
Identification of QTL under Brassinosteroid-Combined Cold Treatment at Seedling Stage in Rice Using Genotyping-by-Sequencing (GBS)


Abstract
:1. Introduction
2. Results
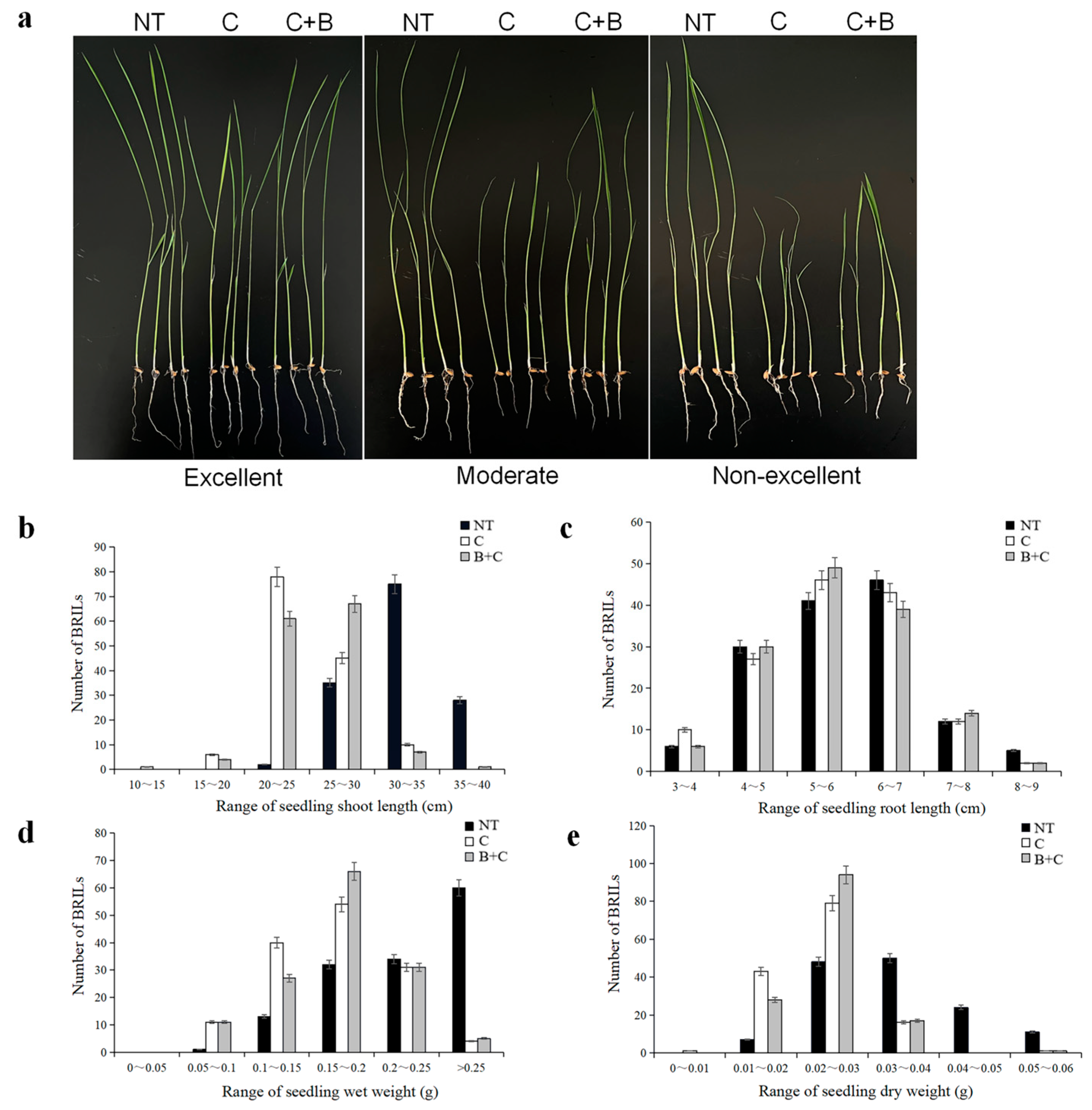
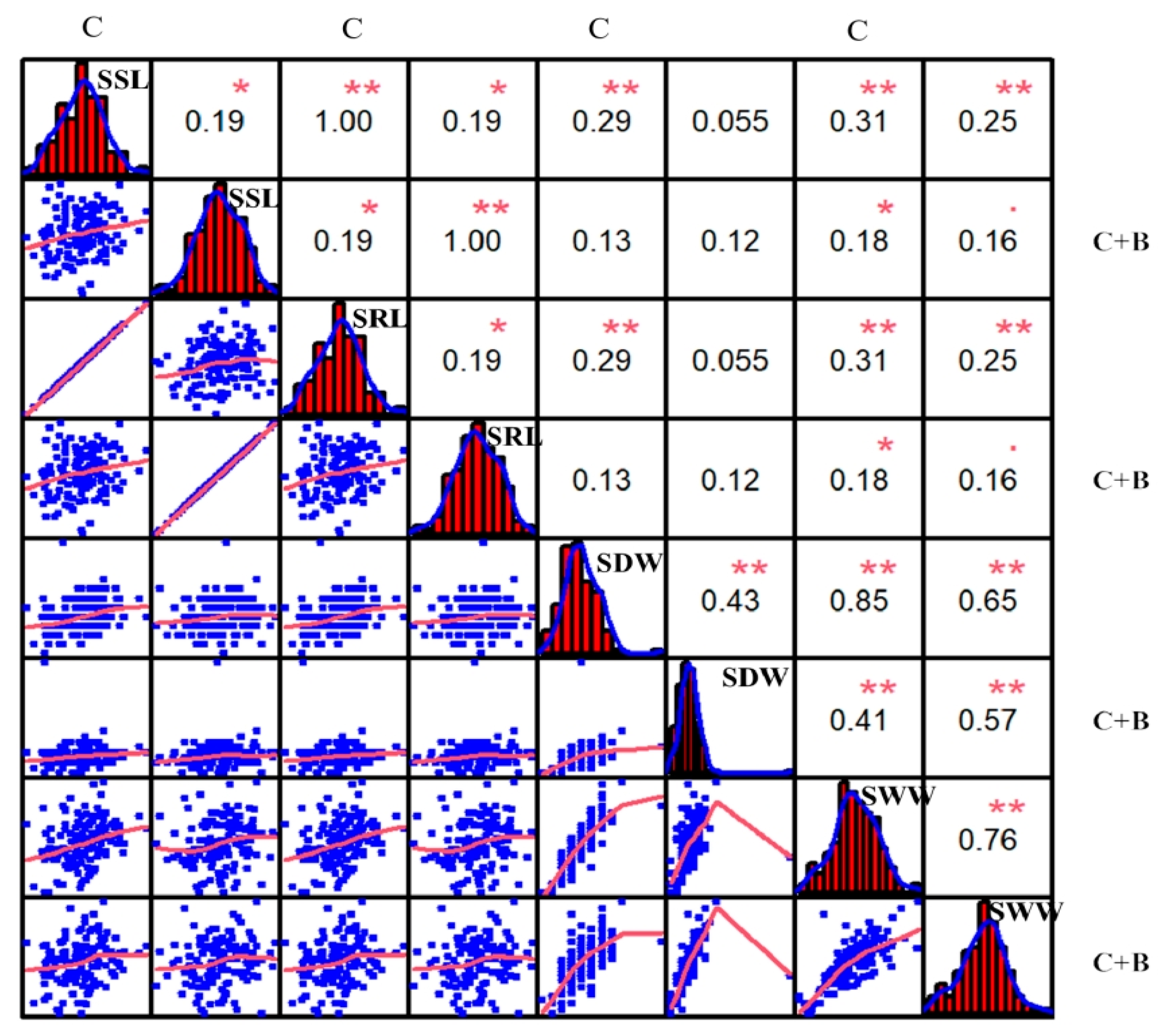
2.1. Phenotypic Characterization under Cold Stress and BR-Combined Cold Treatment
2.2. Construction of the Linkage Maps
2.3. Identification of QTLs for Cold Tolerance
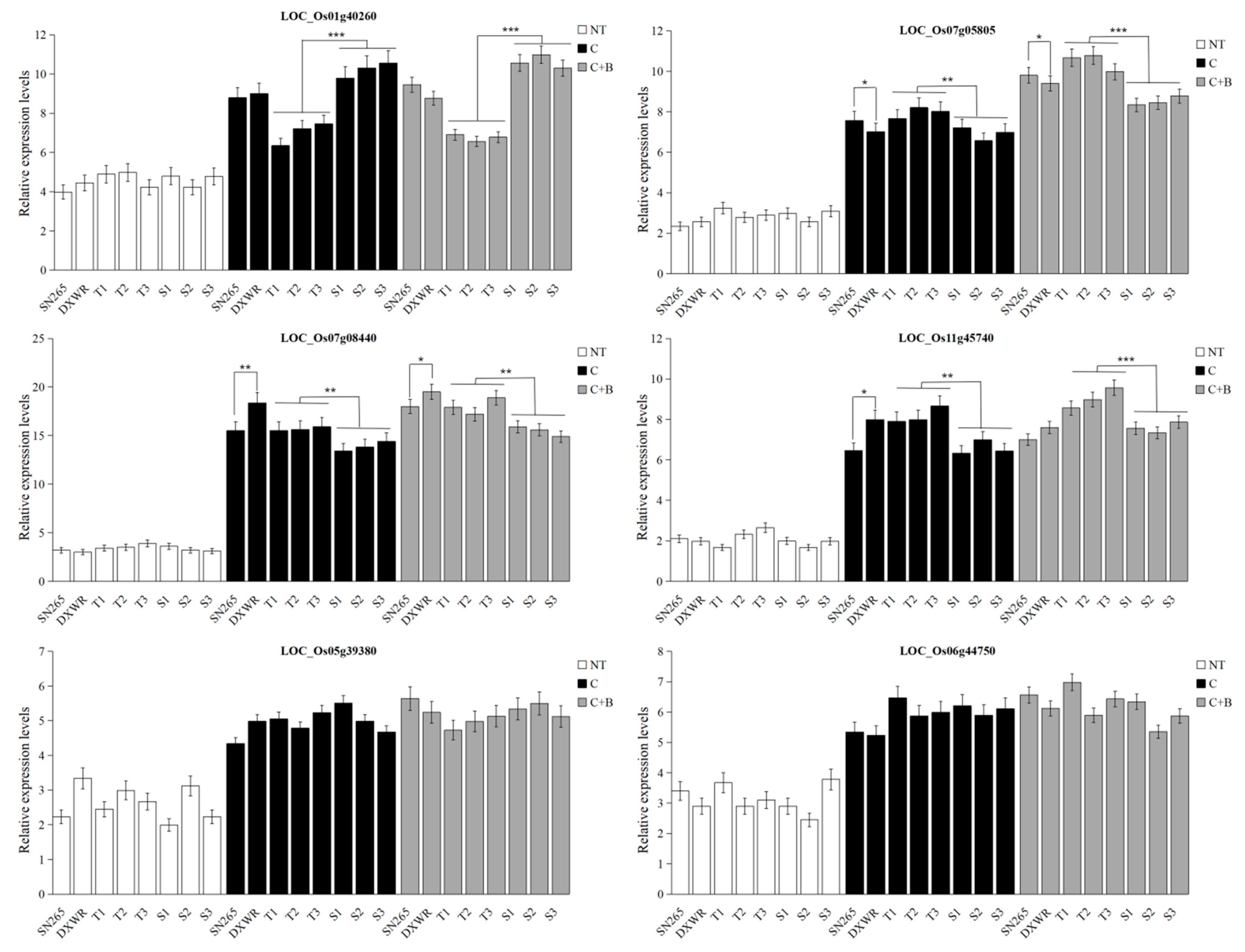
2.4. Prediction of Candidate Genes
2.5. Validation of Candidate Genes
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Population Development
4.2. Phenotypic Evaluation for Cold Tolerance
4.3. Genotyping-by-Sequencing and SNP Identification
4.4. Construction of Linkage Map and QTL Analysis
4.5. Prediction and Validation of Candidate Genes
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cheng, S.H.; Cao, L.Y.; Zhuang, J.Y.; Chen, S.G.; Zhan, X.D.; Fan, Y.Y.; Min, S.K. Super hybrid rice breeding in China: Achievements and prospects. J. Integr. Plant Biol. 2007, 49, 805–810. [Google Scholar] [CrossRef]
- Fujino, K.; Sekiguchi, H.; Sato, T.; Kiuchi, H.; Nonoue, Y.; Takeuchi, Y.; Ando, T.; Lin, S.Y.; Yano, M. Mapping of Quantitative Trait Loci Controlling Low-Temperature Germinability in Rice (Oryza sativa L.). Theor. Appl. Genet. 2004, 108, 794–799. [Google Scholar] [CrossRef]
- Nakagahra, M.; Okuno, K.; Vaughan, D. Rice Genetic Resources: History, Conservation, Investigative Characterization and Use in Japan. Plant Mol. Biol. 1997, 35, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Najeeb, S.; Ali, J.; Mahender, A.; Pang, Y.L.; Zilhas, J.; Murugaiyan, V.; Vemireddy, L.R.; Li, Z. Identification of Main-Effect Quantitative Trait Loci (QTLs) for Low-Temperature Stress Tolerance Germination-and Early Seedling Vigor-Related Traits in Rice (Oryza sativa L.). Mol. Breed. 2020, 40, 10. [Google Scholar] [CrossRef] [PubMed]
- Miura, K.; Lin, S.Y.; Yano, M.; Nagamine, T. Mapping Quantitative Trait Loci Controlling Low Temperature Germinability in Rice (Oryza sativa L.). Breed. Sci. 2001, 51, 293–299. [Google Scholar] [CrossRef]
- Yang, L.; Wang, J.; Han, Z.; Lei, L.; Liu, H.L.; Zheng, H.; Xin, W.; Zou, D. Combining QTL-seq and Linkage Mapping to Fine Map a Candidate Gene in qCTS6 for Cold Tolerance at the Seedling Stage in Rice. BMC Plant Biol. 2021, 21, 278. [Google Scholar] [CrossRef]
- Mielczarek, M.; Szyda, J. Review of Alignment and Snp Calling Algorithms for Next-Generation Sequencing Data. J. Appl. Genet. 2016, 57, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Wang, H.; Michal, J.J.; Zhou, X.; Liu, B.; Woods, L.C.; Fuchs, R.A. Genome Wide Sampling Sequencing for Snp Genotyping: Methods, Challenges and Future Development. Int. J. Biol. Sci. 2016, 12, 100–108. [Google Scholar] [CrossRef]
- Spindel, J.; Wright, M.; Chen, C.; Cobb, J.; Gage, J.; Harrington, S.; Lorieux, M.; Ahmadi, N.; McCouch, S. Bridging the Genotyping Gap: Using Genotyping by Sequencing (GBS) to Add High-Density Snp Markers and New Value to Traditional Bi-Parental Mapping and Breeding Populations. Theor. Appl. Genet. 2013, 126, 2699–2716. [Google Scholar] [CrossRef]
- Arbelaez, J.D.; Moreno, L.T.; Singh, N.; Tung, C.W.; Maron, L.G.; Ospina, Y.; Martinez, C.P.; Grenier, C.; Lorieux, M.; McCouch, S. Development and GBS-Genotyping of Introgression Lines (ILS) Using Two Wild Species of Rice, O. meridionalis and O. rufipogon, in a Common Recurrent Parent, O. sativa cv. Curinga. Mol. Breed. 2015, 35, 81. [Google Scholar] [CrossRef] [Green Version]
