
Protective Function of Malus baccata (L.) Borkh Methanol Extract against UVB/Hydrogen Peroxide-Induced Skin Aging via Inhibition of MAPK and NF-κB Signaling

 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Antioxidant Effect of Mb-ME
2.2. Anti-Photoaging Effect of Mb-ME against UVB Irradiation in HaCaT Cells
2.3. Effect of Mb-ME on H2O2-Induced Damage in HaCaT Cells
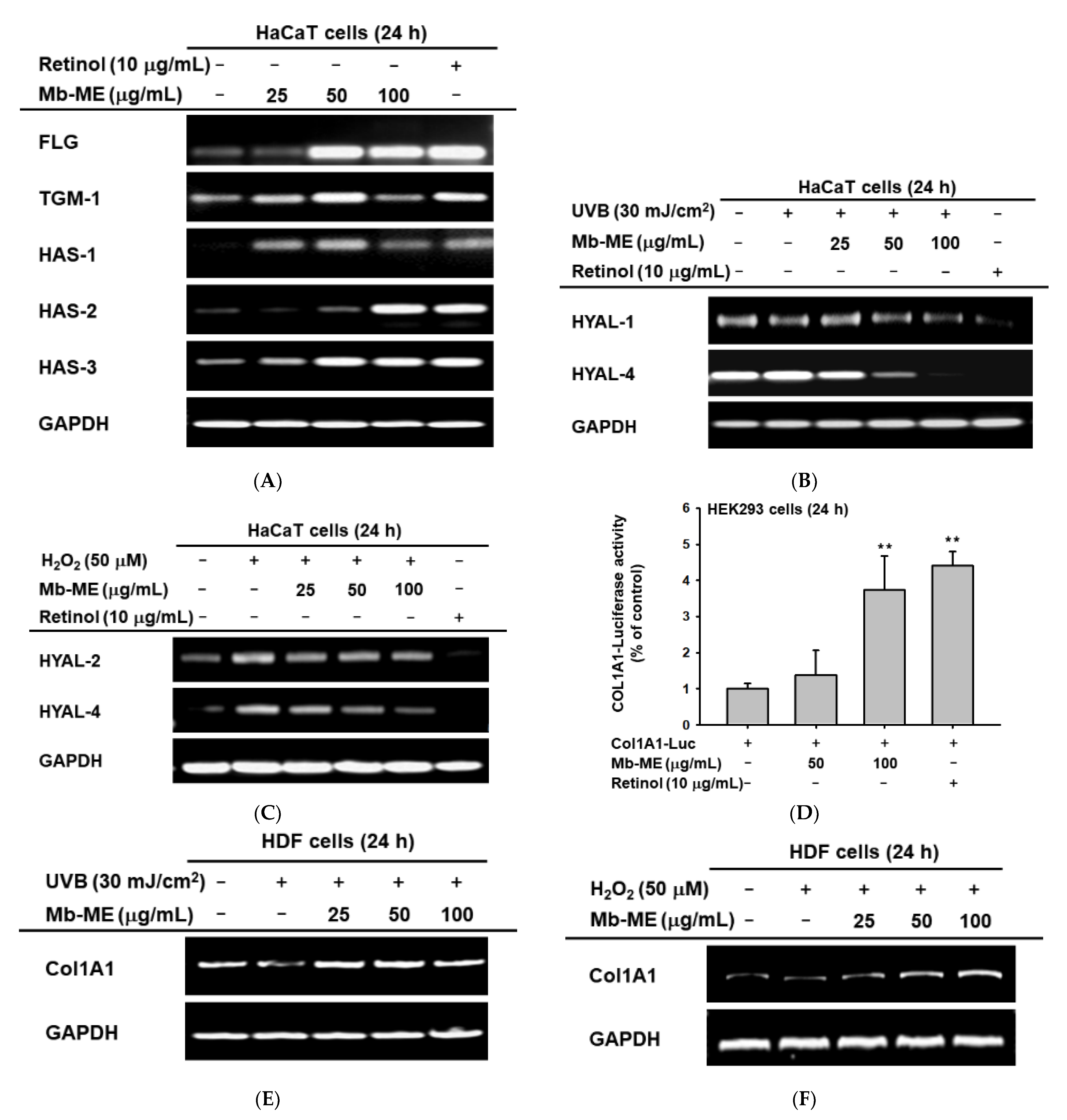
2.4. Mb-ME Promoted Moisture Retention and Collagen Synthesis
2.5. Regulatory Mechanisms of Mb-ME on UVB-Mediated HaCaT Cells
2.6. Anti-Apoptotic Effects of Mb-ME in HaCaT Cells
2.7. Effect of Mb-ME on Melanogenesis
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Plant Material and Extract Preparation
4.3. Cell Culture
4.4. Cell Viability Assay
4.5. DPPH Assay
4.6. ABTS Assay
4.7. UVB and H2O2 Treatment
4.8. ROS Generation
4.9. Gas Chromatography-Mass Spectrometry
4.10. Semi-Quantitative RT-PCR
4.11. PI and Annexin V-FITC Staining
4.12. DAPI Staining
4.13. Plasmid Transfection and Luciferase Reporter Assay
4.14. Protein Lysates Preparation and Immunoblotting
4.15. Tyrosinase Activity Assay
4.16. Melanin Formation and Secretion Analysis
4.17. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Mb-ME | Malus baccata methanol extract |
| AP-1 | Activator protein 1 |
| NF-κB | Nuclear factor-κB |
| ROS | Reactive oxygen species |
| MMP | Matrix metalloproteinases |
| RT-PCR | Reverse transcription polymerase chain reaction |
| IL | Interleukin |
| HA | Hyaluronan or hyaluronic acid |
| HYAL | Hyaluronidases |
| COX2 | Cyclooxygenase |
| MAPK | Mitogen-activated protein kinases |
| α-MSH | α-Melanocyte-stimulating hormone |
| ECM | Extracellular matrix |
References
- You, L.; Cho, J.Y. The regulatory role of Korean ginseng in skin cells. J. Ginseng Res. 2021, 45, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-O.; Kim, E.; Kim, J.H.; Hong, Y.H.; Kim, H.G.; Jeong, D.; Kim, J.; Kim, S.H.; Park, C.; Seo, D.B.; et al. Antimelanogenesis and skin-protective activities of Panax ginseng calyx ethanol extract. J. Ginseng Res. 2018, 42, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Jeong, D.; Qomaladewi, N.P.; Lee, J.; Park, S.H.; Cho, J.Y. The role of autophagy in skin fibroblasts, keratinocytes, melanocytes, and epidermal stem cells. J. Investig. Dermatol. 2020, 140, 1691–1697. [Google Scholar] [CrossRef] [PubMed]
- D’Orazio, J.; Jarrett, S.; Amaro-Ortiz, A.; Scott, T. UV radiation and the skin. Int. J. Mol. Sci. 2013, 14, 12222–12248. