
Effects of UV-B Radiation on the Performance, Antioxidant Response and Protective Compounds of Hazelnut Pollen
 , , , and
, , , and
Abstract
:1. Introduction
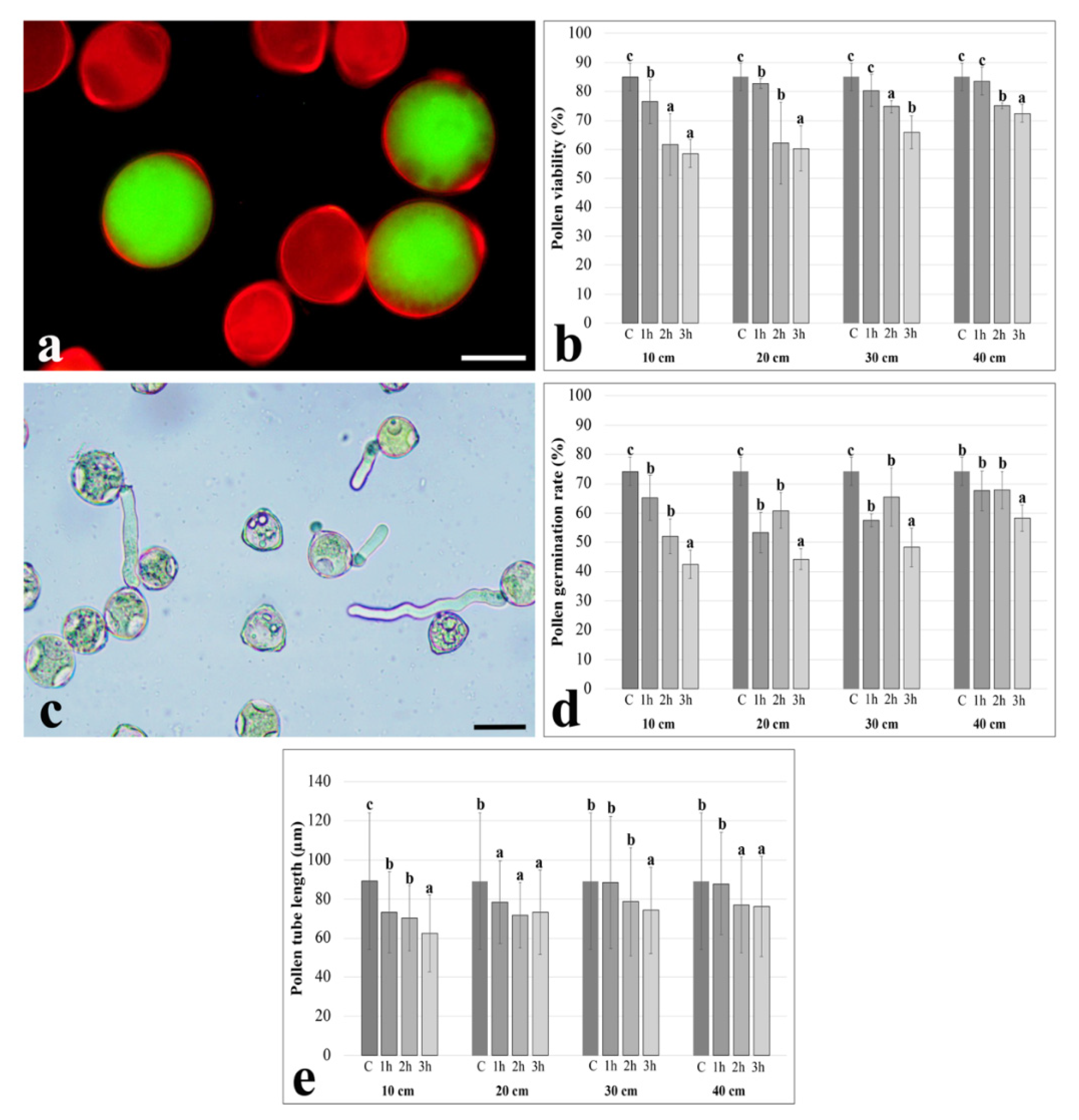
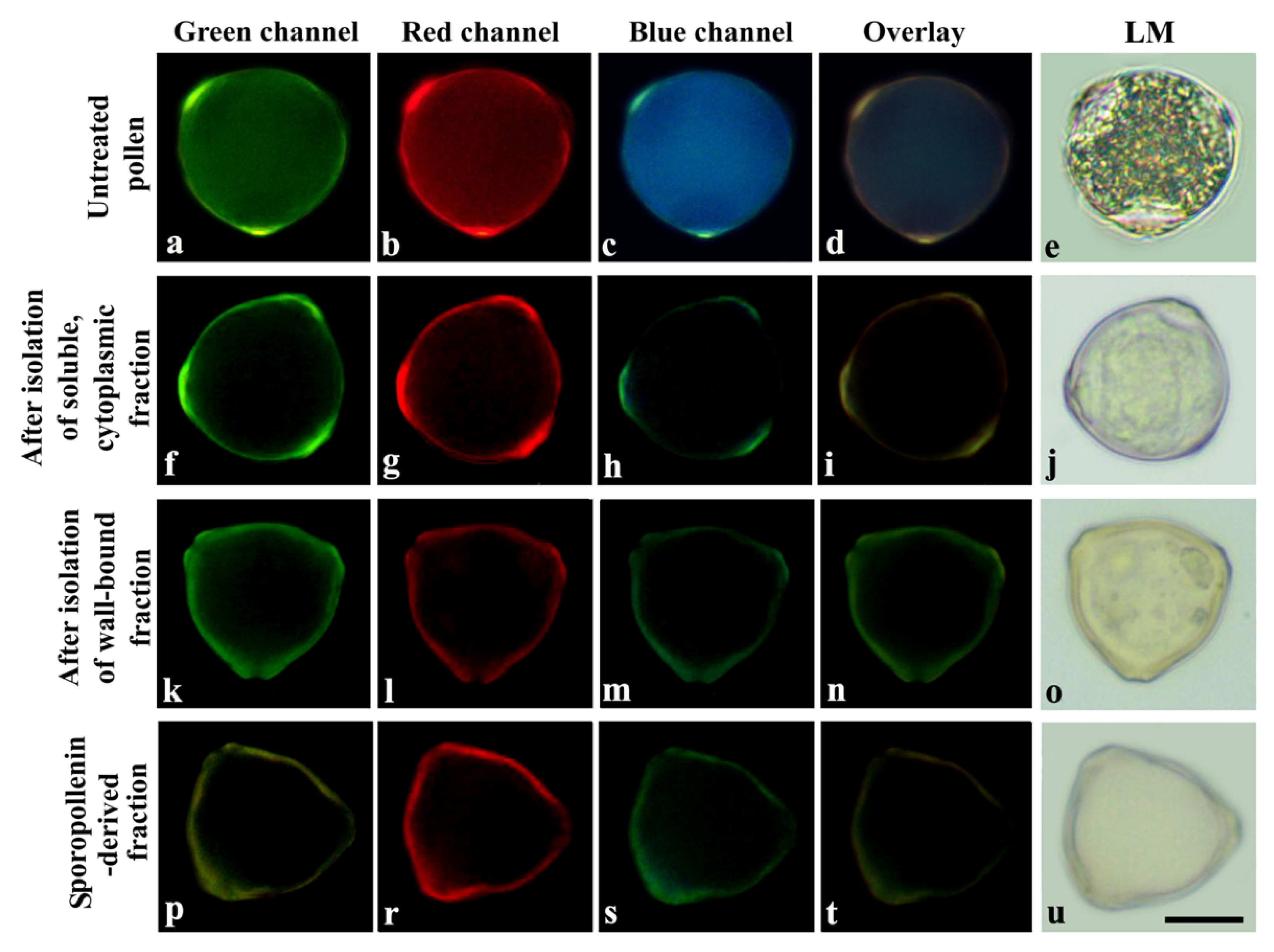
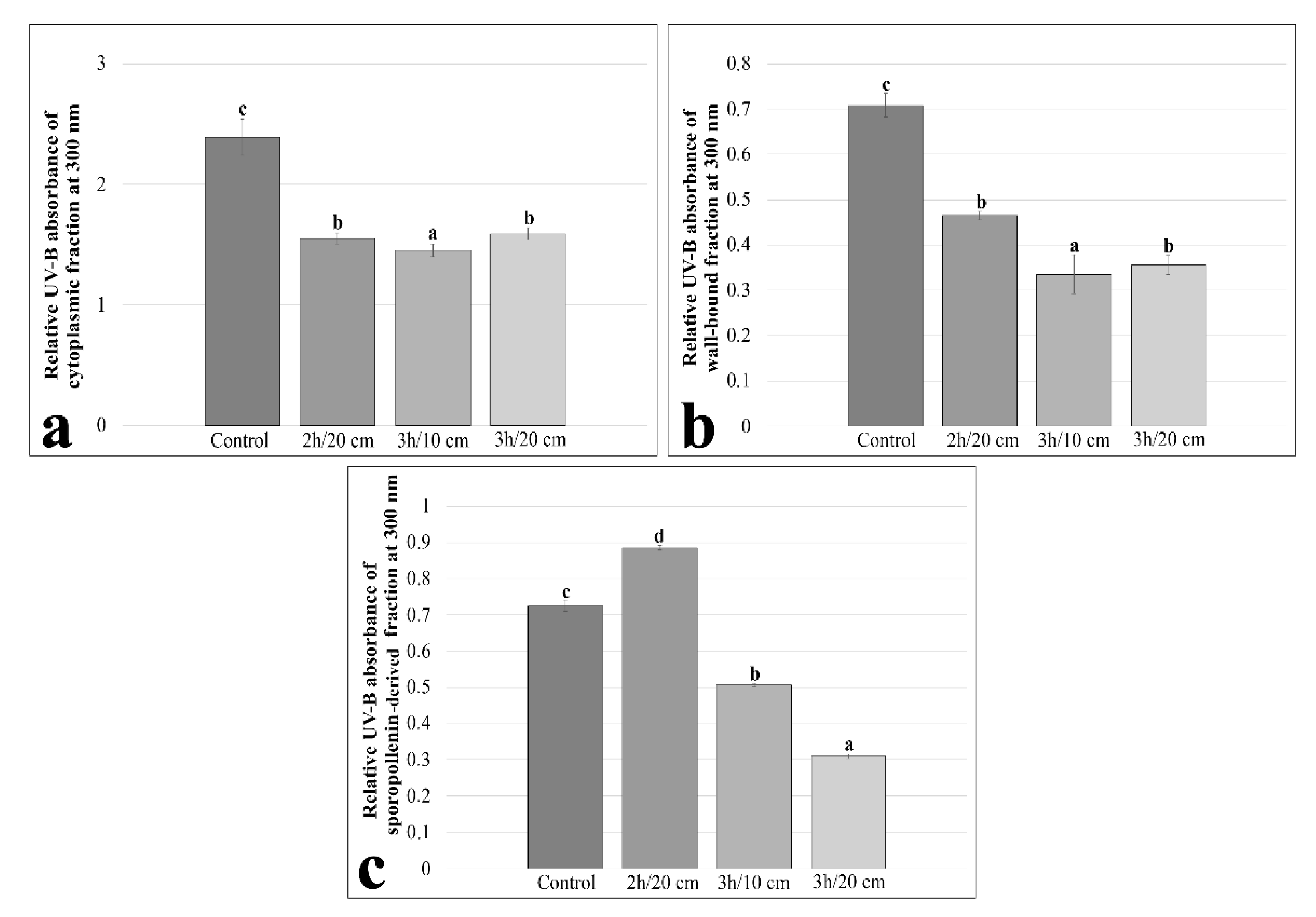
2. Results
3. Discussion
4. Materials and Methods
4.1. Plant Material and Application of UV-B Treatment
4.2. Pollen Viability
4.3. In Vitro Pollen Germination and Tube Length
4.4. Determination of Cumulative Stress Response Index
4.5. Determination of Total Antioxidant Activity, Total Phenolic and Flavonoid Contents
4.6. Sequential Extraction of UACs in Pollen Grains and Fluorescence Microscope Analysis
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Masullo, M.; Lauro, G.; Cerulli, A.; Bifulco, G.; Piacente, S. Corylus avellana: A Source of diarylheptanoids with α-glucosidase inhibitory activity evaluated by in vitro and in silico studies. Front. Plant Sci. 2022, 13, 805660. [Google Scholar] [CrossRef]
- Puliga, F.; Leonardi, P.; Minutella, F.; Zambonelli, A.; Francioso, O. Valorization of hazelnut shells as growing substrate for edible and medicinal mushrooms. Sci. Hortic. 2022, 8, 214. [Google Scholar] [CrossRef]
- Romero-Aroca, A.; Rovira, M.; Cristofori, V.; Silvestri, C. Hazelnut kernel size and industrial aptitude. J. Agric. 2021, 11, 1115. [Google Scholar] [CrossRef]
- Silvestri, C.; Santinelli, G.; Pica, A.L.; Cristofori, V. Mechanical pruning of European hazelnut: Effects on yield and quality and potential to exploit its by-product. Eur. J. Hortic. Sci. 2021, 86, 189–196. [Google Scholar] [CrossRef]
- İslam, A. Hazelnut culture in Turkey. Akademik Ziraat Dergisi 2018, 7, 259–266. [Google Scholar] [CrossRef]
- Chaudhry, S.; Sidhu, G.P.S. Climate change regulated abiotic stress mechanisms in plants: A comprehensive review. Plant Cell Rep. 2021, 41, 1–31. [Google Scholar] [CrossRef]
- Çetinbaş-Genç, A.; Cai, G.; Del Duca, S.; Vardar, F.; Ünal, M. The effect of putrescine on pollen performance in hazelnut (Corylus avellana L.). Sci. Hortic. 2020, 261, 108971. [Google Scholar] [CrossRef]
- Liu, M.; Zhao, R.; Chen, Y.; Zhang, J.; Compton, S.G.; Chen, X.Y. Competitive exclusion among fig wasps achieved via entrainment of host plant flowering phenology. PLoS ONE 2014, 9, e97783. [Google Scholar] [CrossRef] [PubMed]
