
Foliar Application of Rhodopseudomonas palustris Enhances the Rice Crop Growth and Yield under Field Conditions



Abstract
:1. Introduction
2. Results
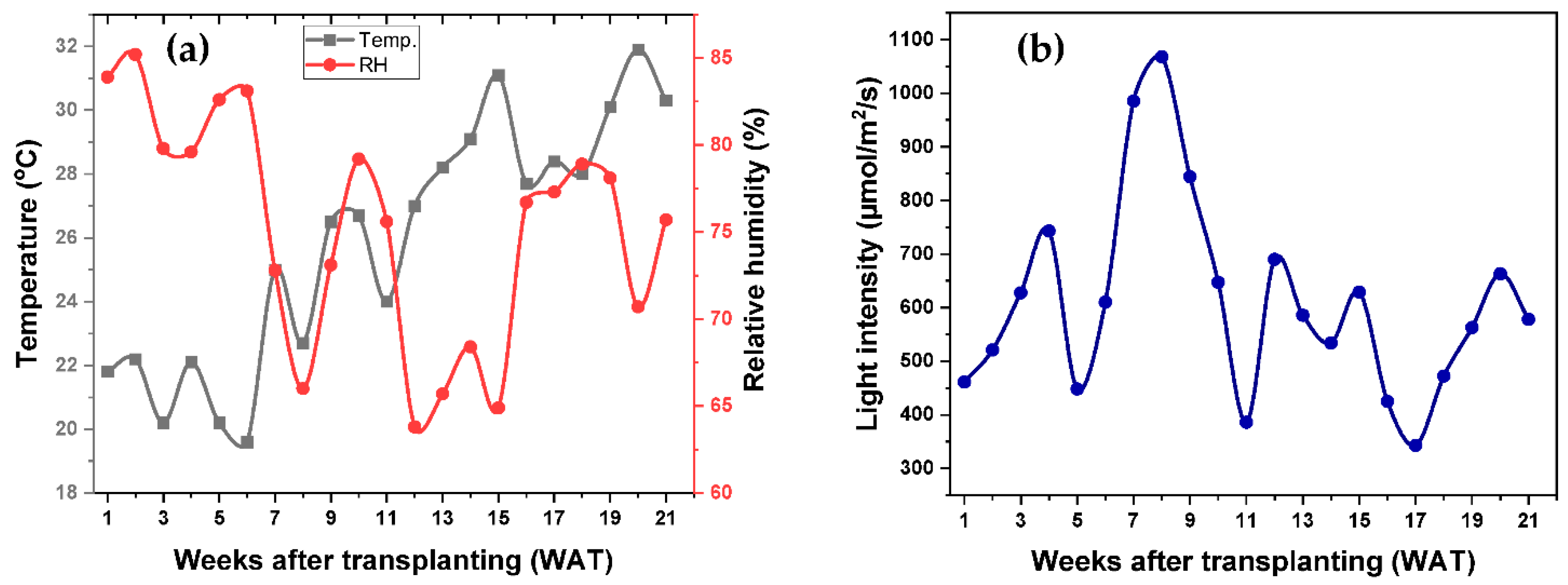
2.1. Agrometeorological Data
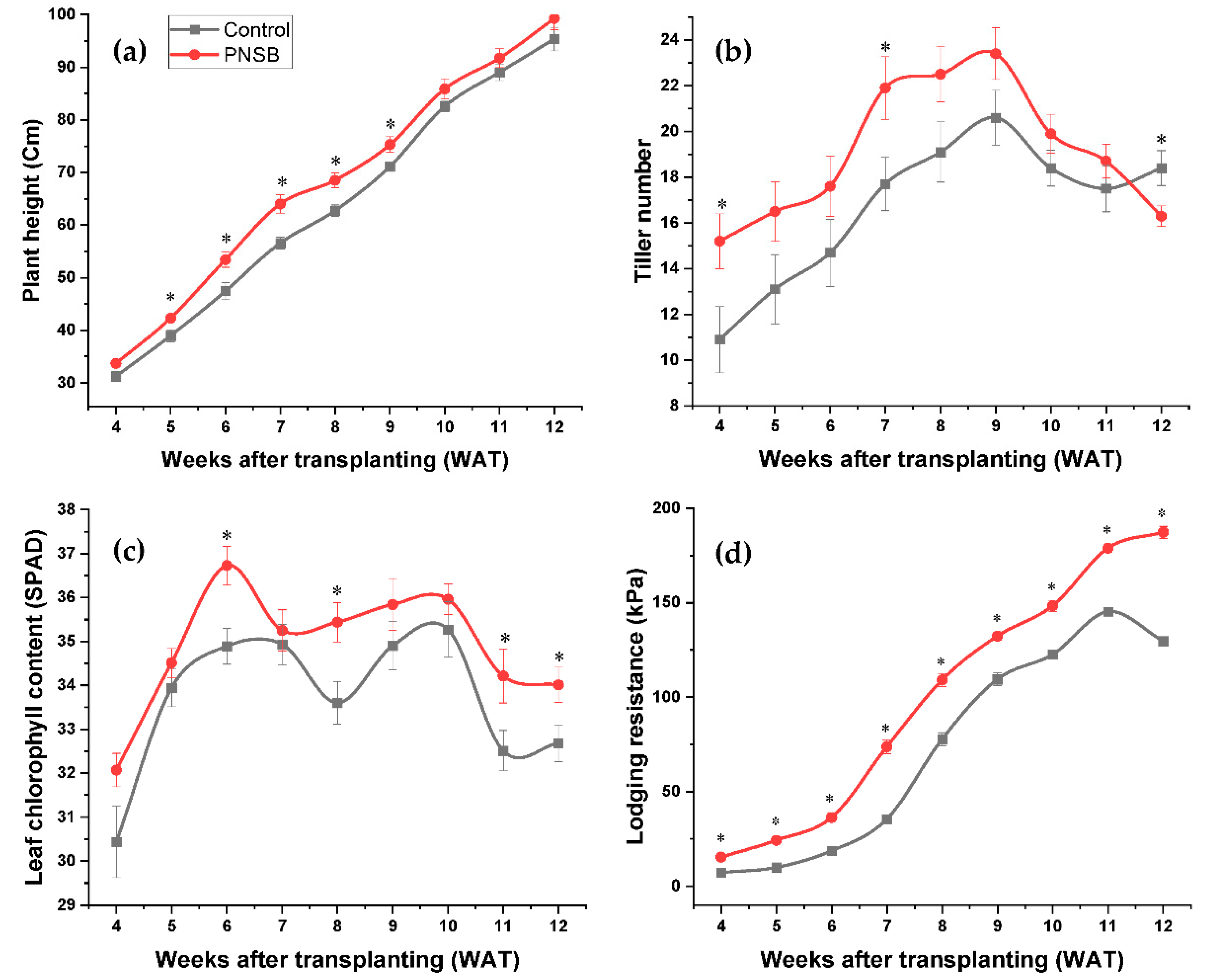
2.2. Agronomic Performances
2.3. Rice Root Growth and Biomass
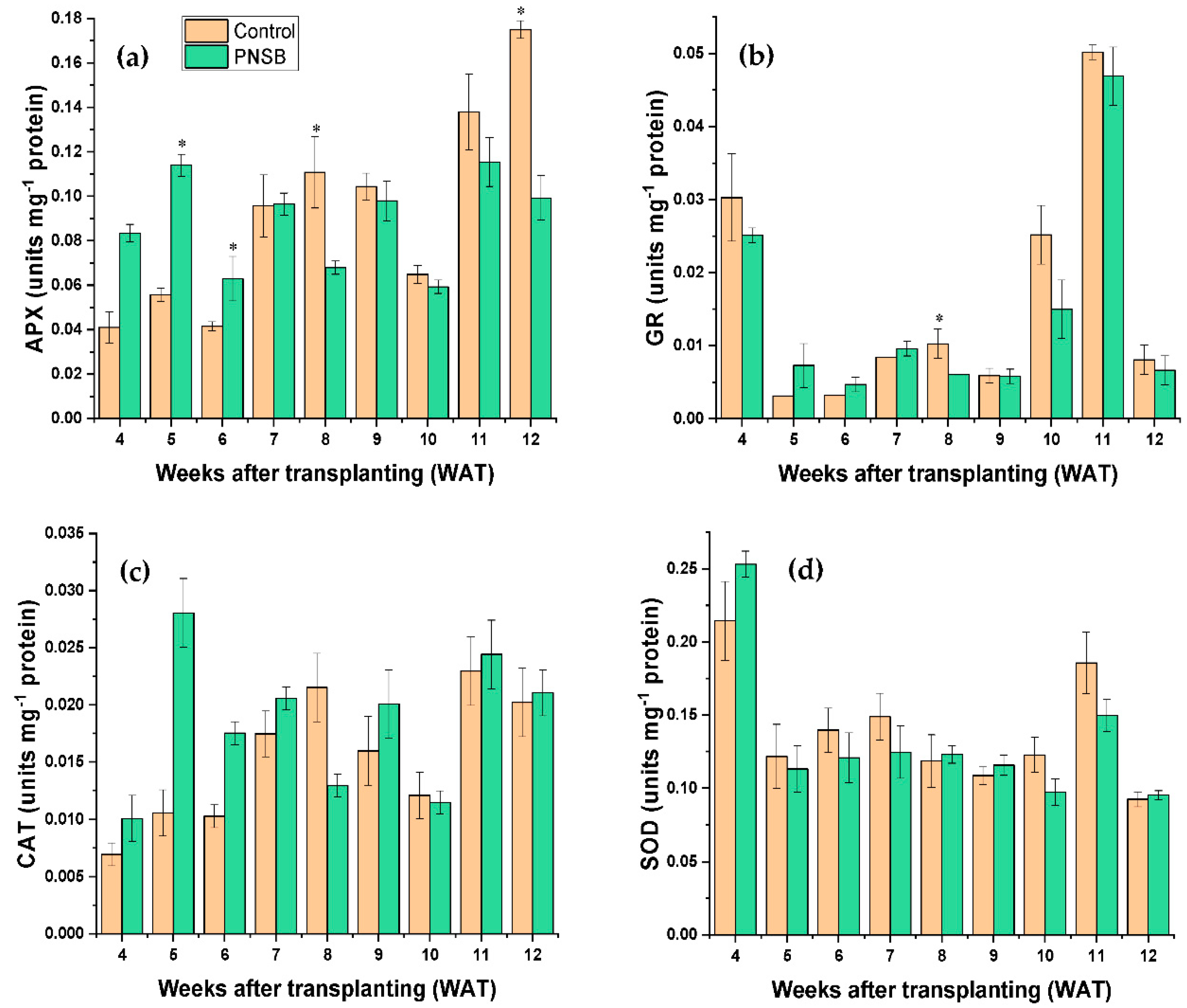
2.4. Antioxidant Enzyme Activity
2.5. Yield and Yield Components
3. Discussion
4. Materials and Methods
4.1. Experimental Location and Setup
4.2. Bacteria Preparation and Application
4.3. Crop Management Practices
4.4. Field Data Collection
4.5. Rice Root Growth and Biomass Analysis
4.6. Antioxidant Enzyme Activity Analysis