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.; Bhattarai, P.Y.; Lim, S.C.; Kim, J.Y.; Choi, H.S. PIN1 facilitates ubiquitin-mediated degradation of serine/threonine kinase 3 and promotes melanoma development via TAZ activation. Cancer Lett. 2021, 499, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Jang, J.; Kopalli, S.R.; Yum, J.; Yoon, K.; Cho, J.Y. Anti-photoaging activities of Sorbaria kirilowii ethanol extract in UVB-damaged cells. Cytotechnology 2021, 73, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.F.; Wang, Y.Y.; Hsiao, M.; Lo, S.; Chang, Y.C.; Jan, Y.H.; Lai, T.C.; Lee, Y.C.; Hsieh, Y.C.; Yuan, S.F. IMPAD1 functions as mitochondrial electron transport inhibitor that prevents ROS production and promotes lung cancer metastasis through the AMPK-Notch1-HEY1 pathway. Cancer Lett. 2020, 485, 27–37. [Google Scholar] [CrossRef]
- Paz, M.L.; González Maglio, D.H.; Weill, F.S.; Bustamante, J.; Leoni, J. Mitochondrial dysfunction and cellular stress progression after ultraviolet B irradiation in human keratinocytes. Photodermatol. Photoimmunol. Photomed. 2008, 24, 115–122. [Google Scholar] [CrossRef]
- Yang, K.; Cao, F.; Xue, Y.; Tao, L.; Zhu, Y. Three classes of antioxidant defense systems and the development of postmenopausal osteoporosis. Front. Physiol. 2022, 13, 840293. [Google Scholar] [CrossRef]
- Jiang, L.; Chen, Y.; Min, G.; Wang, J.; Chen, W.; Wang, H.; Wang, X.; Yao, N. Bcl2-associated athanogene 4 promotes the invasion and metastasis of gastric cancer cells by activating the PI3K/AKT/NF-kappaB/ZEB1 axis. Cancer Lett. 2021, 520, 409–421. [Google Scholar] [CrossRef]
- Lee, M.G.; Lee, K.S.; Nam, K.S. Anti-metastatic effects of arctigenin are regulated by MAPK/AP-1 signaling in 4T-1 mouse breast cancer cells. Mol. Med. Rep. 2020, 21, 1374–1382. [Google Scholar] [CrossRef]
- Cavdar, Z.; Ozbal, S.; Celik, A.; Ergur, B.U.; Guneli, E.; Ural, C.; Camsari, T.; Guner, G.A. The effects of alpha-lipoic acid on MMP-2 and MMP-9 activities in a rat renal ischemia and re-perfusion model. Biotech. Histochem. 2014, 89, 304–314. [Google Scholar] [CrossRef]
- Kawada, C.; Yoshida, T.; Yoshida, H.; Matsuoka, R.; Sakamoto, W.; Odanaka, W.; Sato, T.; Yamasaki, T.; Kanemitsu, T.; Masuda, Y.; et al. Ingested hyaluronan moisturizes dry skin. Nutr. J. 2014, 13, 70. [Google Scholar] [CrossRef]
- Weigel, P.H.; Hascall, V.C.; Tammi, M. Hyaluronan synthases. J. Biol. Chem. 1997, 272, 13997–14000. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Hearing, V.J. Melanocytes and their diseases. Cold Spring Harb. Perspect. Med. 2014, 4, a017046. [Google Scholar] [CrossRef]
- Jiang, L.; Huang, J.; Hu, Y.; Lei, L.; Ouyang, Y.; Long, Y.; Li, H.; Li, S.; Yang, L.; Yang, Y.; et al. Identification of the ceRNA networks in alpha-MSH-induced melanogenesis of melanocytes. Aging 2020, 13, 2700–2726. [Google Scholar] [CrossRef]
- Park, S.H.; Choi, E.; Kim, S.; Kim, D.S.; Kim, J.H.; Chang, S.; Choi, J.S.; Park, K.J.; Roh, K.B.; Lee, J.; et al. Oxidative stress-protective and anti-melanogenic effects of loliolide and ethanol extract from fresh water green algae, Prasiola japonica. Int. J. Mol. Sci. 2018, 19, 2825. [Google Scholar] [CrossRef]