- Hamidi, R.; Calvy, M.; Valentie, E.; Driss, L.; Guignet, J.; Thomas, M.; Tavella, L. Symptoms resulting from the feeding of true bugs on growing hazelnuts. Entomol. Exp. Appl. 2022, 170, 477–487. [Google Scholar] [CrossRef]
- Heslop-Harrison, J.; Heslop-Harrison, Y.; Cresti, M.; Tiezzi, A.; Ciampolini, F. Actin during pollen germination. J. Cell Sci. 1986, 86, 1–8. [Google Scholar] [CrossRef]
- Brito, C.; Dinis, L.T.; Moutinho-Pereira, J.; Correia, C.M. Drought stress effects and olive tree acclimation under a changing climate. Plants 2019, 8, 232. [Google Scholar] [CrossRef] [PubMed]
- Escobar-Bravo, R.; Nederpel, C.; Naranjo, S.; Kim, H.K.; Rodríguez-López, M.J.; Chen, G.; Glauser, G.; Leiss, K.A.; Klinkhamer, P.G. Ultraviolet radiation modulates both constitutive and inducible plant defenses against thrips but is dose and plant genotype dependent. J. Pest Sci. 2021, 94, 69–81. [Google Scholar] [CrossRef]
- Zedek, F.; Šmerda, J.; Veselý, P.; Horová, L.; Kocmanová, J.; Bureš, P. Elevation-dependent endopolyploid response suggests that plants with holocentric chromosomes are less stressed by UV-B. Bot. J. Linn. 2021, 195, 106–113. [Google Scholar] [CrossRef]
- Bornman, J.F.; Barnes, P.W.; Robson, T.M.; Robinson, S.A.; Jansen, M.A.; Ballare, C.L.; Flint, S.D. Linkages between stratospheric ozone, UV radiation and climate change and their implications for terrestrial ecosystems. Photochem. Photobiol. Sci. 2019, 18, 681–716. [Google Scholar] [CrossRef] [PubMed]
- Llorens, L.; Badenes-Pérez, F.R.; Julkunen-Tiitto, R.; Zidorn, C.; Fereres, A.; Jansen, M.A. The role of UV-B radiation in plant sexual reproduction. Perspect. Plant Ecol. Evol. Syst. 2015, 17, 243–254. [Google Scholar] [CrossRef]
- Yadav, A.; Singh, D.; Lingwan, M.; Yadukrishnan, P.; Masakapalli, S.K.; Datta, S. Light signaling and UV-B-mediated plant growth regulation. J. Integr. Plant Biol. 2020, 62, 1270–1292. [Google Scholar] [CrossRef]
- Piccini, C.; Cai, G.; Dias, M.C.; Araújo, M.; Parri, S.; Romi, M.; Faleri, C.; Cantini, C. Olive Varieties under UV-B stress show distinct responses in terms of antioxidant machinery and isoform/activity of RubisCO. Int. J. Mol. Sci. 2021, 22, 11214. [Google Scholar] [CrossRef] [PubMed]