- Atwell, B.J.; Wang, H.; Scafaro, A.P. Could Abiotic Stress Tolerance in Wild Relatives of Rice Be Used to Improve Oryza Sativa? Plant Sci. 2014, 215–216, 48–58. [Google Scholar] [CrossRef]
- Zhang, K.; Su, J.; Xu, M.; Zhou, Z.; Zhu, X.; Ma, X.; Hou, J.; Tan, L.; Zhu, Z.; Cai, H.; et al. A Common Wild Rice-Derived Boc1 Allele Reduces Callus Browning in Indica Rice Transformation. Nat. Commun. 2020, 11, 443. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Chen, Q.; Wang, S.; Hong, Y.; Wang, Z. Rice and Cold Stress: Methods for Its Evaluation and Summary of Cold Tolerance-Related Quantitative Trait Loci. Rice 2014, 7, 24. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Sun, C.; Tan, L.; Fu, Y.; Li, D.; Wang, X. Identification and Mapping of Quantitative Trait Loci Controlling Cold-Tolerance of Chinese Common Wild Rice (O. rufipogon Griff.) at Booting to Flowering Stages. Chin. Sci. Bull. 2003, 48, 2068–2071. [Google Scholar] [CrossRef]
- Tian, F.; Li, D.J.; Fu, Q.; Zhu, Z.F.; Fu, Y.C.; Wang, X.K.; Sun, C.Q. Construction of Introgression Lines Carrying Wild Rice (Oryza Rufipogon Griff.) Segments in Cultivated Rice (Oryza sativa L.) Background and Characterization of Introgressed Segments Associated with Yield-Related Traits. Theor. Appl. Genet. 2006, 112, 570–580. [Google Scholar] [CrossRef]
- Xiao, N.; Huang, W.N.; Li, A.H.; Gao, Y.; Li, Y.H.; Pan, C.H.; Ji, H.; Zhang, X.X.; Dai, Y.; Dai, Z.Y.; et al. Fine Mapping of the qLOP2 and qPSR2-1 Loci Associated with Chilling Stress Tolerance of Wild Rice Seedlings. Theor. Appl. Genet. 2015, 128, 173–185. [Google Scholar] [CrossRef]
- Liang, Y.; Yan, C.; Zheng, J.; Nan, W.; Qin, X.; Zhang, H. Locating Qtl Associated with Spike Traits of Dongxiang Wild Rice (Oryza rufipogon Griff.). Euphytica 2019, 215, 26. [Google Scholar] [CrossRef]
- Liang, Y.; Zheng, J.; Yan, C.; Li, X.; Liu, S.; Zhou, J.; Qin, X.; Nan, W.; Yang, Y.; Zhang, H. Locating Qtls Controlling Overwintering Trait in Chinese Perennial Dongxiang Wild Rice. Mol. Genet. Genom. 2018, 293, 81–93. [Google Scholar] [CrossRef]
- Bai, L.W.; Liu, J.; Dai, L.F.; Deng, Q.W.; Chen, Y.L.; Xie, J.K.; Luo, X.D. Identification and Characterisation of Cold Stress-Related Proteins in Oryza rufipogon at the Seedling Stage Using Label-Free Quantitative Proteomic Analysis. Funct. Plant Biol. 2021, 48, 542–555. [Google Scholar] [CrossRef]
- Sharma, I.; Kaur, N.; Pati, P.K. Brassinosteroids: A Promising Option in Deciphering Remedial Strategies for Abiotic Stress Tolerance in Rice. Front. Plant Sci. 2017, 8, 2151. [Google Scholar] [CrossRef]
- Manghwar, H.; Hussain, A.; Ali, Q.; Liu, F. Brassinosteroids (BRs) Role in Plant Development and Coping with Different Stresses. Int. J. Mol. Sci. 2022, 23, 1012. [Google Scholar] [CrossRef]
- Basit, F.; Liu, J.; An, J.; Chen, M.; He, C.; Zhu, X.; Li, Z.; Hu, J.; Guan, Y. Brassinosteroids as a Multidimensional Regulator of Plant Physiological and Molecular Responses under Various Environmental Stresses. Environ. Sci. Pollut. Res. Int. 2021, 28, 44768–44779. [Google Scholar] [CrossRef] [PubMed]
- Sadura, I.; Janeczko, A. Brassinosteroids and the Tolerance of Cereals to Low and High Temperature Stress: Photosynthesis and the Physicochemical Properties of Cell Membranes. Int. J. Mol. Sci. 2021, 23, 342. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Ye, K.; Shi, Y.; Cheng, J.; Zhang, X.; Yang, S. BZR1 Positively Regulates Freezing Tolerance Via CBF-Dependent and CBF-Independent Pathways in Arabidopsis. Mol. Plant 2017, 10, 545–559. [Google Scholar] [CrossRef] [PubMed]
