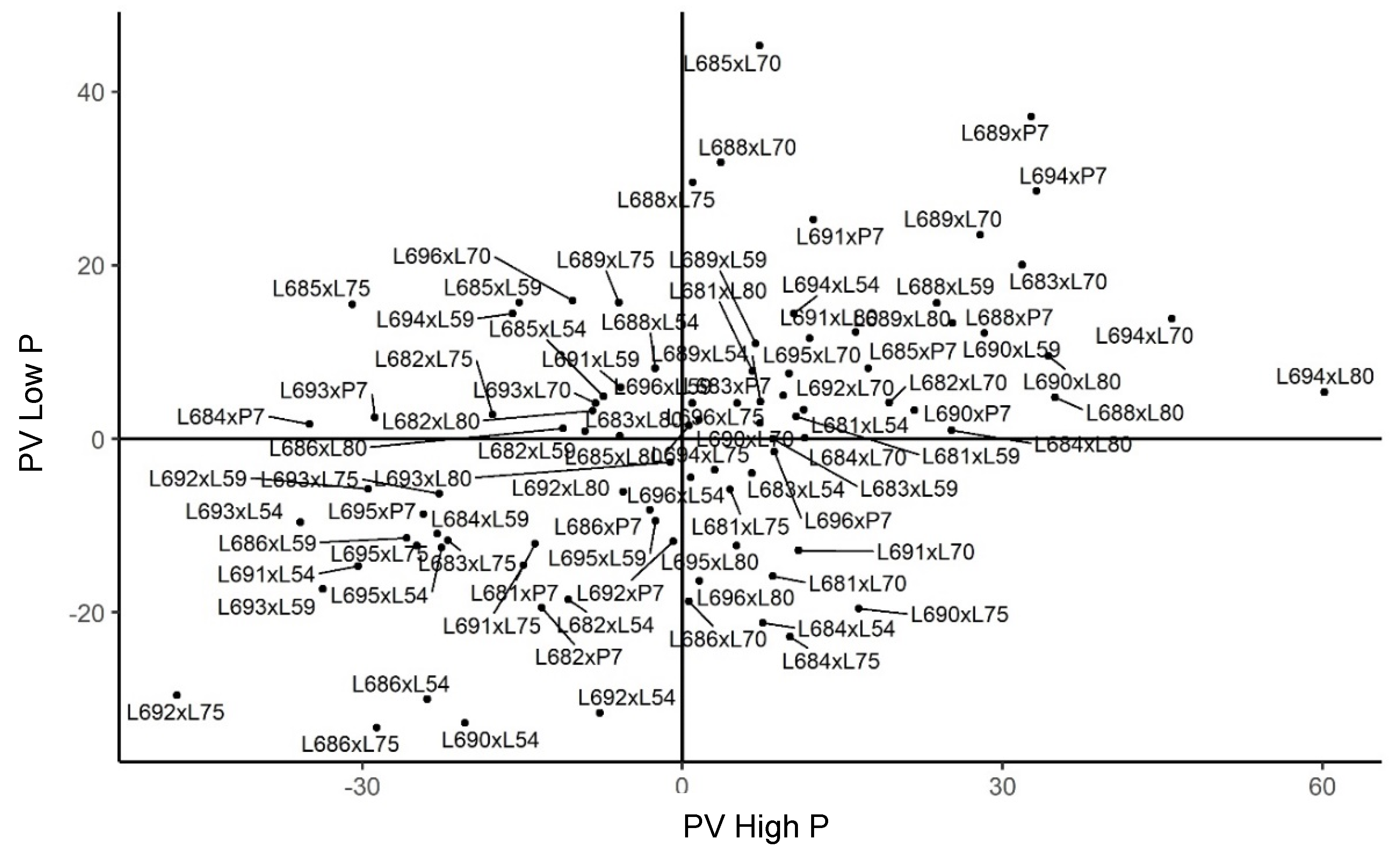
2.3. Genetic Merit of Hybrids
3. Discussion
4. Materials and Methods
4.1. Plant Material, Experimental Design, and Environmental Conditions
4.2. Evaluated Traits
4.3. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zeng, Q.; Mei, T.; Delgado-Baquerizo, M.; Wang, M.; Tan, W. Suppressed phosphorus-mineralizing bacteria after three decades of fertilization. Agric. Ecosyst. Environ. 2021, 323, 107679. [Google Scholar] [CrossRef]
- Collavino, M.M.; Sansberro, P.A.; Mroginski, L.A.; Aguilar, O.M. Comparison of in vitro solubilization activity of diverse phosphate-solubilizing bacteria native to acid soil and their ability to promote Phaseolus vulgaris growth. Biol. Fertil. Soils 2010, 46, 727–738. [Google Scholar] [CrossRef]
- Wahid, F.; Sharif, M.; Fahad, S.; Ali, A.; Adnan, M.; Rafiullah; Saud, S.; Danish, S.; Ali, M.A.; Ahmed, N.; et al. Mycorrhiza and Phosphate Solubilizing Bacteria: Potential Bioagents for Sustainable Phosphorus Management in Agriculture. Phyton 2022, 92, 257–278. [Google Scholar] [CrossRef]
- Hickey, L.T.; Hafeez, A.N.; Robinson, H.; Jackson, S.A.; Leal-Bertioli, S.C.M.; Tester, M.; Gao, C.; Godwin, I.D.; Hayes, B.J.; Wulff, B.B.H. Breeding crops to feed 10 billion. Nat. Biotechnol. 2019, 37, 744–754. [Google Scholar] [CrossRef]
- Duboc, O.; Hernandez-Mora, A.; Wenzel, W.W.; Santner, J. Improving the prediction of fertilizer phosphorus availability to plants with simple, but non-standardized extraction techniques. Sci. Total Environ. 2021, 806, 150486. [Google Scholar] [CrossRef]
- López-Arredondo, D.L.; Leyva-González, M.A.; González-Morales, S.I.; López-Bucio, J.; Herrera-Estrella, L. Phosphate nutrition: Improving low-phosphate tolerance in crops. Annu. Rev. Plant Biol. 2014, 65, 95–123. [Google Scholar] [CrossRef]
- Barros, V.M.D.S.; Martins, L.D.; Rodrigues, W.N.; Ferreira, D.S.; Christo, B.F.; do Amaral, J.F.T.; Tomaz, M.A. Combined doses of nitrogen and phosphorus in conilon coffee plants: Changes in absorption, translocation and use in plant compartments. J. Plant Nutr. 2021, 45, 346–357. [Google Scholar] [CrossRef]
- Lee, J.-I.; Kim, J.-M.; Yoo, S.-C.; Jho, E.H.; Lee, C.-G.; Park, S.-J. Restoring phosphorus from water to soil: Using calcined eggshells for P adsorption and subsequent application of the adsorbent as a P fertilizer. Chemosphere 2021, 287, 132267. [Google Scholar] [CrossRef]
- Meng, X.; Chen, W.-W.; Wang, Y.-Y.; Huang, Z.-R.; Ye, X.; Chen, L.-S.; Yang, L.-T. Effects of phosphorus deficiency on the absorption of mineral nutrients, photosynthetic system performance and antioxidant metabolism in Citrus grandis. PLoS ONE 2021, 16, e0246944. [Google Scholar] [CrossRef]