- Demchik, S.M.; Day, T.A. Effect of enhanced UV-B radiation on pollen quantity, quality, and seed yield in Brassica rapa (Brassicaceae). Am. J. Bot. 1996, 83, 573–579. [Google Scholar] [CrossRef]
- Zhang, C.; Chen, M.; Liu, G.; Huang, G.; Wang, Y.; Yang, S.; Xu, X. Enhanced UV-B radiation aggravates negative effects more in females than in males of Morus alba saplings under drought stress. Environ. Exp. Bot. 2020, 169, 103903. [Google Scholar] [CrossRef]
- Feng, H.; An, L.; Tan, L.; Hou, Z.; Wang, X. Effect of enhanced ultraviolet-B radiation on pollen germination and tube growth of 19 taxa in vitro. Environ. Exp. Bot. 2000, 43, 45–53. [Google Scholar] [CrossRef]
- Wang, S.; Xie, B.; Yin, L.; Duan, L.; Li, Z.; Egrinya Eneji, A.; Tusuji, E.; Tsunekawa, A. Increased UV-B radiation affects the viability, reactive oxygen species accumulation and antioxidant enzyme activities in maize (Zea mays L.) pollen. Photochem. Photobiol. Sci. 2010, 86, 110–116. [Google Scholar] [CrossRef]
- Sailaja, K.; Reddy, K.R.; Reddy, V.R.; Kakani, V.G.; Zhao, D. Interactive effects of carbon dioxide, temperature, and UV-B radiation on Soybean (Glycine max L.) flower and pollen morphology, pollen production, germination, and tube length. J. Exp. Bot. 2005, 56, 725–736. [Google Scholar] [CrossRef]
- Koubouris, G.C.; Kavroulakis, N.; Metzidakis, I.T.; Vasilakakis, M.D.; Sofo, A. Ultraviolet-B radiation or heat cause changes in photosynthesis, antioxidant enzyme activities and pollen performance in olive tree. Photosynthetica 2015, 53, 279–287. [Google Scholar] [CrossRef]
- Del Valle, J.C.; Buide, M.L.; Whittall, J.B.; Valladares, F.; Narbona, E. UV radiation increases phenolic compound protection but decreases reproduction in Silene littorea. PLoS ONE 2020, 15, e0231611. [Google Scholar] [CrossRef] [PubMed]
- Çetinbaş-Genç, A.; Conti, V.; Cai, G. Let’s shape again: The concerted molecular action that builds the pollen tube. Plant Reprod. 2022, 35, 77–103. [Google Scholar] [CrossRef]
- Shivanna, K.R.; Linskens, H.F.; Cresti, M. Pollen viability and pollen vigor. Theor. Appl. Genet. 1991, 81, 38–42. [Google Scholar] [CrossRef]