- Kim, J.H.; Lee, J.E.; Kim, T.; Yeom, M.H.; Park, J.S.; di Luccio, E.; Chen, H.; Dong, Z.; Lee, K.W.; Kang, N.J. 7,3′,4′-Trihydroxyisoflavone, a metabolite of the soy isoflavone daidzein, suppresses α-melanocyte-stimulating hormone-induced melanogenesis by targeting melanocortin 1 receptor. Front. Mol. Biosci. 2020, 7, 577284. [Google Scholar] [CrossRef]
- Floegel, A.; Kim, D.-O.; Chung, S.-J.; Koo, S.I.; Chun, O.K. Comparison of ABTS/DPPH assays to measure antioxidant capacity in popular antioxidant-rich US foods. J. Food Composit. Anal. 2011, 24, 1043–1048. [Google Scholar] [CrossRef]
- Movalia, D.; Mishra, S.; Gajera, F. Free radical scavenging activity of Randia dumetorum Lamk. fruits. Int. Res. J. Pharm. 2010, 1, 122–126. [Google Scholar]
- Oberhammer, F.A.; Hochegger, K.; Fröschl, G.; Tiefenbacher, R.; Pavelka, M. Chromatin condensation during apoptosis is accompanied by degradation of lamin A+B, without enhanced activation of cdc2 kinase. J. Cell Biol. 1994, 126, 827–837. [Google Scholar] [CrossRef] [PubMed]
- Fisher, G.J.; Datta, S.C.; Talwar, H.S.; Wang, Z.Q.; Varani, J.; Kang, S.; Voorhees, J.J. Molecular basis of sun-induced premature skin ageing and retinoid antagonism. Nature 1996, 379, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Lorz, L.R.; Lee, J.; Cho, J.Y. In vitro photoprotective, anti-inflammatory, moisturizing, and antimelanogenic effects of a methanolic extract of Chrysophyllum lucentifolium Cronquist. Plants 2021, 11, 94. [Google Scholar] [CrossRef]
- Piérard-Franchimont, C.; Castelli, D.; Cromphaut, I.V.; Bertin, C.; Ries, G.; Cauwenbergh, G.; Piérard, G.E. Tensile properties and contours of aging facial skin. A controlled double-blind comparative study of the effects of retinol, melibiose-lactose and their association. Skin Res. Technol. 1998, 4, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Todd, V.M.; Johnson, R.W. Hypoxia in bone metastasis and osteolysis. Cancer Lett. 2020, 489, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Jin, G.-H.; Liu, Y.; Jin, S.-Z.; Liu, X.-D.; Liu, S.-Z. UVB induced oxidative stress in human keratinocytes and protective effect of antioxidant agents. Radiat. Environ. Biophys. 2007, 46, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Ryser, S.; Schuppli, M.; Gauthier, B.; Hernandez, D.R.; Roye, O.; Hohl, D.; German, B.; Holzwarth, J.A.; Moodycliffe, A.M. UVB-induced skin inflammation and cutaneous tissue injury is dependent on the MHC class I-like protein, CD1d. J. Investig. Dermatol. 2014, 134, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.O.; Chun, O.K.; Kim, Y.J.; Moon, H.Y.; Lee, C.Y. Quantification of polyphenolics and their antioxidant capacity in fresh plums. J. Agric. Food Chem. 2003, 51, 6509–6515. [Google Scholar] [CrossRef]
- Munteanu, I.G.; Apetrei, C. Analytical methods used in determining antioxidant activity: A review. Int. J. Mol. Sci. 2021, 22, 3380. [Google Scholar] [CrossRef]
