
Evidence of Asexual Overwintering of Melampsora paradoxa and Mapping of Stem Rust Host Resistance in Salix


Abstract
:1. Introduction
2. Results
2.1. Melampsora paradoxa Isolate Collection
2.2. GBS Sequencing and SNP Analysis
2.3. Field Evaluation of Leaf Rust and Stem Canker
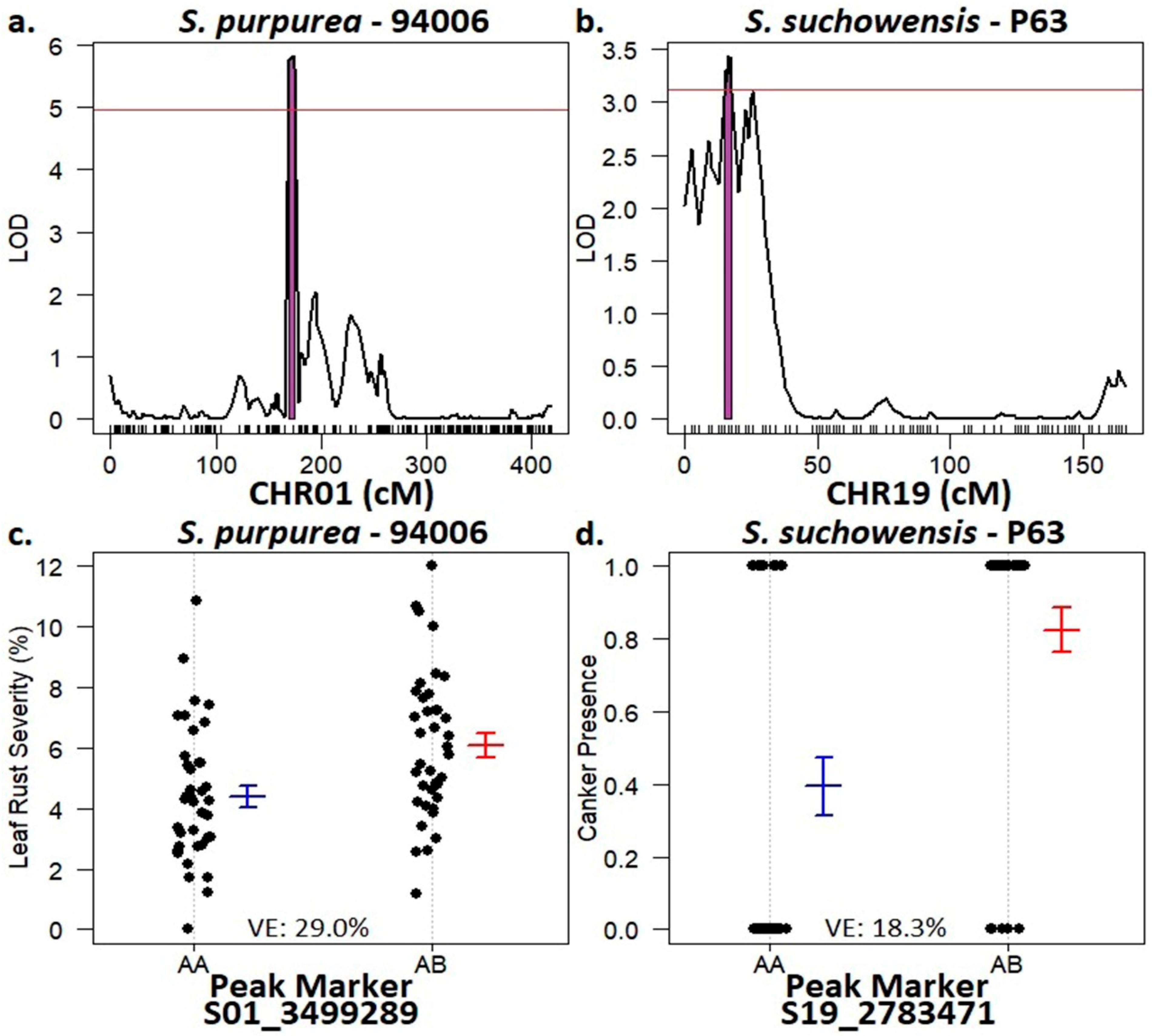
2.4. Host Resistance Mapping
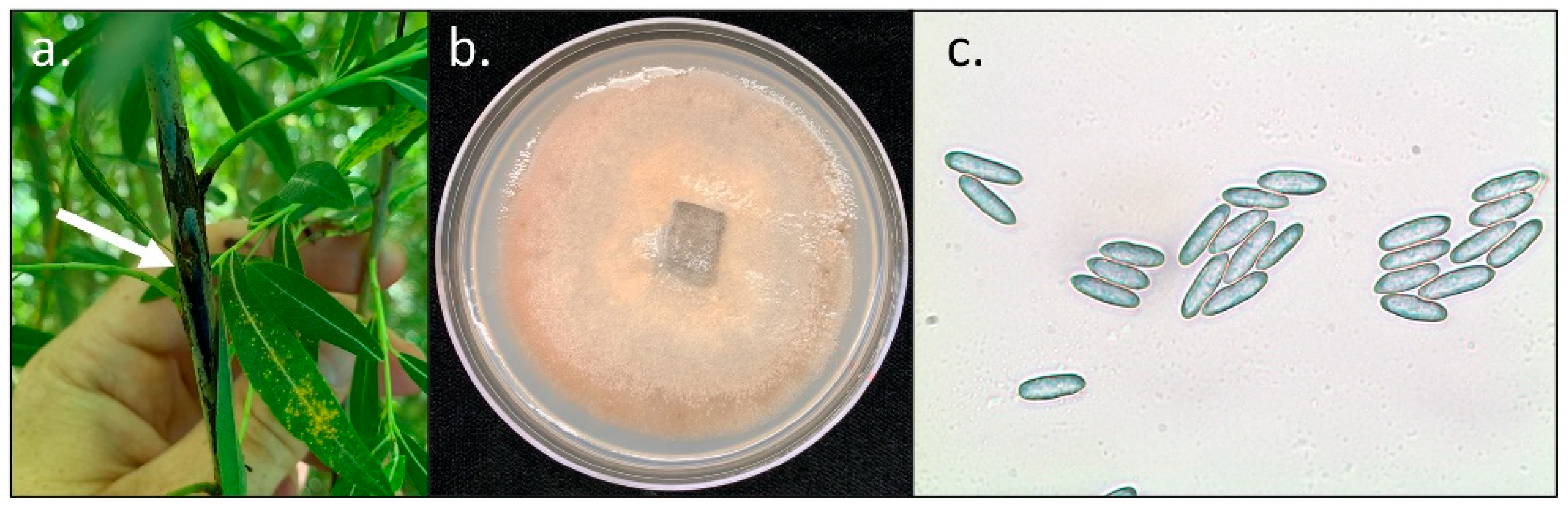
2.5. Recovery of M. paradoxa from Dormant Salix Stems after Overwintering
3. Discussion
4. Materials and Methods
4.1. Melampsora paradoxa Isolate Collection and GBS Genotyping
4.2. SNP Analysis
4.3. Determination of Clonality
4.4. Leaf Rust and Stem Canker Rust Disease Evaluation
4.5. Host Resistance QTL Mapping
4.6. Recovery of M. paradoxa from Dormant Salix Stems after Overwintering
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clifton-Brown, J.; Harfouche, A.; Casler, M.; Jones, H.D.; Macalpine, W.J.; Murphy-Bokern, D.; Smart, L.B.; Adler, A.; Ashman, C.; Awty-Carroll, D.; et al. Breeding progress and preparedness for mass upscaling of perennial lignocellulosic biomass crops switchgrass, miscanthus, willow and poplar. GCB Bioenergy 2019, 11, 118–151. [Google Scholar] [CrossRef] [PubMed]
- Volk, T.A.; Berguson, B.; Daly, C.; Halbleib, M.; Miller, R.; Rials, T.; Abrahamson, L.P.; Buchman, D.; Cunningham, M.; Eisenbies, M.; et al. Poplar and shrub willow energy crops in the United States: Field trial results from the multiyear Regional Feedstock Partnership and yield potential maps based on the PRISM-ELM model. GCB Bioenergy 2018, 10, 735–751. [Google Scholar] [CrossRef]
- Smart, L.B.; Cameron, K.D. Shrub willow. In Handbook of Bioenergy Crop Plants; Kole, C., Joshi, C.P., Shonnard, D.R., Eds.; Taylor and Francis Group: Boca Raton, FL, USA, 2012; pp. 687–708. [Google Scholar]