- Parrotta, L.; Faleri, C.; Guerriero, G.; Cai, G. Cold stress affects cell wall deposition and growth pattern in tobacco pollen tubes. Plant Sci. J. 2019, 283, 329–342. [Google Scholar] [CrossRef]
- Navabpour, S.; Almas, D.E.; Kafi, H.; Aghdam, S.M. The role of reactive oxygen species in uv-b-inhibited pollen germination and tube growth of canola. Int. J. Biomed. Data Min. 2021, 10, 135. [Google Scholar]
- Fatima, K.; Sardar, F.; Imran, A. Regulation of NO biosynthesis under abiotic stresses and modulation due to osmolytes. In Nitric Oxide in Plants: A Molecule with Dual Roles, 1st ed.; Ahanger, M.A., Ahmad, P., Eds.; Wiley: Hoboken, NJ, USA, 2022; pp. 26–41. [Google Scholar] [CrossRef]
- Kumar, S.; Abedin, M.; Singh, A.K.; Das, S. Role of phenolic compounds in plant-defensive mechanisms. In Plant Phenolics in Sustainable Agriculture, 1st ed.; Lone, L., Shuab, R., Kamili, A.R., Eds.; Springer: Singapore, 2020; pp. 517–532. [Google Scholar] [CrossRef]
- Piccini, C.; Cai, G.; Dias, M.C.; Romi, M.; Longo, R.; Cantini, C. UV-B radiation affects photosynthesis-related processes of two Italian Olea europaea (L.) varieties differently. Plants 2020, 9, 1712. [Google Scholar] [CrossRef]
- Jansen, M.A.; Gaba, V.; Greenberg, B.M. Higher plants and UV-B radiation: Balancing damage, repair and acclimation. Trends Plant Sci. 1998, 3, 131–135. [Google Scholar] [CrossRef]
- Rozema, J.; Broekman, R.A.; Blokker, P.; Meijkamp, B.B.; de Bakker, N.; van de Staaij, J.; van Beem, A.; Ariese, F.; Kars, S.M. UV-B absorbance and UV-B absorbing compounds (para-coumaric acid) in pollen and sporopollenin: The perspective to track historic UV-B levels. J. Photochem. Photobiol. B Biol. 2001, 62, 108–117. [Google Scholar] [CrossRef]
- Fraser, W.T.; Lomax, B.H.; Jardine, P.E.; Gosling, W.D.; Sephton, M.A. Pollen and spores as a passive monitor of ultraviolet radiation. Front. Ecol. Evol. 2014, 2, 12. [Google Scholar] [CrossRef]
- Fraser, W.T.; Scott, A.C.; Forbs, A.E.S.; Glasspool, I.J.; Plotnick, R.E.; Kenig, F.; Lomax, B.H. Evolutionary stasis of sporopollenin biochemistry revealed by unaltered Pennsylvanian spores. New Phytol. 2012, 196, 397–401. [Google Scholar] [CrossRef]