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jagadish, S.V.K.; Pal, M. Response of Rice (Oryza sativa L.) to Increasing Temperature and Atmospheric CO2. In Climate Change and Crops; Singh, S.N., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 63–80. ISBN 978-3-540-88246-6. [Google Scholar]
- Young, A. Land Resources: Now and for the Future; University of Cambridge: Cambridge, UK, 1998; ISBN 0-521-59003-5. [Google Scholar]
- Khush, G.S. What it will take to Feed 5.0 Billion Rice consumers in 2030. Plant Mol. Biol. 2005, 59, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Stefanos, A.N.; Anastasios, M.; Fotios, C. Climate change and agricultural productivity. Afr. J. Agric. Res. 2012, 7, 4885–4893. [Google Scholar] [CrossRef]
- Nelson, G.C.; Rosegrant, M.W.; Koo, J.; Robertson, R.; Sulser, T.; Zhu, T.; Ringler, C.; Msangi, S.; Palazzo, A.; Batka, M. Climate Change: Impact on Agriculture and Costs of Adaptation; International Food Policy Research Institute: Washington, DC, USA, 2009; Volume 21, ISBN 0-89629-535-4. [Google Scholar]
- Chen, Y.-H.; Prinn, R.G. Estimation of atmospheric methane emissions between 1996 and 2001 using a three-dimensional global chemical transport model. J. Geophys. Res. Atmos. 2006, 111, 1–25. [Google Scholar] [CrossRef]
- CGIAR The Global Staple. Available online: https://ricepedia.org/rice-as-food/the-global-staple-rice-consumers (accessed on 5 December 2021).