- Stoof, C.R.; Richards, B.K.; Woodbury, P.B.; Fabio, E.S.; Brumbach, A.R.; Cherney, J.; Das, S.; Geohring, L.; Hansen, J.; Hornesky, J.; et al. Untapped potential: Opportunities and challenges for sustainable bioenergy production from marginal lands in the northeast USA. Bioenergy Res. 2015, 8, 482–501. [Google Scholar] [CrossRef]
- Fabio, E.S.; Volk, T.A.; Miller, R.O.; Serapiglia, M.J.; Gauch, H.G.; Van Rees, K.C.J.; Hangs, R.D.; Amichev, B.Y.; Kuzovkina, Y.A.; Labrecque, M.; et al. Genotype × environment interaction analysis of North American shrub willow yield trials confirms superior performance of triploid hybrids. GCB Bioenergy 2017, 9, 445–459. [Google Scholar] [CrossRef]
- Crowell, C.R.; Bekauri, M.M.; Cala, A.R.; McMullen, P.; Smart, L.B.; Smart, C.D. Differential susceptibility of diverse Salix spp. to Melampsora americana and Melampsora paradoxa. Plant Dis. 2020, 104, 2949–2957. [Google Scholar] [CrossRef]
- Kenaley, S.C.; Smart, L.B.; Hudler, G.W. Genetic evidence for three discrete taxa of Melampsora (Pucciniales) affecting willows (Salix spp.) in New York State. Fungal Biol. 2014, 118, 704–720. [Google Scholar] [CrossRef]
- Crowell, C.R.; Wilkerson, D.W.; Beckauri, M.; Cala, A.R.; McMullen, P.W.; Mondo, S.; Andreopoulos, W.; Lipzen, A.; Lail, K.; Yan, M.; et al. The Melampsora americana population on Salix purpurea in the Great Lakes Region is highly diverse with a contributory influence of clonality. Phytopathology 2022, 112, 907–916. [Google Scholar] [CrossRef]
- Wilkerson, D.G.; Crowell, C.R.; Carlson, C.H.; McMullen, P.W.; Smart, C.D.; Smart, L.B. Comparative transcriptomics and eQTL mapping of response to Melampsora americana in selected Salix purpurea F2 progeny. BMC Genom. 2022, 23, 71. [Google Scholar] [CrossRef]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A robust, simple genotyping-by-sequencing (GBS) approach for high diversity species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef]