- Gulcin, I. Antioxidants and antioxidant methods: An updated overview. Arch. Toxicol. 2020, 94, 651–715. [Google Scholar] [CrossRef]
- Chand, K.; Rajeshwari; Hiremathad, A.; Singh, M.; Santos, M.A.; Keri, R.S. A review on antioxidant potential of bioactive heterocycle benzofuran: Natural and synthetic derivatives. Pharmacol. Rep. 2017, 69, 281–295. [Google Scholar] [CrossRef]
- Aswathanarayanappa, C.; Bheemappa, E.; Bodke, Y.D.; Bhovi, V.K.; Ningegowda, R.; Shivakumar, M.C.; Peethambar, S.K.; Telkar, S. 5-phenyl-1-benzofuran-2-yl derivatives: Synthesis, antimicrobial, and antioxidant activity. Med. Chem. Res. 2013, 22, 78–87. [Google Scholar] [CrossRef]
- Desai, S.R. Hyperpigmentation therapy: A review. J. Clin. Aesthet. Dermatol. 2014, 7, 13–17. [Google Scholar]
- Qi, S.; Perrino, S.; Miao, X.; Lamarche-Vane, N.; Brodt, P. The chemokine CCL7 regulates invadopodia maturation and MMP-9 mediated collagen degradation in liver-metastatic carcinoma cells. Cancer Lett. 2020, 483, 98–113. [Google Scholar] [CrossRef]
- Cen, L.; Liu, W.; Cui, L.; Zhang, W.; Cao, Y. Collagen tissue engineering: Development of novel biomaterials and applications. Pediatr. Res. 2008, 63, 492–496. [Google Scholar] [CrossRef]
- Ruiz Martinez, M.A.; Peralta Galisteo, S.; Castan, H.; Morales Hernandez, M.E. Role of proteoglycans on skin ageing: A review. Int. J. Cosmet. Sci. 2020, 42, 529–535. [Google Scholar] [CrossRef]
- Shin, K.K.; Park, S.H.; Lim, H.Y.; Lorza, L.R.; Qomaladewia, N.P.; You, L.; Aziz, N.; Kim, S.A.; Lee, J.S.; Choung, E.S.; et al. In vitro anti-photoaging and skin protective effects of Licania macrocarpa Cuatrec methanol extract. Plants 2022, 11, 1383. [Google Scholar] [CrossRef]
- Papakonstantinou, E.; Roth, M.; Karakiulakis, G. Hyaluronic acid: A key molecule in skin aging. Dermato-Endocrinology 2012, 4, 253–258. [Google Scholar] [CrossRef]
- Kaul, A.; Short, W.D.; Wang, X.; Keswani, S.G. Hyaluronidases in human diseases. Int. J. Mol. Sci. 2021, 22, 3204. [Google Scholar] [CrossRef]
- Ishida-Yamamoto, A.; Iizuka, H. Structural organization of cornified cell envelopes and alterations in inherited skin disorders. Exp. Dermatol. 1998, 7, 1–10. [Google Scholar] [CrossRef]
- Lim, H.Y.; Jeong, D.; Park, S.H.; Shin, K.K.; Hong, Y.H.; Kim, E.; Yu, Y.-G.; Kim, T.-R.; Kim, H.; Lee, J.; et al. Antiwrinkle and antimelanogenesis effects of tyndallized Lactobacillus acidophilus KCCM12625P. Int. J. Mol. Sci. 2020, 21, 1620. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Wu, S.B.; Hong, C.H.; Yu, H.S.; Wei, Y.H. Molecular mechanisms of UV-induced apoptosis and its effects on skin residential cells: The implication in UV-based phototherapy. Int. J. Mol. Sci. 2013, 14, 6414–6435. [Google Scholar] [CrossRef] [Green Version]