- Gerhardt, I.F.S.; do Amaral, A.T., Jr.; Pena, G.F.; Guimarães, L.J.M.; de Lima, V.J.; Vivas, M.; Santos, P.H.A.D.; Ferreira, F.R.A.; Freitas, M.S.M.; Kamphorst, S.H. Genetic effects on the efficiency and responsiveness to phosphorus use in popcorn as estimated by diallel analysis. PLoS ONE 2019, 14, e0216980. [Google Scholar] [CrossRef]
- Cruz, C.D.; Carneiro, P.C.S.; Regazzi, A.J. Modelos Biométricos Aplicados ao Melhoramento Genético, 3rd ed.; Editora UFV: Viçosa, Brazil, 2014; p. 668. [Google Scholar]
- Hallauer, A.R.; Carena, M.J.; Miranda Filho, J.D. Quantitative Genetics in Maize Breeding; Springer: New York, NY, USA, 2010; p. 664. [Google Scholar]
- Davis, R.L. Report of the plant breeder. In Rio de Piedras: Agricultural Experiment Station University; Annual Report; Agricultural Experiment Station University: Davis, CA, USA, 1927; pp. 14–15. [Google Scholar]
- De Lima, V.J.; do Amaral, A.T., Jr.; Kamphorst, S.H.; Bispo, R.B.; Leite, J.T.; de Oliveira Santos, T.; Schmitt, K.F.M.; Chaves, M.M.; de Oliveira, U.A.; Santos, P.H.A.D.; et al. Combined Dominance and Additive Gene Effects in Trait Inheritance of Drought-Stressed and Full Irrigated Popcorn. Agronomy 2019, 9, 782. [Google Scholar] [CrossRef]
- Pereira, M.G.; do Amaral, A.T., Jr. Estimation of Genetic Components in Popcorn Based on the Nested Design. Crop Breed. Appl. Biotechnol. 2001, 1, 3–10. [Google Scholar] [CrossRef]
- Daros, Z.A.; Lone, A.A.; Alie, B.A.; Ahangar, M.A.; Ali, G.; Abidi, I.; Gazal, A.; Lone, R.A. Combining ability analysis for yield and yield contributing traits in Popcorn (Zea mays everta L.) under temperate conditions. J. Pharm. Phytochem. 2018, 7, 361–366. [Google Scholar]
- Schwantes, I.A.; do Amaral, A.T., Jr.; Filho, J.E.A.; Vivas, M.; Cabral, P.D.S.; Guimarães, A.G.; Silva, F.H.L.; Santos, P.H.A.D.; Pereira, M.G.; Viana, A.P.; et al. Genomic selection helps accelerate popcorn population breeding. Crop Sci. 2020, 60, 1373–1385. [Google Scholar] [CrossRef]
- Kamphorst, S.H.; do Amaral, A.T., Jr.; de Lima, V.J.; Carena, M.J.; Azeredo, V.C.; Mafra, G.S.; Santos, P.H.A.D.; Leite, J.T.; Schmitt, K.F.M.; dos Santos, D.R., Jr.; et al. Driving Sustainable Popcorn Breeding for Drought Tolerance in Brazil. Front. Plant Sci. 2021, 12, 732285. [Google Scholar] [CrossRef]
- Kamphorst, S.H.; do Amaral, A.T., Jr.; Vergara-Diaz, O.; Gracia-Romero, A.; Fernandez-Gallego, J.A.; Chang-Espino, M.C.; Buchaillot, M.L.; Rezzouk, F.Z.; Lima, V.J.; Serret, M.; et al. Heterosis and reciprocal effects for physiological and morphological traits of popcorn plants under different water conditions. Agric. Water Manag. 2022, 261, 107371. [Google Scholar] [CrossRef]