- Peres, A.; Soares, J.S.; Tavares, R.G.; Righetto, G.; Zullo, M.A.T.; Mandava, N.B.; Menossi, M. Brassinosteroids, the Sixth Class of Phytohormones: A Molecular View from the Discovery to Hormonal Interactions in Plant Development and Stress Adaptation. Int. J. Mol. Sci. 2019, 20, 331. [Google Scholar] [CrossRef]
- Ye, K.; Li, H.; Ding, Y.; Shi, Y.; Song, C.; Gong, Z.; Yang, S. Brassinosteroid-Insensitive2 Negatively Regulates the Stability of Transcription Factor Ice1 in Response to Cold Stress in Arabidopsis. Plant Cell 2019, 31, 2682–2696. [Google Scholar] [CrossRef]
- Hwang, O.J.; Back, K. Melatonin Deficiency Confers Tolerance to Multiple Abiotic Stresses in Rice Via Decreased Brassinosteroid Levels. Int. J. Mol. Sci. 2019, 20, 5173. [Google Scholar] [CrossRef]
- Wu, W.; Zhou, W.; Tang, C.; Liu, K.; Zeng, H.; Wang, Y. Effects of Exogenous 2,4-Epibrassinolide on Germination and Physiological Characteristics of Rice Seeds under Chilling Stress. Mol. Plant Breed. 2020, 18, 4427–4434. [Google Scholar] [CrossRef]
- Guo, H.; Wu, T.; Li, S.; He, Q.; Yang, Z.; Zhang, W.; Gan, Y.; Sun, P.; Xiang, G.; Zhang, H.; et al. The Methylation Patterns and Transcriptional Responses to Chilling Stress at the Seedling Stage in Rice. Int. J. Mol. Sci. 2019, 20, 5089. [Google Scholar] [CrossRef]
- Guo, Z.F.; Wang, H.T.; Cheng, Y.S.; Guo, S.Q.; Zhang, W.Z.; Xu, Z.J.; Li, M.M.; Huang, J.; Zhao, M.H. QTL Mapping Analysis for Cold Tolerance under Cold Stress and Brassinosteroids combined Cold Treatment at Germination and Bud Burst Stages in Rice. Front. Plant Sci. 2022, 13, 938339. [Google Scholar] [CrossRef]
- Hamzelou, S.; Kamath, K.S.; Masoomi-Aladizgeh, F.; Johnsen, M.M.; Atwell, B.J.; Haynes, P.A. Wild and Cultivated Species of Rice Have Distinctive Proteomic Responses to Drought. Int. J. Mol. Sci. 2020, 21, 5980. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Li, W.; Zhang, Q.J.; Zhang, Y.; Tong, Y.; Li, K.; Liu, Y.L.; Gao, L.Z. The Draft Genome Sequence of an Upland Wild Rice Species, Oryza granulata. Sci. Data 2020, 7, 131. [Google Scholar] [CrossRef] [PubMed]
- Qiao, W.; Wang, Y.; Xu, R.; Yang, Z.; Sun, Y.; Su, L.; Zhang, L.; Wang, J.; Huang, J.; Zheng, X.; et al. A Functional Chromogen Gene C from Wild Rice Is Involved in a Different Anthocyanin Biosynthesis Pathway in Indica and Japonica. Theor. Appl. Genet. 2021, 134, 1531–1543. [Google Scholar] [CrossRef] [PubMed]
- Xing, J.; Zhang, D.; Yin, F.; Zhong, Q.; Wang, B.; Xiao, S.; Ke, X.; Wang, L.; Zhang, Y.; Zhao, C.; et al. Identification and Fine-Mapping of a New Bacterial Blight Resistance Gene, Xa47(T), in G252, an Introgression Line of Yuanjiang Common Wild Rice (Oryza rufipogon). Plant Dis. 2021, 105, 4106–4112. [Google Scholar] [CrossRef]
- Liu, W.; Ghouri, F.; Yu, H.; Li, X.; Yu, S.; Shahid, M.Q.; Liu, X. Genome Wide Re-Sequencing of Newly Developed Rice Lines from Common Wild Rice (Oryza rufipogon Griff.) for the Identification of Nbs-Lrr Genes. PLoS ONE 2017, 12, e0180662. [Google Scholar] [CrossRef]
- Yuan, R.; Zhao, N.; Usman, B.; Luo, L.; Liao, S.; Qin, Y.; Nawaz, G.; Li, R. Development of Chromosome Segment Substitution Lines (Cssls) Derived from Guangxi Wild Rice (Oryza rufipogon Griff.) under Rice (Oryza sativa L.) Background and the Identification of Qtls for Plant Architecture, Agronomic Traits and Cold Tolerance. Genes 2020, 11, 980. [Google Scholar] [CrossRef]