- Porter, A.G.; Jänicke, R.U. Emerging roles of caspase-3 in apoptosis. Cell Death Differ. 1999, 6, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Liu, S.; Li, Y.; Wang, Y.; Deng, Y.; Sun, W.; Huang, H.; Xie, J.; He, A.; Chen, H.; et al. Pyridoxine induces monocyte-macrophages death as specific treatment of acute myeloid leukemia. Cancer Lett. 2020, 492, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Nin, G.H.; Escobar, M.L.; Echeverría, O.M. Necrosis. In Cell Death in Mammalian Ovary; Vázquez-Nin, G.H., Escobar, M.L., De Felici, M., Echeverría, O.M., Klinger, F.G., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 111–121. [Google Scholar]
- Rajamohan, S.B.; Pillai, V.B.; Gupta, M.; Sundaresan, N.R.; Birukov, K.G.; Samant, S.; Hottiger, M.O.; Gupta, M.P. SIRT1 promotes cell survival under stress by deacetylation-dependent deactivation of poly(ADP-ribose) polymerase 1. Mol. Cell Biol. 2009, 29, 4116–4129. [Google Scholar] [CrossRef] [PubMed]
- Vieyra-Garcia, P.A.; Wolf, P. A deep dive into UV-based phototherapy: Mechanisms of action and emerging molecular targets in inflammation and cancer. Pharmacol. Ther. 2021, 222, 107784. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Chen, H.; Kim, S.A.; Lee, J.S.; Choung, E.S.; Zhang, Z.; Kim, S.Y.; Kim, J.H.; Cho, J.Y. Anti-inflammatory functions of methanol extract from Malus baccata (L.) Borkh. leaves and shoots by targeting the NF-κB pathway. Plants 2022, 11, 646. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Jeong, D.; Hong, Y.H.; Li, W.Y.; Lee, S.W.; Hossain, M.A.; Taamalli, A.; Kim, J.H.; Kim, J.-H.; Cho, J.Y. Anti-inflammatory and photoaging-protective effects of Olea europaea through inhibition of AP-1 and NF-κ B pathways. Am. J. Chin. Med. 2020, 48, 1895–1913. [Google Scholar] [CrossRef]
- Zappavigna, S.; Cossu, A.M.; Abate, M.; Misso, G.; Lombardi, A.; Caraglia, M.; Filosa, R. A hydroquinone-based derivative elicits apoptosis and autophagy via activating a ROS-dependent unfolded protein response in human glioblastoma. Int. J. Mol. Sci. 2019, 20, 3836. [Google Scholar] [CrossRef]
- Lee, J.O.; Hwang, S.H.; Shen, T.; Kim, J.H.; You, L.; Hu, W.; Cho, J.Y. Enhancement of skin barrier and hydration-related molecules by protopanaxatriol in human keratinocytes. J. Ginseng Res. 2021, 45, 354–360. [Google Scholar] [CrossRef]
- Kim, E.; Hwang, K.; Lee, J.; Han, S.Y.; Kim, E.-M.; Park, J.; Cho, J.Y. Skin protective effect of epigallocatechin gallate. Int. J. Mol. Sci. 2018, 19, 173. [Google Scholar] [CrossRef]
- Chen, Z.W.; Liu, A.; Liu, Q.; Chen, J.; Li, W.M.; Chao, X.J.; Yang, Q.; Liu, P.Q.; Mao, Z.X.; Pi, R.B. MEF2D mediates the neuroprotective effect of methylene blue against glutamate-induced oxidative damage in HT22 hippocampal cells. Mol. Neurobiol. 2017, 54, 2209–2222. [Google Scholar] [CrossRef]