- Fukagawa, N.K.; Ziska, L.H. Rice: Importance for Global Nutrition. J. Nutr. Sci. Vitaminol. 2019, 65, S2–S3. [Google Scholar] [CrossRef]
- Runkle, B.R.K.; Seyfferth, A.L.; Reid, M.C.; Limmer, M.A.; Moreno-García, B.; Reavis, C.W.; Peña, J.; Reba, M.L.; Adviento-Borbe, M.A.A.; Pinson, S.R.M.; et al. Socio-Technical Changes for Sustainable Rice Production: Rice Husk Amendment, Conservation Irrigation, and System Changes. Front. Agron. 2021, 3, 81. [Google Scholar] [CrossRef]
- Seck, P.A.; Diagne, A.; Mohanty, S.; Wopereis, M.C.S. Crops that feed the world 7: Rice. Food Sec. 2012, 4, 7–24. [Google Scholar] [CrossRef]
- Rashid, M.H.; Kamruzzaman, M.; Haque, A.N.A.; Krehenbrink, M. Soil Microbes for Sustainable Agriculture. In Sustainable Management of Soil and Environment; Meena, R.S., Kumar, S., Bohra, J.S., Jat, M.L., Eds.; Springer: Singapore, 2019; pp. 339–382. ISBN 9789811388323. [Google Scholar]
- Sakarika, M.; Spanoghe, J.; Sui, Y.; Wambacq, E.; Grunert, O.; Haesaert, G.; Spiller, M.; Vlaeminck, S.E. Purple non-sulphur bacteria and plant production: Benefits for fertilization, stress resistance and the environment. Microb. Biotechnol. 2020, 13, 1336–1365. [Google Scholar] [CrossRef]
- Larimer, F.W.; Chain, P.; Hauser, L.; Lamerdin, J.; Malfatti, S.; Do, L.; Land, M.L.; Pelletier, D.A.; Beatty, J.T.; Lang, A.S. Complete genome sequence of the metabolically versatile photosynthetic bacterium Rhodopseudomonas palustris. Nat. Biotechnol. 2004, 22, 55–61. [Google Scholar] [CrossRef]
- Wong, W.-T.; Tseng, C.-H.; Hsu, S.-H.; Lur, H.-S.; Mo, C.-W.; Huang, C.-N.; Hsu, S.-C.; Lee, K.-T.; Liu, C.-T. Promoting Effects of a Single Rhodopseudomonas palustris Inoculant on Plant Growth by Brassica rapa chinensis under Low Fertilizer Input. Microbes Environ. 2014, 29, 303–313. [Google Scholar] [CrossRef]
- Xu, J.; Feng, Y.; Wang, Y.; Luo, X.; Tang, J.; Lin, X. The foliar spray of Rhodopseudomonas palustris grown under Stevia residue extract promotes plant growth via changing soil microbial community. J. Soils Sediments 2016, 16, 916–923. [Google Scholar] [CrossRef]
- ShuHua, H.; KaiJiun, L.; Wei, F.; HuuSheng, L.; ChiTe, L. Application of phototrophic bacterial inoculant to reduce nitrate content in hydroponic leafy vegetables. Crop Environ. Amp. Bioinform. 2015, 12, 30–41. [Google Scholar]
- Xu, J.; Feng, Y.; Wang, Y.; Lin, X. Effect of Rhizobacterium Rhodopseudomonas palustris Inoculation on Stevia rebaudiana Plant Growth and Soil Microbial Community. Pedosphere 2018, 28, 793–803. [Google Scholar] [CrossRef]
- Su, P.; Tan, X.; Li, C.; Zhang, D.; Cheng, J.; Zhang, S.; Zhou, X.; Yan, Q.; Peng, J.; Zhang, Z.; et al. Photosynthetic bacterium Rhodopseudomonas palustris GJ-22 induces systemic resistance against viruses. Microb. Biotechnol. 2017, 10, 612–624. [Google Scholar] [CrossRef]
- JianFeng, H.; YouZhi, F.; JianFeng, B.; YunLong, Y.; XianGui, L. Co-inoculation with am fungus Glomus caledonium and the photoheterotrophic purple nonsulfur bacterium Rhodopseudomonas palustris results in mutual inhibition and lower arsenic accumulation of Nicotiana tabacum L. in an arsenic contaminated soil. Fresenius Environ. Bull. 2014, 23, 867–874. [Google Scholar]