- Yun, Y.T.; Chung, C.T.; Lee, Y.J.; Na, H.J.; Lee, J.C.; Lee, S.G.; Lee, K.W.; Yoon, Y.H.; Kang, J.W.; Lee, H.S.; et al. Qtl Mapping of Grain Quality Traits Using Introgression Lines Carrying Oryza rufipogon Chromosome Segments in Japonica Rice. Rice 2016, 9, 62. [Google Scholar] [CrossRef]
- Cen, W.; Liu, J.; Lu, S.; Jia, P.; Yu, K.; Han, Y.; Li, R.; Luo, J. Comparative Proteomic Analysis of Qtl Cts-12 Derived from Wild Rice (Oryza rufipogon Griff.), in the Regulation of Cold Acclimation and De-Acclimation of Rice (Oryza sativa L.) in Response to Severe Chilling Stress. BMC Plant Biol. 2018, 18, 163. [Google Scholar] [CrossRef]
- Wang, D.Z.; Jin, Y.N.; Ding, X.H.; Wang, W.J.; Zhai, S.S.; Bai, L.P.; Guo, Z.F. Gene Regulation and Signal Transduction in the ICE-CBF-COR Signaling Pathway During Cold Stress in Plants. Biochemistry 2017, 82, 1103–1117. [Google Scholar] [CrossRef]
- Huang, G.T.; Ma, S.L.; Bai, L.P.; Zhang, L.; Ma, H.; Jia, P.; Liu, J.; Zhong, M.; Guo, Z.F. Signal Transduction During Cold, Salt, and Drought Stresses in Plants. Mol. Biol. Rep. 2012, 39, 969–987. [Google Scholar] [CrossRef]
- Hwarari, D.; Guan, Y.; Ahmad, B.; Movahedi, A.; Min, T.; Hao, Z.; Lu, Y.; Chen, J.; Yang, L. ICE-CBF-COR Signaling Cascade and Its Regulation in Plants Responding to Cold Stress. Int. J. Mol. Sci. 2022, 23, 1549. [Google Scholar] [CrossRef]
- Dubouzet, J.G.; Sakuma, Y.; Ito, Y.; Kasuga, M.; Dubouzet, E.G.; Miura, S.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Osdreb Genes in Rice, Oryza sativa L., Encode Transcription Activators That Function in Drought-, High-Salt- and Cold-Responsive Gene Expression. Plant J. 2003, 33, 751–763. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Sun, S.; Xu, D.; Lan, H.; Sun, H.; Wang, Z.; Bao, Y.; Wang, J.; Tang, H.; Zhang, H. A Tfiiia-Type Zinc Finger Protein Confers Multiple Abiotic Stress Tolerances in Transgenic Rice (Oryza sativa L.). Plant Mol. Biol. 2012, 80, 337–350. [Google Scholar] [CrossRef]
- Li, F.; Guo, S.; Zhao, Y.; Chen, D.; Chong, K.; Xu, Y. Overexpression of a Homopeptide Repeat-Containing Bhlh Protein Gene (Orbhlh001) from Dongxiang Wild Rice Confers Freezing and Salt Tolerance in Transgenic Arabidopsis. Plant Cell Rep. 2010, 29, 977–986. [Google Scholar] [CrossRef] [PubMed]
- Eremina, M.; Unterholzner, S.J.; Rathnayake, A.I.; Castellanos, M.; Khan, M.; Kugler, K.G.; May, S.T.; Mayer, K.F.; Rozhon, W.; Poppenberger, B. Brassinosteroids Participate in the Control of Basal and Acquired Freezing Tolerance of Plants. Proc. Natl. Acad. Sci. USA 2016, 113, e5982–e5991. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, J.; Li, F.; Liu, H.; Yang, W.; Chong, K.; Xu, Y. OsMAPK3 Phosphorylates OsbHLH002/OsICE1 and Inhibits Its Ubiquitination to Activate OsTPP1 and Enhances Rice Chilling Tolerance. Dev. Cell 2017, 43, 731–743. [Google Scholar] [CrossRef]
- Jiang, B.; Shi, Y.; Zhang, X.; Xin, X.; Qi, L.; Guo, H.; Li, J.; Yang, S. PIF3 Is a Negative Regulator of the Cbf Pathway and Freezing Tolerance in Arabidopsis. Proc. Natl. Acad. Sci. USA 2017, 114, e6695–e6702. [Google Scholar] [CrossRef]
- Shi, Y.; Tian, S.; Hou, L.; Huang, X.; Zhang, X.; Guo, H.; Yang, S. Ethylene Signaling Negatively Regulates Freezing Tolerance by Repressing Expression of CBF and Type-A ARR Genes in Arabidopsis. Plant Cell 2012, 24, 2578–2595. [Google Scholar] [CrossRef]