- Greenberg, B.M.; Wilson, M.I.; Huang, X.; Duxbury, C.L.; Gerhardt, K.E.; Gensemer, R.W. The effects of ultraviolet-B radiation on higher plants. In Plants for Environmental Studies, 1st ed.; Wang, W., Lower, W.R., Gorsuch, J.W., Eds.; CRC Press LLC: Boca Raton, FL, USA, 1997; pp. 1–36. [Google Scholar]
- Rozema, J.; Noordijk, A.J.; Broekman, R.A.; van Beem, A.; Meijkamp, B.M.; de Bakker, N.V.J.; van de Staaij, J.W.M.; Stroetenga, M.; Bohncke, S.J.P.; Konert, M.; et al. (Poly) phenolic compounds in pollen and spores of Antarctic plants as indicators of solar UV-B—A new proxy for the reconstruction of past solar UV-B? Plant Ecol. 2001, 154, 9–26. [Google Scholar] [CrossRef]
- Hidvegi, S.; Racz, F.; Hadi, G.; Gesztesi, L.; Toth, Z. Effect of UV-radiation on the pollen viability of some parental lines of hybrid maize. Cereal Res. Commun. 2009, 37, 349–352. [Google Scholar]
- Flint, S.D.; Caldwell, M.M. Partial inhibition of in vitro pollen germination by simulated solar ultraviolet-B radiation. Ecology 1984, 65, 792–795. [Google Scholar] [CrossRef]
- Santos, A.; Almeida, J.M.; Santos, I.; Salema, R. Biochemical and ultrastructural changes in pollen of Zea mays L. grown under enhanced UV-B radiation. Ann. Bot. 1998, 82, 641–645. [Google Scholar] [CrossRef]
- Aylor, D.E. Survival of maize (Zea mays) pollen exposed in the atmosphere. Agric. For. Meteorol. 2004, 123, 125–133. [Google Scholar] [CrossRef]
- He, J.M.; Bai, X.L.; Wang, R.B.; Cao, B.; She, X.P. The involvement of nitric oxide in ultraviolet-B-inhibited pollen germination and tube growth of Paulownia tomentosa in vitro. Physiol. Plant. 2007, 131, 273–282. [Google Scholar] [CrossRef]
- Torabinejad, J.; Caldwell, M.M.; Flint, S.D.; Durham, S. Susceptibility of pollen to UV-B radiation: An assay of 34 taxa. Am. J. Bot. 1998, 85, 360–369. [Google Scholar] [CrossRef]
- Chesnokov, Y.V.; Manteuffel, R. Dose effect of UV-B irradiation on pollen tube growth and seed-specific promoter activities in irradiated pollen grains of Nicotiana plumbaginifolia. Sex. Plant Reprod. 2000, 12, 361–364. [Google Scholar] [CrossRef]
- He, J.M.; Liu, Z.H.; Xu, H.; She, X.P.; Huang, C. The involvement of hydrogen peroxide in UV-B-inhibited pollen germination and tube growth of Paeonia suffruticosa and Paulownia tomentosa in vitro. Plant Growth Regul. 2006, 49, 199–208. [Google Scholar] [CrossRef]