- Wang, S.F.; Chang, Y.L.; Tzeng, Y.D.; Wu, C.L.; Wang, Y.Z.; Tseng, L.M.; Chen, S.; Lee, H.C. Mitochondrial stress adaptation promotes resistance to aromatase inhibitor in human breast cancer cells via ROS/calcium up-regulated amphiregulin-estrogen receptor loop signaling. Cancer Lett. 2021, 523, 82–99. [Google Scholar] [CrossRef]
- Hwang, S.H.; Kim, J.H.; Choi, E.; Park, S.H.; Cho, J.Y. Antioxidative and skin protective effects of Canarium subulatum methanol extract on keratinocytes. Evid. Based Complement. Alternat. Med. 2021, 2021, 6692838. [Google Scholar] [CrossRef]
- Chen, Y.J.; Wu, J.Y.; Deng, Y.Y.; Wu, Y.; Wang, X.Q.; Li, A.S.; Wong, L.Y.; Fu, X.Q.; Yu, Z.L.; Liang, C. Ginsenoside Rg3 in combination with artesunate overcomes sorafenib resistance in hepatoma cell and mouse models. J. Ginseng Res. 2022, 46, 418–425. [Google Scholar] [CrossRef]
- El Sayed, A.M.; Basam, S.M.; El-Naggar, E.-M.b.A.; Marzouk, H.S.; El-Hawary, S. LC–MS/MS and GC–MS profiling as well as the antimicrobial effect of leaves of selected Yucca species introduced to Egypt. Sci. Rep. 2020, 10, 17778. [Google Scholar] [CrossRef]
- Lorz, L.R.; Yoo, B.C.; Kim, M.Y.; Cho, J.Y. Anti-wrinkling and anti-melanogenic effect of Pradosia mutisii methanol extract. Int. J. Mol. Sci. 2019, 20, 1043. [Google Scholar] [CrossRef]
- Lee, J.O.; Kim, J.H.; Kim, S.; Kim, M.Y.; Hong, Y.H.; Kim, H.G.; Cho, J.Y. Gastroprotective effects of the nonsaponin fraction of Korean Red Ginseng through cyclooxygenase-1 upregulation. J. Ginseng Res. 2020, 44, 655–663. [Google Scholar] [CrossRef]
- Jungwirth, G.; Yu, T.; Cao, J.; Eddine, M.A.; Moustafa, M.; Warta, R.; Debus, J.; Unterberg, A.; Abdollahi, A.; Herold-Mende, C. KIF11 inhibitors filanesib and ispinesib inhibit meningioma growth in vitro and in vivo. Cancer Lett 2021, 506, 1–10. [Google Scholar] [CrossRef]
- Choi, E.; Kim, E.; Kim, J.H.; Yoon, K.; Kim, S.; Lee, J.; Cho, J.Y. AKT1-targeted proapoptotic activity of compound K in human breast cancer cells. J. Ginseng Res. 2019, 43, 692–698. [Google Scholar] [CrossRef]
- Bisht, S.; Khan, M.A.; Bekhit, M.; Bai, H.; Cornish, T.; Mizuma, M.; Rudek, M.A.; Zhao, M.; Maitra, A.; Ray, B.; et al. A polymeric nanoparticle formulation of curcumin (NanoCurc™) ameliorates CCl4-induced hepatic injury and fibrosis through reduction of pro-inflammatory cytokines and stellate cell activation. Lab. Investig. 2011, 91, 1383–1395. [Google Scholar] [CrossRef]
- Hunto, S.T.; Shin, K.K.; Kim, H.G.; Park, S.H.; Oh, J.; Sung, G.-H.; Hossain, M.A.; Rho, H.S.; Lee, J.; Kim, J.-H.; et al. Phosphatidylinositide 3-kinase contributes to the anti-inflammatory effect of Abutilon crispum L. Medik methanol extract. Evid. Based Complement. Alternat. Med. 2018, 2018, 1935902. [Google Scholar] [CrossRef] [Green Version]
- Ma, D.; Zhan, D.; Fu, Y.; Wei, S.; Lal, B.; Wang, J.; Li, Y.; Lopez-Bertoni, H.; Yalcin, F.; Dzaye, O.; et al. Mutant IDH1 promotes phagocytic function of microglia/macrophages in gliomas by downregulating ICAM1. Cancer Lett. 2021, 517, 35–45. [Google Scholar] [CrossRef]