- Singh, I.; Kumar, U.; Singh, S.K.; Gupta, C.; Singh, M.; Kushwaha, S.R. Physiological and Biochemical Effect of 24-Epibrassinoslide on Cold Tolerance in Maize Seedlings. Physiol. Mol. Biol. Plants 2012, 18, 229–236. [Google Scholar] [CrossRef]
- Janeczko, A.; Pociecha, E.; Dziurka, M.; Jurczyk, B.; Libik-Konieczny, M.; Oklestkova, J.; Novák, O.; Pilarska, M.; Filek, M.; Rudolphi-Skórska, E.; et al. Changes in Content of Steroid Regulators During Cold Hardening of Winter Wheat-Steroid Physiological/Biochemical Activity and Impact on Frost Tolerance. Plant Physiol. Biochem. 2019, 139, 215–228. [Google Scholar] [CrossRef]
- Pociecha, E.; Dziurka, M.; Oklestkova, J.; Janeczko, A. Brassinosteroids Increase Winter Survival of Winter Rye (Secale Cereale L.) by Affecting Photosynthetic Capacity and Carbohydrate Metabolism During the Cold Acclimation Process. Plant Growth Regul. 2016, 80, 127–135. [Google Scholar] [CrossRef]
- Viana, V.E.; Carlos da Maia, L.; Busanello, C.; Pegoraro, C.; Costa de Oliveira, A. When Rice Gets the Chills: Comparative Transcriptome Profiling at Germination Shows Wrky Transcription Factor Responses. Plant Biol. 2021, 23, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, Y.; de la Bastide, M.; Hamilton, J.P.; Kanamori, H.; McCombie, W.R.; Ouyang, S.; Schwartz, D.C.; Tanaka, T.; Wu, J.; Zhou, S.; et al. Improvement of the Oryza Sativa Nipponbare Reference Genome Using Next Generation Sequence and Optical Map Data. Rice 2013, 6, 4. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.; Li, H.H.; Zhang, L.Y.; Wang, J.K. QTL IciMapping: Integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations. Crop J. 2015, 3, 269–283. [Google Scholar] [CrossRef]
- McCouch, S.R. Gene Nomenclature System for Rice. Rice 2008, 1, 72–84. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative Pcr and the 2-DDCt Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chr | Number of Bin Markers | Genetic Distance (cM) | Ave. Genetic Distance between Markers (cM) | Ave. Interval (cM) | Interval Range (cM) |
|---|---|---|---|---|---|
| 1 | 104 | 268.20 | 0.4 | 25.8 | 5.1–120.5 |
| 2 | 112 | 288.60 | 0.3 | 25.8 | 4.0–74.5 |
| 3 | 116 | 314.04 | 0.3 | 27.1 | 4.9–73.6 |
| 4 | 117 | 346.46 | 0.3 | 29.6 | 7.5–88.7 |
| 5 | 81 | 229.41 | 0.4 | 28.3 | 3.6–127.8 |
| 6 | 100 | 293.94 | 0.4 | 29.4 | 1.9–98.6 |
| 7 | 77 | 186.74 | 0.4 | 24.3 | 4.0–75.6 |
| 8 | 91 | 271.54 | 0.5 | 29.8 | 6.3–146.3 |
| 9 | 79 | 227.08 | 0.3 | 28.7 | 6.2–121.3 |
| 10 | 85 | 230.99 | 0.3 | 27.2 | 5.4–112.6 |
| 11 | 105 | 303.46 | 0.3 | 28.9 | 2.0–126.8 |
| 12 | 78 | 227.87 | 0.3 | 29.2 | 2.0–137.7 |
| All | 1145 | 3188.33 | 0.3 (Ave.) | 27.8 (Ave.) | 1.9–146.3 |
| Condition | QTL | Chr | Interval | LOD | PVE (%) | Add |
|---|---|---|---|---|---|---|
| Cold | qSSL1-2 | 1 | 22,514,210–23,217,591 | 4.7469 | 9.8918 | 0.0402 |
| qSSL3-1 | 3 | 11,339,312–12,047,081 | 3.3850 | 7.7212 | −0.0398 | |
| qSSL11-1 | 11 | 6,640,029–6,662,625 | 4.8162 | 8.6890 | −0.0356 | |
| BR + C-N | qSSL1-1 | 1 | 12,150,755–12,275,956 | 4.3992 | 6.3653 | −0.0364 |
| qSSL2-1 | 2 | 8,052,187–8,229,991 | 3.6184 | 10.7271 | 0.0374 | |
| qSSL4-1 | 4 | 35,027,717–35,296,119 | 5.0857 | 9.4770 | 0.0401 | |
| qSSL11-2 | 11 | 16,545,655–16,884,915 | 3.9168 | 9.1290 | −0.0356 | |
| BR + C-C | qSSL2-2 | 2 | 34,425,783–34,742,671 | 3.9258 | 15.0490 | −0.0537 |
| qSSL7-1 | 7 | 23,483,968–23,485,531 | 3.1103 | 4.1682 | 0.0259 |
| Condition | QTL | Chr | Interval | LOD | PVE (%) | Add |