- Varela, M.C. Phenolic compounds: Natural antioxidants, other benefices and future perspectives. Austin Biomol. Open Access 2016, 1, 1001. [Google Scholar]
- Mohammadrezakhani, S.; Hajilou, J.; Rezanejad, F. Evaluation of phenolic and flavonoid compounds in pollen grains of three Citrus species in response to low temperature. Grana 2018, 57, 214–222. [Google Scholar] [CrossRef]
- Rezanejad, F. Air pollution effects on flavonoids in pollen grains of some ornamental plants. Turk. J. Bot. 2012, 36, 49–54. [Google Scholar] [CrossRef]
- Baskar, V.; Venkatesh, R.; Ramalingam, S. Flavonoids (antioxidants systems) in higher plants and their response to stresses. In Antioxidants and Antioxidant Enzymes in Higher Plants, 1st ed.; Baskar, V., Venkatesh, R., Ramalingam, S., Eds.; Springer: Cham, Switzerland, 2018; pp. 253–268. [Google Scholar] [CrossRef]
- Kim, Y.S.; Song, K.S.; Cheong, H.S. Effects of flavonoids on pollen tube growth in Arabidopsis thaliana. J. Plant Biol. 1996, 39, 273–278. [Google Scholar]
- Ylstra, B.; Touraev, A.; Moreno, R.M.B.; Stöger, E.; Van Tunen, A.J.; Vicente, O.; Mol, J.N.M.; Heberle-Bors, E. Flavonols stimulate development, germination, and tube growth of tobacco pollen. Plant Physiol. 1992, 100, 902–907. [Google Scholar] [CrossRef] [PubMed]
- Seddon, A.W.; Festi, D.; Robson, T.M.; Zimmermann, B. Fossil pollen and spores as a tool for reconstructing ancient solar-ultraviolet irradiance received by plants: An assessment of prospects and challenges using proxy-system modelling. Photochem. Photobiol. Sci. 2019, 18, 275–294. [Google Scholar] [CrossRef] [PubMed]
- Bell, B.A.; Bishop, T.H.; Fletcher, W.J.; Ryan, P.; Ilmen, R. Cedrus atlantica pollen morphology and investigation of grain size variability using laser diffraction granulometry. Palynology 2018, 42, 339–353. [Google Scholar] [CrossRef]
- Lomax, B.H.; Fraser, W.T. Palaeoproxies: Botanical monitors and recorders of atmospheric change. Palaeontology 2015, 58, 759–768. [Google Scholar] [CrossRef]
- Musil, C.F. Differential effects of elevated ultraviolet-B radiation on the photochemical and reproductive performances of dicotyledonous and monocotyledonous arid-environment ephemerals. Plant Cell Environ. 1995, 18, 844–854. [Google Scholar] [CrossRef]