- Han, J. The influence of photosynthetic bacteria treatments on the crop yield, dry matter content, and protein content of the mushroom Agaricus bisporus. Sci. Hortic. 1999, 82, 171–178. [Google Scholar] [CrossRef]
- Yin, Z.P.; Shang, Z.W.; Wei, C.; Ren, J.; Song, X.S. Foliar sprays of photosynthetic bacteria improve the growth and anti-oxidative capability on Chinese dwarf cherry seedlings. J. Plant Nutr. 2012, 35, 840–853. [Google Scholar] [CrossRef]
- Batool, K.; Rehman, Y. Arsenic-redox transformation and plant growth promotion by purple nonsulfur bacteria Rhodopseudomonas palustris CS2 and Rhodopseudomonas faecalis SS5. BioMed Res. Int. 2017, 2017, 6250327. [Google Scholar] [CrossRef]
- Harada, N.; Nishiyama, M.; Otsuka, S.; Matsumoto, S. Effects of inoculation of phototrophic purple bacteria on grain yield of rice and nitrogenase activity of paddy soil in a pot experiment. Soil Sci. Plant Nutr. 2005, 51, 361–367. [Google Scholar] [CrossRef]
- Kantha, T.; Kantachote, D.; Klongdee, N. Potential of biofertilizers from selected Rhodopseudomonas palustris strains to assist rice (Oryza sativa L. subsp. indica) growth under salt stress and to reduce greenhouse gas emissions. Ann. Microbiol. 2015, 65, 2109–2118. [Google Scholar] [CrossRef]
- Kantachote, D.; Nunkaew, T.; Kantha, T.; Chaiprapat, S. Biofertilizers from Rhodopseudomonas palustris strains to enhance rice yields and reduce methane emissions. Appl. Soil Ecol. 2016, 100, 154–161. [Google Scholar] [CrossRef]
- Nookongbut, P.; Kantachote, D.; Megharaj, M.; Naidu, R. Reduction in arsenic toxicity and uptake in rice (Oryza sativa L.) by As-resistant purple nonsulfur bacteria. Environ. Sci. Pollut. Res. 2018, 25, 36530–36544. [Google Scholar] [CrossRef] [PubMed]
- de los Reyes, B.G.; Myers, S.J.; McGrath, J.M. Differential induction of glyoxylate cycle enzymes by stress as a marker for seedling vigor in sugar beet (Beta vulgaris). Mol. Gen. Genom. 2003, 269, 692–698. [Google Scholar] [CrossRef] [PubMed]
- Downton, J.; Slatyer, R.O. Temperature Dependence of Photosynthesis in Cotton. Plant Physiol. 1972, 50, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Horie, T. Crop Ontogeny and Development. In Physiology and Determination of Crop Yield; John Wiley & Sons, Ltd.: New York, NY, USA, 1994; pp. 153–180. ISBN 978-0-89118-246-7. [Google Scholar]
- Yoshida, S.; Satake, T.; Mackill, D.S. High-temperature stress in rice [study conducted at IRRI, Philippines]. IRRI Res. Pap. Ser. 1981, 67, 15. [Google Scholar]
- Porter, J.R.; Semenov, M.A. Crop responses to climatic variation. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 2021–2035. [Google Scholar] [CrossRef]
- Harrington, S. Greenhouse Procedures Manual. Available online: http://ricelab.plbr.cornell.edu/greenhouse_procedures_manual#lighting (accessed on 3 August 2022).
- Liu, Q.; Wu, X.; Chen, B.; Ma, J.; Gao, J. Effects of Low Light on Agronomic and Physiological Characteristics of Rice Including Grain Yield and Quality. Rice Sci. 2014, 21, 243–251. [Google Scholar] [CrossRef]
- Taiwan News. Abnormal Weather Dampens Millet Harvest in East Taiwan. Available online: https://www.taiwannews.com.tw/en/news/4567308 (accessed on 5 August 2022).
