
Pea Breeding for Resistance to Rhizospheric Pathogens




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
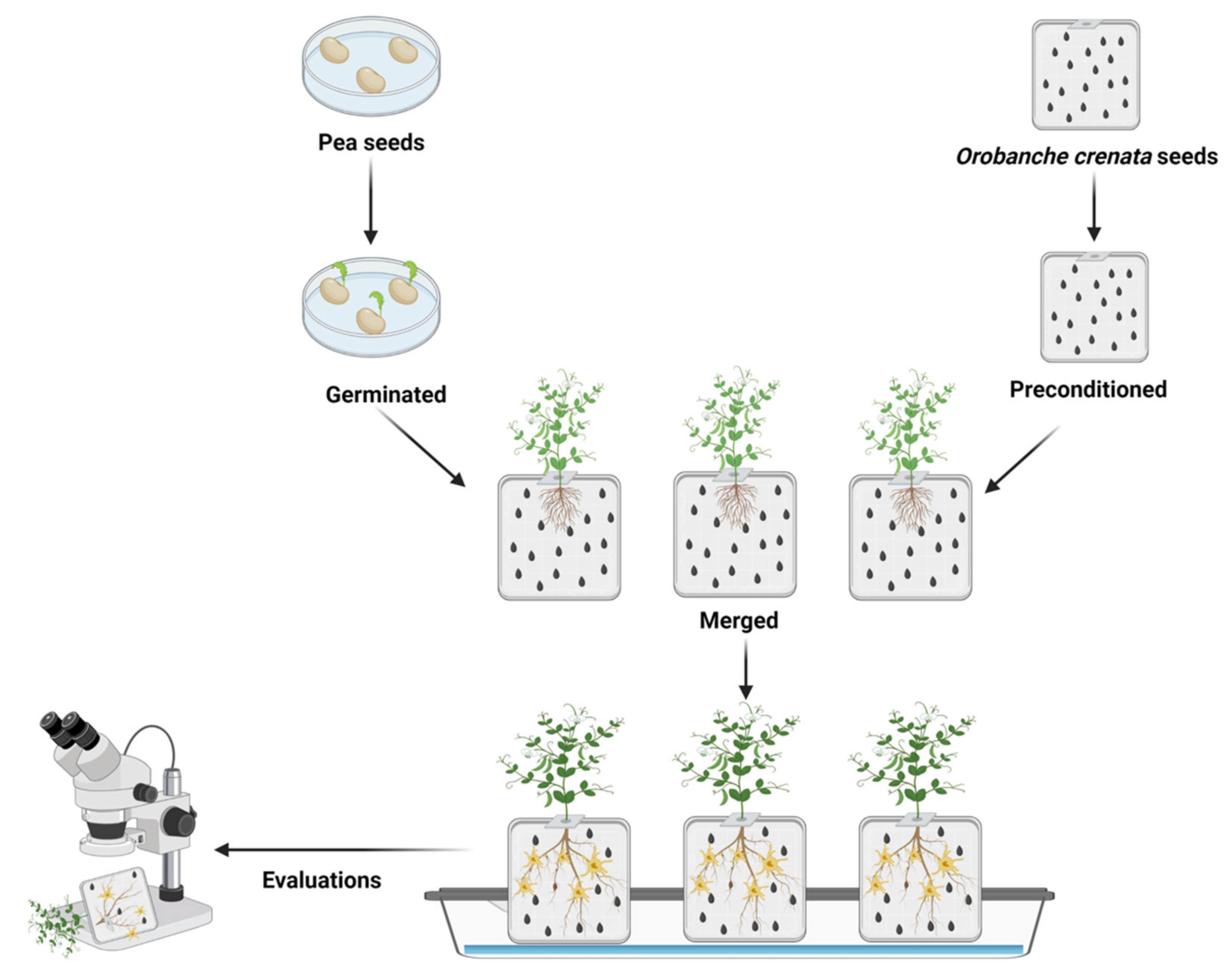
:1. Introduction