- Muthreich, F.; Zimmermann, B.; Birks, H.J.B.; Vila-Viçosa, C.M.; Seddon, A.W. Chemical variations in Quercus pollen as a tool for taxonomic identification: Implications for long-term ecological and biogeographical research. J. Biogeogr. 2020, 47, 1298–1309. [Google Scholar] [CrossRef]
- Seddon, A.W.; Festi, D.; Nieuwkerk, M.; Gya, R.; Hamre, B.; Krüger, L.C.; Östman, S.A.H.; Robson, T.M. Pollen-chemistry variations along elevation gradients and their implications for a proxy for UV-B radiation in the plant-fossil record. J. Ecol. 2021, 109, 3060–3073. [Google Scholar] [CrossRef]
- Musil, C.F.; Wand, S.J.E. Differential stimulation of an arid-environment winter ephemeral Dimorphotheca pluvialis (L.) Moench by ultraviolet-B radiation under nutrient limitation. Plant Cell Environ. 1994, 17, 245–255. [Google Scholar] [CrossRef]
- Novara, C.; Ascari, L.; La Morgia, V.; Reale, L.; Genre, A.; Siniscalco, C. Viability and germinability in long term storage of Corylus avellana pollen. Sci. Hortic. 2017, 214, 295–303. [Google Scholar] [CrossRef]
- Brewbaker, J.L.; Kwack, B.H. The essential role of calcium ion in pollen germination and pollen tube growth. Am. J. Bot. 1963, 50, 859–865. [Google Scholar] [CrossRef]
- Dai, Q.; Shaobing, P.; Chavez, A.Q.; Vergara, B.S. Intraspecific responses of 188 rice cultivars to enhanced UVB radiation. Environ. Exp. Bot. 1994, 34, 433–442. [Google Scholar] [CrossRef]
- Oroian, M.; Ursachi, F.; Dranca, F. Ultrasound-assisted extraction of polyphenols from crude pollen. Antioxidants 2020, 9, 322. [Google Scholar] [CrossRef]
- Chatatikun, M.; Chiabchalard, A. Phytochemical screening and free radical scavenging activities of orange baby carrot and carrot (Daucus carota Linn.) root crude extracts. J. Chem. Pharm. 2013, 5, 97–102. [Google Scholar]
- Clarke, G.; Ting, K.N.; Wiart, C.; Fry, J. High correlation of 2, 2-diphenyl-1-picrylhydrazyl (DPPH) radical scavenging, ferric reducing activity potential and total phenolics content indicates redundancy in use of all three assays to screen for antioxidant activity of extracts of plants from the Malaysian rainforest. Antioxidants 2013, 2, 1–10. [Google Scholar] [CrossRef]