- Carlson, C.H.; Gouker, F.E.; Crowell, C.R.; Evans, L.; Difazio, S.P.; Smart, C.D.; Smart, L.B. Joint linkage and association mapping of complex traits in shrub willow (Salix purpurea L.). Ann. Bot. 2019, 124, 701–715. [Google Scholar] [CrossRef] [Green Version]
- Wilkerson, D.G.; Taskiran, B.; Carlson, C.H.; Smart, L.B. Mapping the sex determination region in the Salix F1 hybrid common parent population confirms a ZW system in six diverse species. G3 2022, 12, jkac071. [Google Scholar] [CrossRef]
- Vogel, G.; Gore, M.A.; Smart, C.D. Genome-wide association study in New York Phytophthora capsici isolates reveals loci involved in mating type and mefenoxam sensitivity. Phytopathology 2021, 111, 204–216. [Google Scholar] [CrossRef]
- Torkamaneh, D.; Laroche, J.; Belzile, F. Genome-wide SNP calling from genotyping by sequencing (GBS) data: A comparison of seven pipelines and two sequencing technologies. PLoS ONE 2016, 11, e0161333. [Google Scholar] [CrossRef]
- Melo, A.T.O.; Hale, I. Genetics and population analysis expanded functionality, increased accuracy, and enhanced speed in the de novo genotyping- by-sequencing pipeline GBS-SNP-CROP. Bioinformatics 2019, 35, 1783–1785. [Google Scholar] [CrossRef]
- Pei, M.H.; Royle, D.J.; Hunter, T. A comparative study of stem- and leaf-infecting forms of Melampsora rust on Salix viminalis in the U.K. Mycol. Res. 1995, 99, 357–363. [Google Scholar] [CrossRef]
- Pei, M.H.; Ruiz, C. AFLP evidence of the distinctive patterns of life-cycle in two forms of Melampsora rust on Salix viminalis. Mycol. Res. 2000, 104, 937–942. [Google Scholar] [CrossRef]
- Kenaley, S.C.; Hudler, G.W.; O’Brien, D.D.; Cameron, K.D.; Smart, L.B. Willowpedia: Black Canker. 2011. Available online: https://cpb-us-e1.wpmucdn.com/blogs.cornell.edu/dist/5/4999/files/2014/10/Black-Canker-fact-sheet-2011_07_22-vmud3y.pdf (accessed on 29 June 2021).