- Dos Santos, A.; do Amaral, A.T., Jr.; Fritsche-Neto, R.; Kamphorst, S.H.; Ferreira, F.R.A.; Amaral, J.F.T.; Vivas, J.M.S.; Santos, P.H.A.D.; de Lima, V.J.; Khan, S.; et al. Relative importance of gene effects for nitrogen-use efficiency in popcorn. PLoS ONE 2019, 14, e0222726. [Google Scholar] [CrossRef]
- Dos Santos, A.; do Amaral, A.T., Jr.; Kamphorst, S.H.; Gonçalves, G.M.B.; Santos, P.H.A.D.; Vivas, J.M.S.; Mafra, G.S.; Khan, S.; de Oliveira, F.T.; Schmitt, K.F.M.; et al. Evaluation of Popcorn Hybrids for Nitrogen Use Efficiency and Responsiveness. Agronomy 2020, 10, 485. [Google Scholar] [CrossRef]
- Schwantes, I.A.; do Amaral, A.T., Jr.; Gerhardt, I.F.S.; Vivas, M.; Silva, F.H.D.L.E.; Kamphorst, S.H. Diallel analysis of resistance to Fusarium ear rot in Brazilian popcorn genotypes. Trop. Plant Pathol. 2017, 42, 70–75. [Google Scholar] [CrossRef]
- Mafra, G.S.; do Amaral, A.T., Jr.; Vivas, M.; Santos, J.S.D.; Guimarães, A.G.; Pena, G.F. The combining ability of popcorn S7 lines for Puccinia polysora resistance purposes. Bragantia 2018, 77, 519–526. [Google Scholar] [CrossRef]
- Do Amaral, A.T., Jr.; do Poltronieri, T.P.S.; dos Santos, P.H.D.; Vivas, M.; Gerhardt, I.F.S.; Carvalho, B.M.; Freitas, C.S.; Silveira, S.F. Reaction of popcorn lines (S7) cultivated in distinct phosphorus levels to Bipolaris maydis infection. Summa Phytopathol. 2019, 45, 18–22. [Google Scholar] [CrossRef]
- Kurosawa, R.D.N.F.; do Amaral, A.T., Jr.; Vivas, J.M.S.; Vivas, M.; Kamphorst, S.H.; de Lima, V.J.; de Almeida, R.N. Selection of popcorn hybrids resistant to southern corn leaf blight grown in distinct N availability. Eur. J. Plant Pathol. 2020, 158, 485–493. [Google Scholar] [CrossRef]
- Kurosawa, R.D.N.F.; do Amaral, A.T., Jr.; do Vivas, M.; de Almeida, R.N.; Vivas, J.M.S.; de Lima, V.J.; da Silveira, S.F. Diallel analysis for resistance to northern leaf blight in popcorn under contrasting nitrogen availability. Agron. J. 2021, 113, 1029–1038. [Google Scholar] [CrossRef]
- Calderón-Vázquez, C.L.; Sawers, R.J.; Herrera-Estrella, L. Phosphate Deprivation in Maize: Genetics and Genomics. Plant Physiol. 2011, 156, 1067–1077. [Google Scholar] [CrossRef] [PubMed]
- Vencovsky, R.; Barriga, P. Genética Biométrica no Fitomelhoramento; Sociedade Brasileira de Genética: Ribeirao Preto, Brazil, 1992; p. 496. [Google Scholar]
- Fritsche-Neto, R.; Miranda, G.V.; de Lima, R.O.; de Souza, L.V.; da Silva, J. Herança de caracteres associados à eficiência de utilização do fósforo em milho. Pesq. Agropec. Bras. 2010, 45, 465–471. [Google Scholar] [CrossRef]
- Dofing, S.M.; D’Croz-Mason, N.; Thomas-Compton, M.A. Inheritance of Expansion Volume and Yield in Two Popcorn × Dent Corn Crosses. Crop Sci. 1991, 31, 715–718. [Google Scholar] [CrossRef]
- Pacheco, C.A.; Meirelles, W.F.; Magalhães, J.V.; Guimarães, L.J.M.; Silva, A.R.; Ferreira Mendes, F.; Schaffert, R.E. Inheritanceand Breeding strategies for phosphorus efficiency in tropical maize (Zea mays L.). Maydica 2010, 55, 1–15. [Google Scholar]