- Jeong, D.; Lee, J.; Park, S.H.; Kim, Y.A.; Park, B.J.; Oh, J.; Sung, G.-H.; Aravinthan, A.; Kim, J.-H.; Kang, H.; et al. Antiphotoaging and antimelanogenic effects of Penthorum chinense Pursh ethanol extract due to antioxidant- and autophagy-inducing properties. Oxid. Med. Cell. Longev. 2019, 2019, 9679731. [Google Scholar] [CrossRef]
- Jeong, D.; Lee, J.; Jeong, S.-G.; Hong, Y.H.; Yoo, S.; Han, S.Y.; Kim, J.H.; Kim, S.; Kim, J.S.; Chung, Y.S.; et al. Artemisia asiatica ethanol extract exhibits anti-photoaging activity. J. Ethnopharmacol. 2018, 220, 57–66. [Google Scholar] [CrossRef]
- Lim, H.Y.; Kim, E.; Park, S.H.; Hwang, K.H.; Kim, D.; Jung, Y.J.; Kopalli, S.R.; Hong, Y.D.; Sung, G.H.; Cho, J.Y. Antimelanogenesis effects of theasinensin A. Int. J. Mol. Sci. 2021, 22, 7453. [Google Scholar] [CrossRef]
- Lee, J.Y.; Cho, Y.-R.; Park, J.H.; Ahn, E.-K.; Jeong, W.; Shin, H.S.; Kim, M.-S.; Yang, S.H.; Oh, J.S. Anti-melanogenic and anti-oxidant activities of ethanol extract of Kummerowia striata: Kummerowia striata regulate anti-melanogenic activity through down-regulation of TRP-1, TRP-2 and MITF expression. Toxicol. Rep. 2019, 6, 10–17. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | R.T. (min) | Compound Name | Peak Area % |
|---|---|---|---|
| 1 | 8.500 | 2-Propanone | 4.212 |
| 2 | 10.125 | 1-Hydroxy-2-butanone, 2-Propenoic acid, 2-hydroxyethyl ester | 2.007 |
| 3 | 10.499 | Hexanal, Butanal | 0.531 |
| 4 | 10.625 | 1-Propanol,3-Isopropoxy alanine, Butyric acid hydrazide | 0.868 |
| 5 | 10.780 | Di-n-propyl ether, Propanoic acid, Methyl ester | 1.038 |
| 6 | 11.696 | Furfural | 0.401 |
| 7 | 12.192 | 2-Furanmethanol | 1.597 |
| 8 | 13.797 | 2-Heptenal, (Z)-2-Heptenal, (E)-Cyclopentane | 0.732 |
| 9 | 13.959 | 2,4-Dihydroxy-2,5-dimethyl-3(2H)-furan-3-one | 0.557 |
| 10 | 15.074 | Phenol | 5.569 |
| 11 | 15.755 | 2-Butylamine, N-pentyl-Thiophene-3 | 1.723 |
| 12 | 16.413 | 1,3,5-Triazine-2,4,6-triamine,monoamide,propargyl ester | 7.152 |
| 13 | 17.717 | 4H-Pyran-4-one, 2,3-dihydro-3,5-dihydroxy-6-methyl- | 5.802 |
| 14 | 18.106 | Methyl salicylate | 7.156 |
| 15 | 19.104 | Benzofuran | 20.304 |
| 16 | 19.551 | 5-Hydroxymethylfurfural | 6.290 |
| 17 | 20.092 | 2-Methoxy-4-vinylphenol | 3.622 |
| 18 | 20.512 | Hydroquinone, Isosorbide | 11.142 |
| 19 | 25.288 | N-m-Tolyl-succinamic acid, 2-Butanone, 4-(4-hydroxyphenyl)-Acetamide | 4.232 |
| 20 | 27.565 | Benzeneacetic acid,4-hydroxy-Benzeneacetic acid, methyl ester | 9.241 |
| 21 | 28.281 | piperidine, 4-(4-methoxyphenoxy)- | 4.235 |
| Gene Name | Sequencing (5′ to 3′) | |
|---|---|---|
| FLG | F | AGGGAAGATCCAAGAGCCCA |
| R | ACTCTGGATCCCCTACGCTT | |
| MMP-3 | F | ATCCTACTGTTGCTGTGCGT |
| R | CATCACCTCCAGAGTGTCGG | |