- Serapiglia, M.J.; Cameron, K.D.; Stipanovic, A.J.; Abrahamson, L.P.; Volk, T.A.; Smart, L.B. Yield and woody biomass traits of novel shrub willow hybrids at two contrasting sites. Bioenergy Res. 2013, 6, 533–546. [Google Scholar] [CrossRef]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef]
- Glaubitz, J.C.; Casstevens, T.M.; Lu, F.; Harriman, J.; Elshire, R.J.; Sun, Q.; Buckler, E.S. TASSEL-GBS: A high capacity genotyping by sequencing analysis pipeline. PLoS ONE 2014, 9, e90346. [Google Scholar]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows-Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. The variant call format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- RStudio Team. RStudio: Integrated Development for R; RStudio, PBC: Boston, MA, USA, 2020. [Google Scholar]
- Serapiglia, M.J.; Gouker, F.E.; Hart, J.F.; Unda, F.; Mansfield, S.D.; Stipanovic, A.J.; Smart, L.B. Ploidy level affects important biomass traits of novel shrub willow (Salix) hybrids. Bioenergy Res. 2015, 8, 259–269. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S; Springer: New York, NY, USA, 2002. [Google Scholar]
- Broman, K.W.; Wu, H.; Sen, S.; Churchill, G.A. R/qtl: QTL mapping in experimental crosses. Bioinformatics 2003, 19, 889–890. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rust ID | Year | Collection Date | Field | Host Pedigree | Clone/Cultivar |
|---|---|---|---|---|---|
| R15-001-01 | 2015 | 10 July 2015 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-051 |
| R15-002-01 | 2015 | 10 July 2015 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-051 |
| R15-003-01 | 2015 | 20 July 2015 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-051 |
| R15-004-01 | 2015 | 20 July 2015 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-051 |
| R15-005-01 | 2015 | 30 July 2015 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-051 |
| R15-006-01 | 2015 | 30 July 2015 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-051 |
| R15-007-01 | 2015 | 31 July 2015 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-051 |
| R15-008-01 | 2015 | 1 August 2015 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-051 |
| R15-009-01 | 2015 | 12 August 2015 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-051 |
| R15-010-01 | 2015 | 13 August 2015 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-051 |
| R16-003-03 | 2016 | 1 June 2016 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-048 |
| R16-004-01 | 2016 | 1 June 2016 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-051 |
| R16-005-01 | 2016 | 1 June 2016 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-053 |
| R16-006-01 | 2016 | 1 June 2016 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-055 |
| R17-001-02 | 2017 | 25 July 2017 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-051 |
| R17-002-01 | 2017 | 25 July 2017 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-051 |
| R17-002-02 | 2017 | 25 July 2017 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-051 |
| R17-002-03 | 2017 | 25 July 2017 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-051 |
| R17-003-01 | 2017 | 25 July 2017 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-054 |
| R17-004-01 | 2017 | 25 July 2017 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-076 |
| R17-004-03 | 2017 | 25 July 2017 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-076 |
| R17-005-01 | 2017 | 25 July 2017 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-063 |
| R17-007-03 | 2017 | 25 July 2017 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-059 |
| R17-008-02 | 2017 | 25 July 2017 | Salix F1 HCP * | S. purpurea × S. koriyanagi | 13X-438-045 |
| R18-002-01 | 2018 | 14 June 2018 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-094 |
| R18-003-02 | 2018 | 14 June 2018 | Family Selection Trial | S. miyabeana | 01-200-007 |
| R18-003-03 | 2018 | 14 June 2018 | Family Selection Trial | S. miyabeana | 01-200-007 |
| R18-004-02 | 2018 | 21 June 2018 | Salix F1 HCP | S. purpurea × S. udensis | 13X-358-003 |