- Larish, L.L.B.; Brewbaker, J.L. Diallel analyses of temperate and tropical popcorns. Maydica 1999, 44, 279–284. [Google Scholar]
- Coan, M.M.D.; Barth, R.J.P.; Kuki, M.C.; do Amaral, A.T.D., Jr.; Figueiredo, A.S.T.; Scapim, C.A.; Warburton, M. Inheritance study for popping expansion in popcorn vs. flint corn genotypes. Agron. J. 2019, 111, 1–10. [Google Scholar] [CrossRef]
- Scapim, C.A.; Carvalho, C.G.P.; Cruz, C.D. Uma proposta de classificação dos coeficientes de variação para a cultura do milho. Pesq. Agropec. Bras. 1995, 30, 683–686. [Google Scholar]
- Dos Santos, A.; do Amaral, A.T.D., Jr.; Kurosawa, R.D.N.F.; Gerhardt, I.F.S.; Neto, R.F. GGE Biplot projection in discriminating the efficiency of popcorn lines to use nitrogen. Ciência e Agrotecnologia 2017, 41, 22–31. [Google Scholar] [CrossRef]
- Sprague, G.F.; Tatum, L.A. General vs. Specific combining ability in single crosses of corn. J. Am. Soc. Agron. 1942, 34, 923–932. [Google Scholar] [CrossRef]
- Inocente, G.; Garbuglio, D.D.; Ruas, P.M. Multilayer perceptron applied to genotypes classification in diallel studies. Sci. Agricola 2022, 79, e20200365. [Google Scholar] [CrossRef]
- Cruz, C.D.; Vencovsky, R. Comparação de alguns métodos de análise dialélica. Rev. Bras. Genét. 1989, 12, 425–438. [Google Scholar]
- Scapim, C.A.; Pacheco, C.A.P.; Tonet, A.; Braccini, A.D.L.E.; Pinto, R.J.B. Análise Dialélica e Heterose de Populações de Milho-Pipoca. Bragantia 2002, 61, 219–230. [Google Scholar] [CrossRef]
- Kamphorst, S.H.; de Lima, V.; do Amaral, A.T.D., Jr.; Schmitt, K.F.M.; Leite, J.T.; Carvalho, C.M.; Silva, R.R.M.; Xavier, K.B.; Ferreira, F.R.A.; Santos, P.H.A.D.; et al. Popcorn breeding for water-stress tolerance or for agronomic water-use efficiency? Genet. Mol. Res. 2018, 17, 1–18. [Google Scholar] [CrossRef]
- Do Amaral, A.T.D., Jr.; dos Santos, A.; Gerhardt, I.F.S.; Kurosawa, R.N.F.; Moreira, N.F.; Pereira, M.G.; Gravina, G.D.A.; Silva, F.D.L. Proposal of a super trait for the optimum selection of popcorn progenies based on path analysis. Genet. Mol. Res. 2016, 15, 1–9. [Google Scholar] [CrossRef]
- Schmitt, K.F.M.; Lima, V.J.; do Amaral, A.T.D., Jr.; Santos, J.S.; Mafra, G.S.; Vivas, M.; Kamphorst, S.H.; Souza, Y.P.; Oliveira, F.T.; Ferreira, F.R.A.; et al. Combining ability of popcorn lines for resistance to the fungus Puccinia polysora (Pucciniaceae). Genet. Mol. Res. 2019, 18, 1–11. [Google Scholar] [CrossRef]
- Santos, T.D.O.; do Amaral, A.T.D., Jr.; Bispo, R.B.; de Lima, V.J.; Kamphorst, S.H.; Leite, J.T.; dos Santos Júnior, D.R.; Santos, P.H.A.D.; de Oliveira, U.A.; Schmitt, K.F.M.; et al. Phenotyping Latin American Open-Pollinated Varieties of Popcorn for Environments with Low Water Availability. Plants 2021, 10, 1211. [Google Scholar] [CrossRef]