|---|---|---|---|---|---|---|
| Cold | qSRL10-2 | 10 | 20,367,970–20,569,786 | 3.0102 | 7.7200 | 0.1398 |
| qSRL11-2 | 11 | 16,889,698–17,163,458 | 3.1469 | 7.6157 | −0.1713 | |
| BR + C-N | qSRL1-3 | 1 | 28,250,887–28,381,910 | 3.0384 | 9.1064 | 0.1096 |
| qSRL8-1 | 8 | 23,572,813–23,607,999 | 3.6878 | 9.4108 | 0.1096 | |
| BR + C-C | qSRL1-1 | 1 | 9,795,818–9,824,282 | 4.0158 | 1.6769 | −0.2551 |
| qSRL1-2 | 1 | 27,714,994–27,766,901 | 3.1117 | 1.1912 | −0.1597 | |
| qSRL1-4 | 1 | 36,461,005–37,047,678 | 4.0028 | 0.7550 | 0.1129 | |
| qSRL1-5 | 1 | 40,759,127–40,984,457 | 3.0319 | 0.9515 | 0.9515 | |
| qSRL2-1 | 2 | 18,887,339–19,398,580 | 3.3044 | 1.8057 | −0.2096 | |
| qSRL3-1 | 3 | 7,023,285–7,063,090 | 3.0500 | 1.6192 | −0.2303 | |
| qSRL4-1 | 4 | 35,487,338–35,496,266 | 3.2674 | 1.7908 | −0.2234 | |
| qSRL5-1 | 5 | 1,622,816–2,206,645 | 3.1717 | 1.7711 | −0.2284 | |
| qSRL5-2 | 5 | 15,157,600–16,159,826 | 3.2548 | 1.7136 | −0.2395 | |
| qSRL5-3 | 5 | 23,101,672–23,110,060 | 4.1914 | 1.7021 | −0.2340 | |
| qSRL5-4 | 5 | 29,195,759–29,365,239 | 3.0047 | 1.7353 | −0.2379 | |
| qSRL6-1 | 6 | 34,503–49,710 | 3.2385 | 1.4471 | −0.2590 | |
| qSRL6-2 | 6 | 49,710–75,033 | 3.7394 | 1.5946 | −0.2537 | |
| qSRL6-3 | 6 | 2,026,942–2,573,648 | 3.3184 | 1.6972 | −0.2214 | |
| qSRL6-4 | 6 | 26,319,487–27,021,092 | 4.1196 | 1.6335 | −0.2495 | |
| qSRL6-5 | 6 | 27,021,092–27,066,725 | 3.6184 | 1.6633 | −0.2488 | |
| qSRL7-1 | 7 | 2,206,752–2,342,190 | 3.1001 | 1.2549 | 0.1573 | |
| qSRL7-2 | 7 | 29,374,392–29,637,653 | 3.3382 | 1.5548 | −0.2212 | |
| qSRL9-1 | 9 | 2,137,063–2,395,182 | 3.3416 | 1.6710 | −0.2369 | |
| qSRL9-2 | 9 | 2,395,182–2,547,966 | 3.8906 | 1.7777 | −0.2301 | |
| qSRL9–3 | 9 | 11,240,451–11,542,598 | 3.5838 | 1.6649 | −0.2410 | |
| qSRL10-1 | 10 | 5,080,068–5,747,438 | 3.0349 | 1.6997 | −0.2413 | |
| qSRL11-1 | 11 | 8,584,535–9,024,140 | 3.4737 | 1.3106 | 0.2003 | |
| qSRL12-1 | 12 | 978,610–3,267,494 | 3.2328 | 1.6911 | −0.2408 | |
| qSRL12-2 | 12 | 18,216,503–18,482,234 | 3.1496 | 1.8460 | −0.1963 | |
| qSRL12-3 | 12 | 18,482,234–19,002,436 | 4.3086 | 1.6868 | −0.2264 | |
| qSRL12-4 | 12 | 20,179,243–20,499,113 | 3.1560 | 1.6292 | −0.2544 | |
| qSRL12-5 | 12 | 20,499,113–20,522,990 | 3.0551 | 1.6346 | −0.2470 | |
| qSRL12-6 | 12 | 25,801,108–25,921,238 | 4.0588 | 1.5263 | −0.2428 |
| Condition | QTL | Chr | Interval | LOD | PVE (%) | Add |
|---|---|---|---|---|---|---|
| Cold | qSDW1-1 | 1 | 8,430,459–9,795,818 | 3.0094 | 1.6880 | −0.4911 |
| qSDW1-3 | 1 | 9,795,818–9,824,282 | 3.6108 | 1.7076 | −0.4813 | |
| qSDW2-3 | 2 | 23,029,608–23,036,193 | 3.2848 | 1.8389 | −0.3107 | |
| qSDW4-1 | 4 | 2,035,471–2,255,931 | 3.6631 | 0.8453 | 0.1214 | |
| qSDW5-1 | 5 | 28,253,886–28,304,136 | 3.2173 | 1.8218 | −0.3146 | |
| qSDW6-1 | 6 | 34,503–49,710 | 5.2751 | 1.8124 | −0.3869 | |
| qSDW6-5 | 6 | 49,710–75,033 | 4.6802 | 1.7782 | −0.3912 | |
| qSDW6-8 | 6 | 75,033–86,257 | 3.9971 | 1.7254 | −0.3676 | |
| qSDW6-11 | 6 | 543,717–961,403 | 4.5950 | 1.7380 | −0.3096 | |
| qSDW6-13 | 6 | 918,728–8,551,070 | 3.1224 | 1.7444 | −0.3801 | |
| qSDW6-15 | 6 | 26,319,487–27,021,092 | 3.5723 | 1.7885 | −0.3040 | |
| qSDW6-18 | 6 | 27,021,092–27,066,725 | 3.0443 | 1.7837 | −0.3076 | |
| qSDW8-4 | 8 | 22,126,649–23,227,340 | 3.4343 | 1.9619 | −0.2550 | |
| qSDW8-6 | 8 | 27,136,983–27,371,918 | 3.5504 | 1.8118 | −0.3283 | |
| qSDW9-2 | 9 | 5,908,433–5,989,512 | 3.2579 | 1.5819 | −0.2097 | |