- Hsing, Y.I.C. Rice in Taiwan. In Encyclopaedia of the History of Science, Technology, and Medicine in Non-Western Cultures; Selin, H., Ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 1–3. ISBN 978-94-007-3934-5. [Google Scholar]
- Council of Agriculture, Executive Yuan, R.O.C. Kaohsiung No. 147. Available online: https://kmweb.coa.gov.tw (accessed on 6 August 2022).
- Yoshida, T.; Tabata, T.; Saraswati, R.; Kobayashi, M. Study on resourceful disposal of organic waste and high-yielding culture of rice plant. J. Environ. Conserv. Eng. 1991, 20, 607–610. [Google Scholar] [CrossRef]
- Elbadry, M.; Gamal-Eldin, H.; Elbanna, K. Effects of Rhodobacter capsulatus inoculation in combination with graded levels of nitrogen fertilizer on growth and yield of rice in pots and lysimeter experiments. World J. Microbiol. Biotechnol. 1999, 15, 393–395. [Google Scholar] [CrossRef]
- Kato, M.; Shimizu, S. Chlorophyll metabolism in higher plants. VII. Chlorophyll degradation in senescing tobacco leaves; phenolic-dependent peroxidative degradation. Can. J. Bot. 1987, 65, 729–735. [Google Scholar] [CrossRef]
- Elbadry, M.; Elbanna, K. Response of four rice varieties to Rhodobacter capsulatus at seedling stage. World J. Microbiol. Biotechnol. 1999, 15, 363–367. [Google Scholar] [CrossRef]
- Gamal-Eldin, H.; Elbanna, K. Field evidence for the potential of Rhodobacter capsulatus as biofertilizer for flooded rice. Curr. Microbiol. 2011, 62, 391–395. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Lu, K.; Nie, H.; Zeng, Q.; Wu, B.; Qian, J.; Fang, Z. Rice nitrate transporter OsNPF7.2 positively regulates tiller number and grain yield. Rice 2018, 11, 12. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Wang, D.; Yan, S.; Liu, S.; Liu, B.; Kang, H.; Wang, G.-L. Dissection of the Genetic Architecture of Rice Tillering using a Genome-wide Association Study. Rice 2019, 12, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.; Jang, S.; Lee, Y.K.; Kim, D.-G.; Jin, Z.; Koh, H.-J. Genetic Basis of Tiller Dynamics of Rice Revealed by Genome-Wide Association Studies. Plants 2020, 9, 1695. [Google Scholar] [CrossRef]
- Pawar, S.; Radhakrishnan, V.; Mohanan, K.V. The Importance of Optimum Tillering in Rice—An overview. South Indian J. Biol. Sci. 2016, 2, 125. [Google Scholar] [CrossRef]
- Moldenhauer, K.; Slaton, N. Rice growth and development. Rice Prod. Handb. 2001, 192, 7–14. [Google Scholar]
- Nunkaew, T.; Kantachote, D.; Kanzaki, H.; Nitoda, T.; Ritchie, R.J. Effects of 5-aminolevulinic acid (ALA)-containing supernatants from selected Rhodopseudomonas palustris strains on rice growth under NaCl stress, with mediating effects on chlorophyll, photosynthetic electron transport and antioxidative enzymes. Electron. J. Biotechnol. 2014, 17, 4. [Google Scholar] [CrossRef]
- Maudinas, B.; Chemardin, M.; Yovanovitch, E.; Gadal, P. Gnotobiotic cultures of rice plants up to ear stage in the absence of combined nitrogen source but in the presence of free living nitrogen fixing bacteria Azotobacter vinelandii and Rhodopseudomonas capsulata. Plant Soil 1981, 60, 85–97. [Google Scholar] [CrossRef]
- Kreiner, M.; Harvey, L.M.; McNeil, B. Oxidative stress response of a recombinant Aspergillus niger to exogenous menadione and H2O2 addition. Enzym. Microb. Technol. 2002, 30, 346–353. [Google Scholar] [CrossRef]