- Willier, G. Factors affecting the popping quality of popcorn. J. Agric. Res. 1927, 35, 615–624. [Google Scholar]
- Hallauer, A.R. Specialty Corns, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2001; p. 496. [Google Scholar]
- Daros, M.; do Amaral, A.T.D., Jr.; Pereira, M.G.; Santos, F.S.; Gabriel, A.P.C.; Scapim, C.A.; Silvério, D.P.F., Jr. Recurrent selection in inbred popcorn families. Sci. Agricola 2004, 61, 609–614. [Google Scholar] [CrossRef]
- Rangel, R.M.; do Amaral, A.T.D., Jr.; Gonçalves, L.S.A.; Freitas, S.P., Jr. Análise biométrica de ganhos por seleção em população de milho-pipoca de quinto ciclo de seleção recorrente. Rev. Ciência Agronômica 2011, 42, 473–481. [Google Scholar] [CrossRef]
- Zinsly, J.R.; Machado, J.A. Milho-pipoca. In Melhoramento e Produção do Milho, 2nd ed.; Paterniani, E., Viégas, G.P., Eds.; Fundação Cargill: Campinas, Brazil, 1987; pp. 413–421. [Google Scholar]
- De Andrade, R.A.; Cruz, C.D.; Scapim, C.A.; Silverio, L.; Pinto, R.J.B.; Tonet, A. Análise dialélica da capacidade combinatória de variedades de milho-pipoca. Acta Scientiarum. Agron. 2008, 24, 1197–1204. [Google Scholar] [CrossRef]
- Gerhardt, I.F.S.; do Amaral, A.T.D., Jr.; Guimarães, L.J.M.; Schwantes, I.A.; Santos, A.; Kamphorst, S.H.; de Lima, V.J.; Poblete, F.M.; Myers, G.O. Intraspecific variability of popcorn S7 lines for phosphorus efficiency in the soil. Genet. Mol. Res. 2017, 16, gmr16039763. [Google Scholar] [CrossRef] [PubMed]
- Do Amaral, A.T.D., Jr.; Goncalves, L.S.A.; Freitas, S.D.P., Jr.; Candido, L.S.; Vittorazzi, C.; Pena, G.F.; Ribeiro, R.M.; Silva, T.R.D.C.; Pereira, M.G.; Scapim, C.A.; et al. UENF 14: A new popcorn cultivar. Crop Breed. Appl. Biotechnol. 2013, 13, 218–220. [Google Scholar] [CrossRef]
- Ribeiro, A.C.; Guimarães, P.T.G.; Alvarez, V.H.A. Recomendações Para o Uso de Corretivos e Fertilizantes em Minas Gerais: 5° Aproximação; Comissão de Fertilidade do Solo do Estado de Minas Gerais: Viçosa, Brazil, 1999; p. 195. [Google Scholar]
- Vitorazzi, C.; do Amaral, A.T.D., Jr.; Candido, L.S.; Freitas, I.L.J.; Silva, T.R.C. Arranjo populacional para a variedade UENF-14 de milho-pipoca. Rev. Bras. Milho Sorgo 2017, 16, 401–413. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, R.M.; do Amaral, A.T.D., Jr.; Pena, G.F.; Vivas, M.; Kurosawa, R.F.N.; Goncalves, L.S.A. Effect of recurrent selection on the variability of the UENF-14 popcorn population. Crop Breed. Appl. Biotechnol. 2016, 16, 123–131. [Google Scholar] [CrossRef]