| qSDW9-4 | 9 | 11,240,451–11,542,598 | 3.1615 | 1.8326 | −0.2532 | |
| qSDW12-1 | 12 | 978,610–3,267,494 | 3.7306 | 1.8958 | −0.3331 | |
| qSDW12-4 | 12 | 20,179,243–20,499,113 | 3.0638 | 1.8157 | −0.3170 | |
| qSDW12-5 | 12 | 20,499,113–20,522,990 | 3.1036 | 1.8231 | −0.3093 | |
| BR + C-N | qSDW1-2 | 1 | 8,430,459–9,795,818 | 4.1163 | 1.0370 | −0.4150 |
| qSDW1-4 | 1 | 9,795,818–9,824,282 | 9.6158 | 1.6340 | −1.3208 | |
| qSDW2-2 | 2 | 14,192,829–15,359,168 | 9.4888 | 1.6340 | 1.3208 | |
| qSDW6-2 | 6 | 34,503–49,710 | 10.4920 | 1.6340 | −1.3208 | |
| qSDW6-6 | 6 | 49,710–75,033 | 11.4737 | 1.6340 | −1.3208 | |
| qSDW6-7 | 6 | 75,033–86,257 | 9.8876 | 1.6340 | −1.3208 | |
| qSDW6-9 | 6 | 536,139–543,717 | 9.1114 | 1.6340 | −1.3208 | |
| qSDW6-12 | 6 | 543,717–961,403 | 9.1993 | 1.6340 | −1.3208 | |
| qSDW6-14 | 6 | 8,551,070–9,187,287 | 10.4330 | 1.6340 | −1.3208 | |
| qSDW6-16 | 6 | 26,319,487–27,021,092 | 3.3065 | 0.9447 | −0.3687 | |
| qSDW6-17 | 6 | 27,021,092–27,066,725 | 8.0183 | 1.6340 | −1.3208 | |
| qSDW8-1 | 8 | 20,095,951–20,541,282 | 4.6413 | 1.6340 | 1.3208 | |
| qSDW8-5 | 8 | 27,136,983–27,371,918 | 8.6442 | 1.6340 | −1.3208 | |
| qSDW9-1 | 9 | 2,395,182–2,547,966 | 7.9756 | 1.6340 | −1.3208 | |
| qSDW9-3 | 9 | 5,908,433–5,989,512 | 3.0530 | 0.4007 | −0.1190 | |
| qSDW9-5 | 9 | 16,774,761–16,829,125 | 8.6080 | 1.6340 | −1.3208 | |
| qSDW10-1 | 10 | 2,188,025–2,329,734 | 8.8075 | 1.6340 | −1.3208 | |
| qSDW11-2 | 11 | 26,184,650–26,729,025 | 8.8993 | 1.6340 | −1.3208 | |
| qSDW12-2 | 12 | 978,610–3,267,494 | 8.3168 | 1.6340 | −1.3208 | |
| qSDW12-3 | 12 | 20,179,243–20,499,113 | 8.3537 | 1.6340 | −1.3208 | |
| qSDW12-6 | 12 | 20,499,113–20,522,990 | 8.0866 | 1.6340 | −1.3208 | |
| BR + C-C | qSDW2-1 | 2 | 14,192,829–15,359,168 | 9.2210 | 6.5896 | 0.9641 |
| qSDW3-1 | 3 | 28,456,345–29,627,234 | 3.9577 | 1.7956 | −0.1035 | |
| qSDW6-3 | 6 | 34,503–49,710 | 6.6853 | 4.5867 | −0.8043 | |
| qSDW6-4 | 6 | 49,710–75,033 | 6.5255 | 4.5867 | −0.8043 | |
| qSDW6-10 | 6 | 536,139–543,717 | 5.2886 | 4.5867 | −0.8043 | |
| qSDW8-2 | 8 | 20,095,951–20,541,282 | 3.9832 | 6.0808 | 0.9261 | |
| qSDW8-3 | 8 | 20,095,951–20,541,282 | 3.6278 | 1.6461 | 0.1350 | |
| qSDW11-1 | 11 | 23,793,431–23,795,247 | 4.9421 | 4.5867 | −0.8043 | |
| qSDW12-7 | 12 | 25,921,238–26,742,306 | 4.0396 | 1.8563 | 0.1470 |
| Condition | QTL | Chr | Interval | LOD | PVE (%) | Add |
|---|---|---|---|---|---|---|
| Cold | qSWW1-3 | 1 | 17,059,460–18,491,456 | 8.4626 | 18.2439 | 0.1201 |
| qSWW7-1 | 7 | 187,721–321,199 | 4.1511 | 8.7113 | −0.1427 | |
| qSWW12-2 | 12 | 19,002,436–19,612,427 | 3.9255 | 7.5760 | 0.0745 | |
| BR + C-N | qSWW3-1 | 3 | 17,877,339–18,608,236 | 3.7539 | 2.3749 | 0.1061 |
| qSWW6-2 | 6 | 49,710–75,033 | 5.2304 | 5.4240 | −0.7790 | |
| qSWW6-3 | 6 | 75,033–86,257 | 3.6894 | 5.4240 | −0.7790 | |
| qSWW6-6 | 6 | 8,551,070–9,187,287 | 4.6827 | 5.4241 | −0.7790 | |
| qSWW11-2 | 11 | 27,344,371–27,803,834 | 4.9243 | 3.2351 | 0.1113 | |
| BR + C-C | qSWW1-1 | 1 | 8,430,459–9,795,818 | 3.9405 | 0.7179 | −0.6502 |
| qSWW1-2 | 1 | 9,795,818–9,824,282 | 4.2951 | 0.7179 | −0.6501 | |
| qSWW4-1 | 4 | 30,444,673–30,569,820 | 3.1941 | 0.7601 | −0.4591 | |
| qSWW5-1 | 5 | 28,253,886–28,304,136 | 3.1392 | 0.8057 | −0.3553 | |
| qSWW6-1 | 6 | 49,710–75,033 | 5.6121 | 0.7489 | −0.4876 | |
| qSWW6-4 | 6 | 75,033–86,257 | 4.0525 | 0.7134 | −0.2935 | |
| qSWW6-5 | 6 | 536,139–543,717 | 3.3069 | 0.7941 | −0.3672 | |