| R18-004-03 | 2018 | 21 June 2018 | Salix F1 HCP | S. purpurea × S. udensis | 13X-358-003 |
| R18-005-03 | 2018 | 21 June 2018 | Salix F1 HCP | S. purpurea × S. udensis | 13X-358-001 |
| R18-006-01 | 2018 | 21 June 2018 | Salix F1 HCP | S. purpurea × S. udensis | 13X-358-157 |
| R18-006-03 | 2018 | 21 June 2018 | Salix F1 HCP | S. purpurea × S. udensis | 13X-358-157 |
| R18-007-02 | 2018 | 21 June 2018 | Salix F1 HCP | S. purpurea × S. udensis | 13X-358-124 |
| R18-007-03 | 2018 | 21 June 2018 | Salix F1 HCP | S. purpurea × S. udensis | 13X-358-124 |
| R18-008-01 | 2018 | 21 June 2018 | Salix F1 HCP | S. purpurea × S. udensis | 13X-358-177 |
| R18-009-02 | 2018 | 21 June 2018 | Salix F1 HCP | S. purpurea × S. koriyanagi | 13X-438-027 |
| R18-010-01 | 2018 | 21 June 2018 | Salix F1 HCP | S. purpurea × S. udensis | 13X-358-003 |
| R18-010-02 | 2018 | 21 June 2018 | Salix F1 HCP | S. purpurea × S. udensis | 13X-358-003 |
| R18-010-03 | 2018 | 21 June 2018 | Salix F1 HCP | S. purpurea × S. udensis | 13X-358-003 |
| R18-013-01 | 2018 | 25 June 2018 | Salix F1 HCP | S. purpurea × S. suchowensis | 13X-443-020 |
| R18-013-03 | 2018 | 25 June 2018 | Salix F1 HCP | S. purpurea × S. suchowensis | 13X-443-020 |
| R18-014-01 | 2018 | 25 June 2018 | Salix F1 HCP | S. purpurea × S. udensis | 13X-358-171 |
| R18-015-01 | 2018 | 25 June 2018 | Salix F1 HCP | S. purpurea × S. suchowensis | 13X-440-148 |
| R18-017-01 | 2018 | 25 June 2018 | Salix F1 HCP | S. purpurea × S. udensis | 13X-358-192 |
| R18-018-02 | 2018 | 25 June 2018 | Salix F1 HCP | S. purpurea × S. udensis | 13X-358-118 |
| R18-019-01 | 2018 | 25 June 2018 | Salix F1 HCP | S. purpurea × S. suchowensis | 13X-440-144 |
| R18-019-02 | 2018 | 25 June 2018 | Salix F1 HCP | S. purpurea × S. suchowensis | 13X-440-144 |
| R18-020-01 | 2018 | 25 June 2018 | Salix F1 HCP | S. purpurea × S. koriyanagi | 13X-438-094 |
| R18-022-03 | 2018 | 28 June 2018 | Salix F1 HCP | S. purpurea × S. udensis | 13X-358-013 |
| R18-023-03 | 2018 | 28 June 2018 | Salix F1 HCP | S. purpurea × S. suchowensis | 13X-443-031 |
| R18-024-01 | 2018 | 28 June 2018 | Salix F1 HCP | S. purpurea × S. udensis | 13X-358-135 |
| R18-024-02 | 2018 | 28 June 2018 | Salix F1 HCP | S. purpurea × S. udensis | 13X-358-135 |
| R18-027-03 | 2018 | 28 June 2018 | Salix F1 HCP | S. purpurea × S. udensis | 13X-358-085 |
| R20-002 | 2020 | 8 June 2020 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-043 |
| R20-003 | 2020 | 8 June 2020 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-053 |
| R20-027 | 2020 | 8 June 2020 | Family Selection Trial | S. purpurea × S. sociogenesis | 10X-400-051 |
| R20-027(S) | 2020 | 8 June 2020 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-051 |
| R20-047 | 2020 | 13 June 2020 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-057 |
| Overwinter-Canker1 | 2020 | 10 February 2021 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-009 |
| Overwinter-Canker2 | 2020 | 10 February 2021 | Family Selection Trial | S. purpurea × S. suchowensis | 10X-400-009 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crowell, C.R.; Wilkerson, D.G.; Smart, L.B.; Smart, C.D. Evidence of Asexual Overwintering of Melampsora paradoxa and Mapping of Stem Rust Host Resistance in Salix. Plants 2022, 11, 2385. https://doi.org/10.3390/plants11182385
Crowell CR, Wilkerson DG, Smart LB, Smart CD. Evidence of Asexual Overwintering of Melampsora paradoxa and Mapping of Stem Rust Host Resistance in Salix. Plants. 2022; 11(18):2385. https://doi.org/10.3390/plants11182385
Chicago/Turabian StyleCrowell, Chase R., Dustin G. Wilkerson, Lawrence B. Smart, and Christine D. Smart. 2022. "Evidence of Asexual Overwintering of Melampsora paradoxa and Mapping of Stem Rust Host Resistance in Salix" Plants 11, no. 18: 2385. https://doi.org/10.3390/plants11182385
APA StyleCrowell, C. R., Wilkerson, D. G., Smart, L. B., & Smart, C. D. (2022). Evidence of Asexual Overwintering of Melampsora paradoxa and Mapping of Stem Rust Host Resistance in Salix. Plants, 11(18), 2385. https://doi.org/10.3390/plants11182385