- Guimarães, A.G.; do Amaral, A.T.D., Jr.; Pena, G.F.; Filho, J.E.A.; Pereira, M.G.; Santos, P.H.A.D. Genetic gains in the popcorn population UENF-14: Developing the ninth generation of intrapopulation recurrent selection. Rev. Caatinga 2019, 32, 625–633. [Google Scholar] [CrossRef]
- Griffing, B. Concept of general and specific Combining ability in relation to diallel crossing systems. Aust. J. Biol. Sci. 1956, 9, 463–493. [Google Scholar] [CrossRef]
- Cruz, C.D. GENES—A software package for analysis in experimental statistics and quantitative genetics. Acta Sci. Agron. 2013, 35, 271–276. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SV | DF | Mean Squares | |||||
|---|---|---|---|---|---|---|---|
| Campos dos Goytacazes | Itaocara | ||||||
| GY | PE | PV | GY | PE | PV | ||
| Block/Phosphorus | 4 | 554,624.5 | 9.5 | 529.0 | 103,022.6 | 3.3 | 116.5 |
| Genotype (G) | 89 | 954,080.6 ** | 69.6 ** | 1101.1 ** | 992,731.8 ** | 87.40 ** | 1414.1 ** |
| GCA I | 14 | 1,672,189.3 ** | 247.9 ** | 2585.7 ** | 2,244,297.7 * | 314.3 ** | 4149.6 ** |
| GCA II | 5 | 4,395,517.2 * | 279.9 ** | 2618.4 ns | 4,047,292.2 * | 398.7 ** | 5616.3 * |
| SCA | 70 | 564,640.9 ** | 18.9 ** | 695.8 ** | 524,233.5 * | 19.8 ns | 566.9 ns |
| Phosphorus (P) | 1 | 36,456,840.8 ** | 246.3 ** | 37,105.2 ** | 17,515,196.9 ** | 255.7 ** | 27,642.8 ** |
| G × P | 89 | 330,925.0 ** | 14.6 ** | 317.0 ** | 423,063.8 ** | 16.7 ** | 509.3 ** |
| GCA I × P | 14 | 439,193.5 ** | 33.0 ** | 287.6 ns | 423,063.9 ** | 13.9 ** | 848.3 ** |
| GCA II × P | 5 | 684,529.0 ** | 20.6 * | 956.3 ** | 845,321.8 ** | 8.8 ** | 868.8 ** |
| SCA × P | 70 | 284,013.2 ** | 10.5 ** | 277.2 ** | 727,343.1 ** | 17.8 ** | 415.9 ** |
| Residue | 356 | 178,327.5 | 7.0 | 179.3 | 209,610.6 | 2.8 | 171.1 |
| Mean | 2514.8 | 27.6 | 69.6 | 3143.7 | 26.2 | 82.4 | |
| CVe (%) | 16.8 | 9.6 | 19.2 | 14.6 | 6.4 | 15.9 | |
| Tester | Genealogy | Climate Adaptation | Classification Regarding P Use | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| L59 | Beija-Flor: UFV | Temperate/Tropical | Efficient and Responsive | ||||||||
| L70 | BRS Angela: EMBRAPA | Tropical | Efficient and Responsive | ||||||||
| P7 | Híbrido Zaeli | Temperate/tropical | Efficient and responsive | ||||||||
| L54 | Beija-flor: UFV | Temperate/tropical | Inefficient and non-responsive | ||||||||
| L75 | Viçosa: UFV | Temperate/tropical | Inefficient and non-responsive | ||||||||
| L80 | Viçosa: UFV | Temperate/tropical | Inefficient and non-responsive | ||||||||
| IRS Progenies | Genealogy and Climate Adaptation | IRS Progenies | Genealogy and Climate Adaptation | IRS Progenies | Genealogy and Climate Adaptation | ||||||
| 1 | L681 | UENF-14 | Tropical | 6 | L686 | UENF-14 | Tropical | 11 | L692 | UENF-14 | Tropical |
| 2 | L682 | UENF-14 | Tropical | 7 | L688 | UENF-14 | Tropical | 12 | L693 | UENF-14 | Tropical |