| HAS-3 | F | GTTCGCGGCTGCTTTGAC |
| R | GTAGCCCGTCACATAGGCTG | |
| GAPDH | F | GGTCACCAGGGCTGCTTTTA |
| R | GATGGCATGGACTGTGGTCA | |
| Sirt1 | F | CAGTGTCATGGTTCCTTTGC |
| R | CACCGAGGAACTACCTGAT | |
| HAS-1 | F | CCACCCAGTACAGCGTCAAC |
| R | CATGGTGCTTCTGTCGCTCT | |
| COX-2 | F | CAAAAGCTGGGAAGCCTTCT |
| R | CCATCCTTCAAAAGGCGCAG | |
| MMP-2 | F | ACGACCGCGACAAGAAGTAT |
| R | CTGCAAAGAACACAGCCTTCTC | |
| TGM1 | F | GAAATGCGGCAGATGACGAC |
| R | AACTCCCCAGCGTCTGATTG | |
| IL-6 | F | TTCGGTCCAGTTGCCTTCTCC |
| R | TGAGGTGCCCATGCTACATTT | |
| HYAL-2 | F | TACACCACAAGCACGGAGAC |
| R | ATGCAGGAAGGTACTGGCAC | |
| MMP-9 | F | CAACATCACCTATTGGATCC |
| R | CGGGTGTAGAGTCTCTCGCT | |
| HYAL-1 | F | CAGAATGCCAGCCTGATTGC |
| R | CCGGTGTAGTTGGGGCTTAG | |
| p53 | F | CAGCCAAGTCTGTGACTTGCACGTAC |
| R | CTATGTCGAAAAGTGTTTCTGTCATC | |
| HYAL-4 | F | TGAGCTCTCTTGGCTCTGGA |
| R | AGGCAGCACTTTCTCCTATGG | |
| COL1A1 | F | CAGGTACCATGACCGAGACG |
| R | AGCACCATCATTTCCACGAG | |
| IL-1β | F | TGAGCTCGCCAGTGAAATGA |
| R | AACACGCAGGACAGGTACAG | |
| HAS-2 | F | TTCTTTATGTGACTCATCTGTCTCACCGG |
| R | ATTGTTGGCTACCAGTTTATCCAAACG |
| Gene Name | Sequencing (5′ to 3′) | |
|---|---|---|
| MITF | F | GGGAGCTCACAGCGTGTATT |
| R | CTAGCCTGCATCTCCAGCTC | |
| TYRP1 | F | GTGAGCAGCTCTGTGCTGTA |
| R | AGGGGGAGGACGTTGTAAGA | |
| Tyrosinase | F | GGCCAGCTTTCAGGCAGAGG |
| R | TGGTGCTTCATGGGCAAAAT | |
| GAPDH | F | ACCCTTAAGAGGGATGCTGC |
| R | GTTCACACCGACCTTCACCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, C.; Lee, C.Y.; Lee, H.P.; Hossain, M.A.; Zhang, Z.; Kim, S.-Y.; Song, M.; Kim, J.-H.; Cho, J.Y. Protective Function of Malus baccata (L.) Borkh Methanol Extract against UVB/Hydrogen Peroxide-Induced Skin Aging via Inhibition of MAPK and NF-κB Signaling. Plants 2022, 11, 2368. https://doi.org/10.3390/plants11182368
Song C, Lee CY, Lee HP, Hossain MA, Zhang Z, Kim S-Y, Song M, Kim J-H, Cho JY. Protective Function of Malus baccata (L.) Borkh Methanol Extract against UVB/Hydrogen Peroxide-Induced Skin Aging via Inhibition of MAPK and NF-κB Signaling. Plants. 2022; 11(18):2368. https://doi.org/10.3390/plants11182368
Chicago/Turabian StyleSong, Chaoran, Chae Young Lee, Hwa Pyoung Lee, Mohammad Amjad Hossain, Zhiyun Zhang, Soo-Yong Kim, Minkyung Song, Jong-Hoon Kim, and Jae Youl Cho. 2022. "Protective Function of Malus baccata (L.) Borkh Methanol Extract against UVB/Hydrogen Peroxide-Induced Skin Aging via Inhibition of MAPK and NF-κB Signaling" Plants 11, no. 18: 2368. https://doi.org/10.3390/plants11182368
APA StyleSong, C., Lee, C. Y., Lee, H. P., Hossain, M. A., Zhang, Z., Kim, S. -Y., Song, M., Kim, J. -H., & Cho, J. Y. (2022). Protective Function of Malus baccata (L.) Borkh Methanol Extract against UVB/Hydrogen Peroxide-Induced Skin Aging via Inhibition of MAPK and NF-κB Signaling. Plants, 11(18), 2368. https://doi.org/10.3390/plants11182368