- McDonald, A.E.; Vanlerberghe, G.C. Alternative oxidase and plastoquinol terminal oxidase in marine prokaryotes of the Sargasso Sea. Gene 2005, 349, 15–24. [Google Scholar] [CrossRef]
- Liu, L.; Wang, L.; Deng, F.; Huang, Y.; Liu, D.; Ren, W.; Yang, W. Osmotic regulation substance contents and activities of protective enzymes in leaves of different hybrid rice combinations as affected by shading. Chin. J. Rice Sci. 2012, 26, 569–575. [Google Scholar] [CrossRef]
- Lee, S.-K.; Lur, H.-S.; Lo, K.-J.; Cheng, K.-C.; Chuang, C.-C.; Tang, S.-J.; Yang, Z.-W.; Liu, C.-T. Evaluation of the effects of different liquid inoculant formulations on the survival and plant-growth-promoting efficiency of Rhodopseudomonas palustris strain PS3. Appl. Microbiol. Biotechnol. 2016, 100, 7977–7987. [Google Scholar] [CrossRef]
- Sundar, L.S.; Chen, G.S. Study on the Growth Performance of Lettuce (Lactuca sativa) and Pak Choi (Brassica chinensis) in Different Aquaponic Growing Systems. Horticulturae 2020, 6, 69. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Foster, J.G.; Hess, J.L. Responses of superoxide dismutase and glutathione reductase activities in cotton leaf tissue exposed to an atmosphere enriched in oxygen. Plant Physiol. 1980, 66, 482–487. [Google Scholar] [CrossRef]
- Paoletti, F.; Aldinucci, D.; Mocali, A.; Caparrini, A. A sensitive spectrophotometric method for the determination of superoxide dismutase activity in tissue extracts. Anal. Biochem. 1986, 154, 536–541. [Google Scholar] [CrossRef]
- Lee, S.-K.; Lur, H.-S.; Liu, C.-T. From Lab to Farm: Elucidating the Beneficial Roles of Photosynthetic Bacteria in Sustainable Agriculture. Microorganisms 2021, 9, 2453. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Root Length (cm) | Root Volume (cm3) | Root Dry Weight (g) |
|---|---|---|---|
| Control | 43.5 ± 0.21 b | 1200 ± 0.00 a | 18.1 ± 2.20 b |
| PNSB | 57.8 ± 2.65 a | 1333 ± 66.7 a | 41.6 ± 4.01 a |
| Treatment | Productive Tillers/Plant (%) | Average Grains/Plant | Average Grains/Panicle | Biological Yield (t ha−1) |
|---|---|---|---|---|
| Control | 48.00 ± 2.40 b | 1613.70 ± 55.75 b | 80.50 ± 3.51 a | 17.46 ± 1.42 a |
| PNSB | 64.44 ± 2.72 a | 2584.00 ± 172.32 a | 82.17 ± 3.22 a | 14.37 ± 0.90 a |
| Treatment | Grain Yield (t ha−1) | Grain Fertility (%) | 1000 Grain Weight (g) | Harvest Index (%) |
|---|---|---|---|---|
| Control | 5.42 ± 0.35 b | 42.02 ± 3.68 a | 23.98 ± 0.07 b | 0.33 ± 0.03 b |
| PNSB | 8.10 ± 0.73 a | 46.99 ± 1.81 a | 24.37 ± 0.11 a | 0.56 ± 0.04 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yen, K.S.; Sundar, L.S.; Chao, Y.-Y. Foliar Application of Rhodopseudomonas palustris Enhances the Rice Crop Growth and Yield under Field Conditions. Plants 2022, 11, 2452. https://doi.org/10.3390/plants11192452
Yen KS, Sundar LS, Chao Y-Y. Foliar Application of Rhodopseudomonas palustris Enhances the Rice Crop Growth and Yield under Field Conditions. Plants. 2022; 11(19):2452. https://doi.org/10.3390/plants11192452
Chicago/Turabian StyleYen, Kuei Shan, Laurence Shiva Sundar, and Yun-Yang Chao. 2022. "Foliar Application of Rhodopseudomonas palustris Enhances the Rice Crop Growth and Yield under Field Conditions" Plants 11, no. 19: 2452. https://doi.org/10.3390/plants11192452
APA StyleYen, K. S., Sundar, L. S., & Chao, Y. -Y. (2022). Foliar Application of Rhodopseudomonas palustris Enhances the Rice Crop Growth and Yield under Field Conditions. Plants, 11(19), 2452. https://doi.org/10.3390/plants11192452