| 3 | L683 | UENF-14 | Tropical | 8 | L689 | UENF-14 | Tropical | 13 | L694 | UENF-14 | Tropical |
| 4 | L684 | UENF-14 | Tropical | 9 | L690 | UENF-14 | Tropical | 14 | L695 | UENF-14 | Tropical |
| 5 | L685 | UENF-14 | Tropical | 10 | L691 | UENF-14 | Tropical | 15 | L696 | UENF-14 | Tropical |
| Local | Soil Layer | pH | P | K | Ca | Mg | Al | Na | C | OM | CEC | BS | V | Clay |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| H2O | mg/dm3 | mmolc/dm3 | g/dm3 | mmolc/dm3 | % | g/dm3 | ||||||||
| Campos dos Goytacazes | 0–10 cm | 5.7 | 4.0 | 3.4 | 14.6 | 8.3 | 0.0 | 1.3 | 7.6 | 13.1 | 48.9 | 27.6 | 56.0 | 305.0 |
| 10–20 cm | 5.4 | 3.0 | 2.3 | 14.4 | 6.7 | 0.9 | 1.2 | 7.2 | 12.4 | 44.5 | 24.6 | 55.0 | ||
| Itaocara | 0–10 cm | 5.2 | 5.0 | 1.2 | 38.9 | 30.0 | 1.2 | 0.6 | 14.9 | 25.7 | 103.8 | 70.7 | 68.0 | 140.0 |
| 10–20 cm | 5.3 | 3.0 | 0.7 | 36.2 | 31.0 | 1.4 | 0.4 | 11.0 | 19.0 | 96.2 | 68.3 | 71.0 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, T.d.O.; de Oliveira, F.T.; Amaral Junior, A.T.d.; de Almeida Filho, J.E.; Bispo, R.B.; de Freitas, M.S.M.; Amaral, J.F.T.d.; Kamphorst, S.H.; de Lima, V.J.; Viana, F.N.; et al. Additive and Non-Additive Effects on the Control of Key Agronomic Traits in Popcorn Lines under Contrasting Phosphorus Conditions. Plants 2022, 11, 2216. https://doi.org/10.3390/plants11172216
Santos TdO, de Oliveira FT, Amaral Junior ATd, de Almeida Filho JE, Bispo RB, de Freitas MSM, Amaral JFTd, Kamphorst SH, de Lima VJ, Viana FN, et al. Additive and Non-Additive Effects on the Control of Key Agronomic Traits in Popcorn Lines under Contrasting Phosphorus Conditions. Plants. 2022; 11(17):2216. https://doi.org/10.3390/plants11172216
Chicago/Turabian StyleSantos, Talles de Oliveira, Fábio Tomaz de Oliveira, Antônio Teixeira do Amaral Junior, Janeo Eustáquio de Almeida Filho, Rosimeire Barboza Bispo, Marta Simone Mendonça de Freitas, José Francisco Teixeira do Amaral, Samuel Henrique Kamphorst, Valter Jário de Lima, Flávia Nicácio Viana, and et al. 2022. "Additive and Non-Additive Effects on the Control of Key Agronomic Traits in Popcorn Lines under Contrasting Phosphorus Conditions" Plants 11, no. 17: 2216. https://doi.org/10.3390/plants11172216
APA StyleSantos, T. d. O., de Oliveira, F. T., Amaral Junior, A. T. d., de Almeida Filho, J. E., Bispo, R. B., de Freitas, M. S. M., Amaral, J. F. T. d., Kamphorst, S. H., de Lima, V. J., Viana, F. N., Pena, G. F., Santos, P. H. A. D., Bernado, W. d. P., Pereira, M. G., Oliveira, J. G. d., Bressan-Smith, R. E., & Trindade, R. d. S. (2022). Additive and Non-Additive Effects on the Control of Key Agronomic Traits in Popcorn Lines under Contrasting Phosphorus Conditions. Plants, 11(17), 2216. https://doi.org/10.3390/plants11172216