- Yang, H.; Dong, Y.; Du, H.; Shi, H.; Peng, Y.; Li, X. Antioxidant compounds from propolis collected in Anhui, China. Molecules 2011, 16, 3444–3455. [Google Scholar] [CrossRef] [PubMed]
- Mundargi, R.C.; Potroz, M.G.; Park, J.H.; Seo, J.; Lee, J.H.; Cho, N.J. Extraction of sporopollenin exine capsules from sunflower pollen grains. RSC Adv. 2016, 6, 16533–16539. [Google Scholar] [CrossRef]
- Ruhland, C.T.; Day, T.A. Effects of ultraviolet-B radiation on leaf elongation, production and phenylpropanoid concentrations of Deschampsia antarctica and Colobanthus quitensis in Antarctica. Physiol. Plant. 2000, 109, 244–251. [Google Scholar] [CrossRef]
- Xiong, F.; Komenda, J.; Kopecký, J.; Nedbal, L. Strategies of ultraviolet-B protection in microscopic algae. Physiol. Plant. 1997, 100, 378–388. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Times | ||||
|---|---|---|---|---|
| 1 h | 2 h | 3 h | ||
| Treatment distances | 10 cm | −39,941 | −89,516 | −122,685 |
| 20 cm | −42,664 | −146,282 | −106,176 | |
| 30 cm | −28,603 | −46,672 | −94,886 | |
| 40 cm | −11,915 | −42,361 | −67,687 | |
| Sensitive < −104, 7 < Intermediate < −63, 2 < Tolerant < −21, 7 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Çetinbaş-Genç, A.; Toksöz, O.; Piccini, C.; Kilin, Ö.; Sesal, N.C.; Cai, G. Effects of UV-B Radiation on the Performance, Antioxidant Response and Protective Compounds of Hazelnut Pollen. Plants 2022, 11, 2574. https://doi.org/10.3390/plants11192574
Çetinbaş-Genç A, Toksöz O, Piccini C, Kilin Ö, Sesal NC, Cai G. Effects of UV-B Radiation on the Performance, Antioxidant Response and Protective Compounds of Hazelnut Pollen. Plants. 2022; 11(19):2574. https://doi.org/10.3390/plants11192574
Chicago/Turabian StyleÇetinbaş-Genç, Aslıhan, Orçun Toksöz, Chiara Piccini, Özkan Kilin, Nüzhet Cenk Sesal, and Giampiero Cai. 2022. "Effects of UV-B Radiation on the Performance, Antioxidant Response and Protective Compounds of Hazelnut Pollen" Plants 11, no. 19: 2574. https://doi.org/10.3390/plants11192574
APA StyleÇetinbaş-Genç, A., Toksöz, O., Piccini, C., Kilin, Ö., Sesal, N. C., & Cai, G. (2022). Effects of UV-B Radiation on the Performance, Antioxidant Response and Protective Compounds of Hazelnut Pollen. Plants, 11(19), 2574. https://doi.org/10.3390/plants11192574