| qSWW6-7 | 6 | 8,551,070–9,187,287 | 5.4922 | 0.7179 | −0.6503 | |
| qSWW6-8 | 6 | 9,187,287–9,300,770 | 3.5735 | 0.7532 | −0.4835 | |
| qSWW9-1 | 9 | 16,774,761–16,829,125 | 4.1466 | 0.8111 | −0.3491 | |
| qSWW10-1 | 10 | 2,188,025–2,329,734 | 3.4112 | 0.7576 | −0.4731 | |
| qSWW11-1 | 11 | 23,793,431–23,795,247 | 3.4724 | 0.7573 | −0.4714 | |
| qSWW12-1 | 12 | 18,482,234–19,002,436 | 4.0664 | 0.7574 | −0.4714 | |
| qSWW12-3 | 12 | 20,179,243–20,499,113 | 3.5313 | 0.7576 | −0.4694 | |
| qSWW12-4 | 12 | 20,499,113–20,522,990 | 3.3591 | 0.7582 | −0.4672 |
| Condition | QTL | Chr | Locus Name | Gene Coordinates | Gene Product |
|---|---|---|---|---|---|
| C | qSSL1-2 | 1 | LOC_Os01g40260 | 22,731,943–22,733,237 | OsWRKY77- Superfamily of TFs with WRKY and zinc finger domains |
| C + BR | qSWW11-2 | 11 | LOC_Os11g45740 | 27,670,321–27,673,334 | MYB family transcription factor |
| C + BR-N | qSSL7-1 | 7 | LOC_Os07g05805 | 23,483,968–23,485,531 | OsBZR1, transcription factor, Brassinosteroid (BR)-regulated growth response |
| C | qSSL7-3 | 7 | LOC_Os07g08440 | 4,338,514–4,342,219 | bHLH transcription factor |
| C/C + BR-N | qSDW6-18/17 | 6 | LOC_Os06g44750 | 27,025,437–27,029,339 | AP2 domain-containing protein, expressed |
| C + BR-N/C + BR-C | qSDW6-9/10 | 6 | LOC_Os06g01966 | 543,057–545,780 | auxin-induced protein 5NG4, putative, expressed |
| C + BR-C | qSRL12-6 | 12 | LOC_Os12g41820 | 25,901,456–25,907,573 | heat shock protein DnaJ, putative, expressed |
| C | qSSL7-1 | 7 | LOC_Os07g01480 | 306,009–307,555 | oxygen evolving enhancer protein 3 domain-containing protein, expressed |
| C + BR-C | qSRL12-6 | 12 | LOC_Os12g41700 | 25,815,277–25,818,874 | LSD1, zinc finger domain-containing protein, expressed |
| C + BR-C | qSRL5-3 | 5 | LOC_Os05g39380 | 23,101,671–23,104,153 | zinc finger, C3HC4 type domain-containing protein, expressed |
| C + BR-C | qSWW9-1 | 9 | LOC_Os09g27650 | 16,822,234–16,825,686 | ZOS9-14, C2H2 zinc finger protein, expressed |
| C + BR-N | qSDW9-5 | 9 | LOC_Os09g27660 | 16,829,409–16,837,307 | OsFBO21, F-box, and other domain-containing protein, expressed |
| C + BR-N | qSDW10-1 | 10 | LOC_Os10g04590 | 2,185,226–2,188,547 | OsFBX358, F-box domain-containing protein, expressed |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Z.; Yao, J.; Cheng, Y.; Zhang, W.; Xu, Z.; Li, M.; Huang, J.; Ma, D.; Zhao, M. Identification of QTL under Brassinosteroid-Combined Cold Treatment at Seedling Stage in Rice Using Genotyping-by-Sequencing (GBS). Plants 2022, 11, 2324. https://doi.org/10.3390/plants11172324
Guo Z, Yao J, Cheng Y, Zhang W, Xu Z, Li M, Huang J, Ma D, Zhao M. Identification of QTL under Brassinosteroid-Combined Cold Treatment at Seedling Stage in Rice Using Genotyping-by-Sequencing (GBS). Plants. 2022; 11(17):2324. https://doi.org/10.3390/plants11172324
Chicago/Turabian StyleGuo, Zhifu, Jialu Yao, Yishan Cheng, Wenzhong Zhang, Zhengjin Xu, Maomao Li, Jing Huang, Dianrong Ma, and Minghui Zhao. 2022. "Identification of QTL under Brassinosteroid-Combined Cold Treatment at Seedling Stage in Rice Using Genotyping-by-Sequencing (GBS)" Plants 11, no. 17: 2324. https://doi.org/10.3390/plants11172324
APA StyleGuo, Z., Yao, J., Cheng, Y., Zhang, W., Xu, Z., Li, M., Huang, J., Ma, D., & Zhao, M. (2022). Identification of QTL under Brassinosteroid-Combined Cold Treatment at Seedling Stage in Rice Using Genotyping-by-Sequencing (GBS). Plants, 11(17), 2324. https://doi.org/10.3390/plants11172324