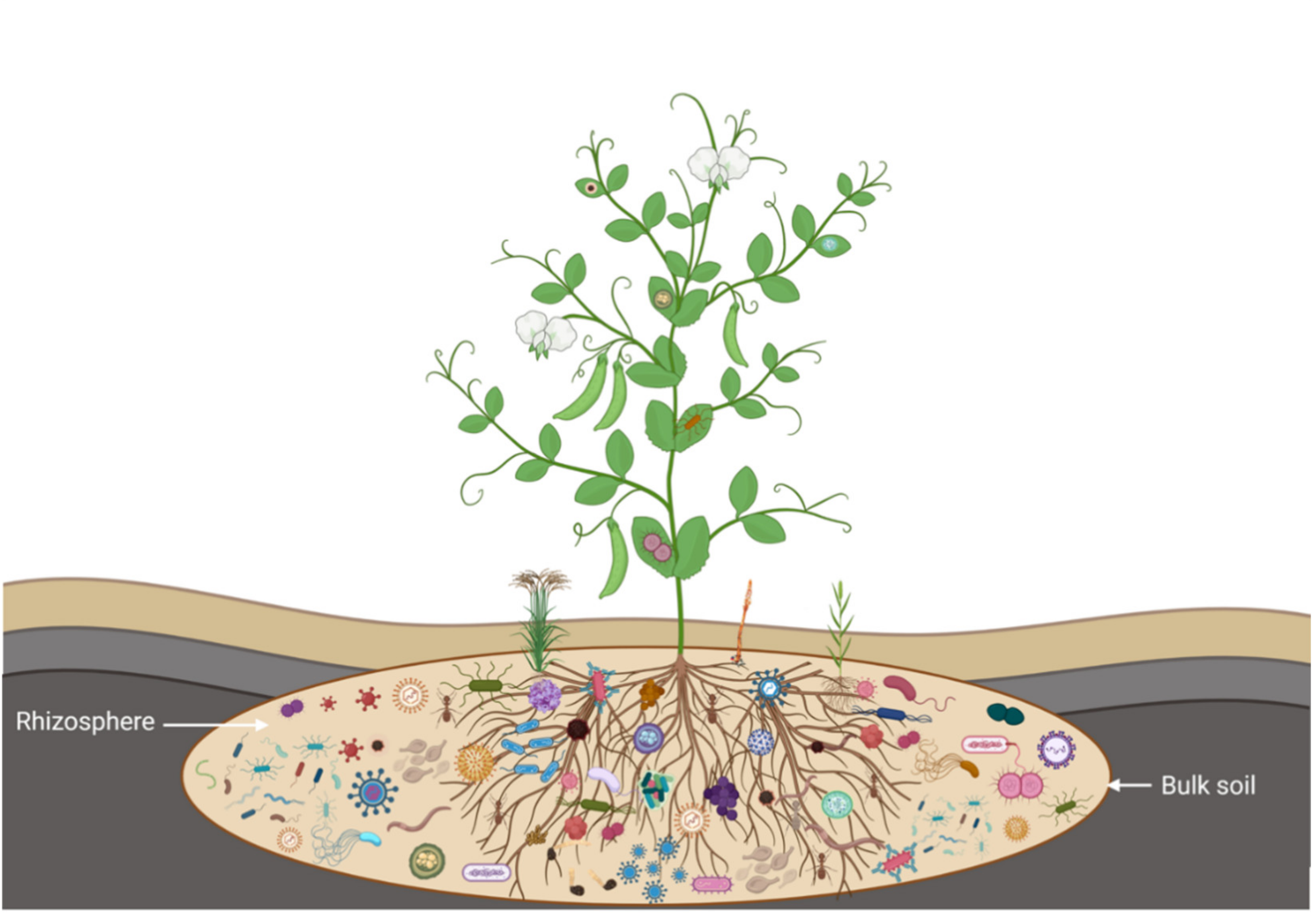
2. Host Pea-Rhizosphere-Microbial Interactions and Stress Amelioration
3. Pea Rhizospheric Diseases
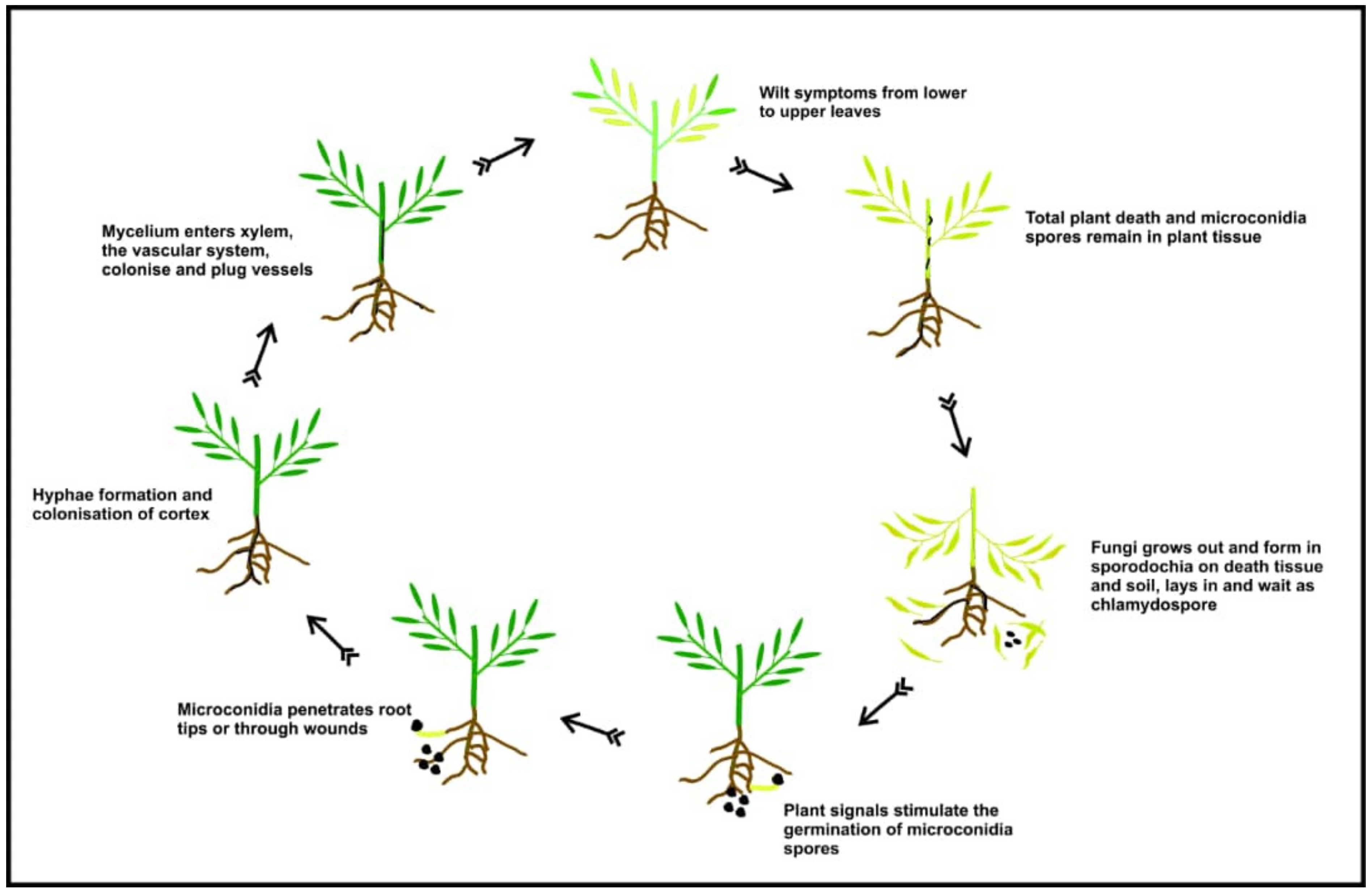
3.1. Fusarium Wilt
3.2. The Rots Complex
3.2.1. Aphanomyces Root Rot
3.2.2. Fusarium Root Rot
3.2.3. Black Root Rot
3.2.4. Rhizoctonia Root Rot
3.2.5. Pythium Seed/Seedling Rot
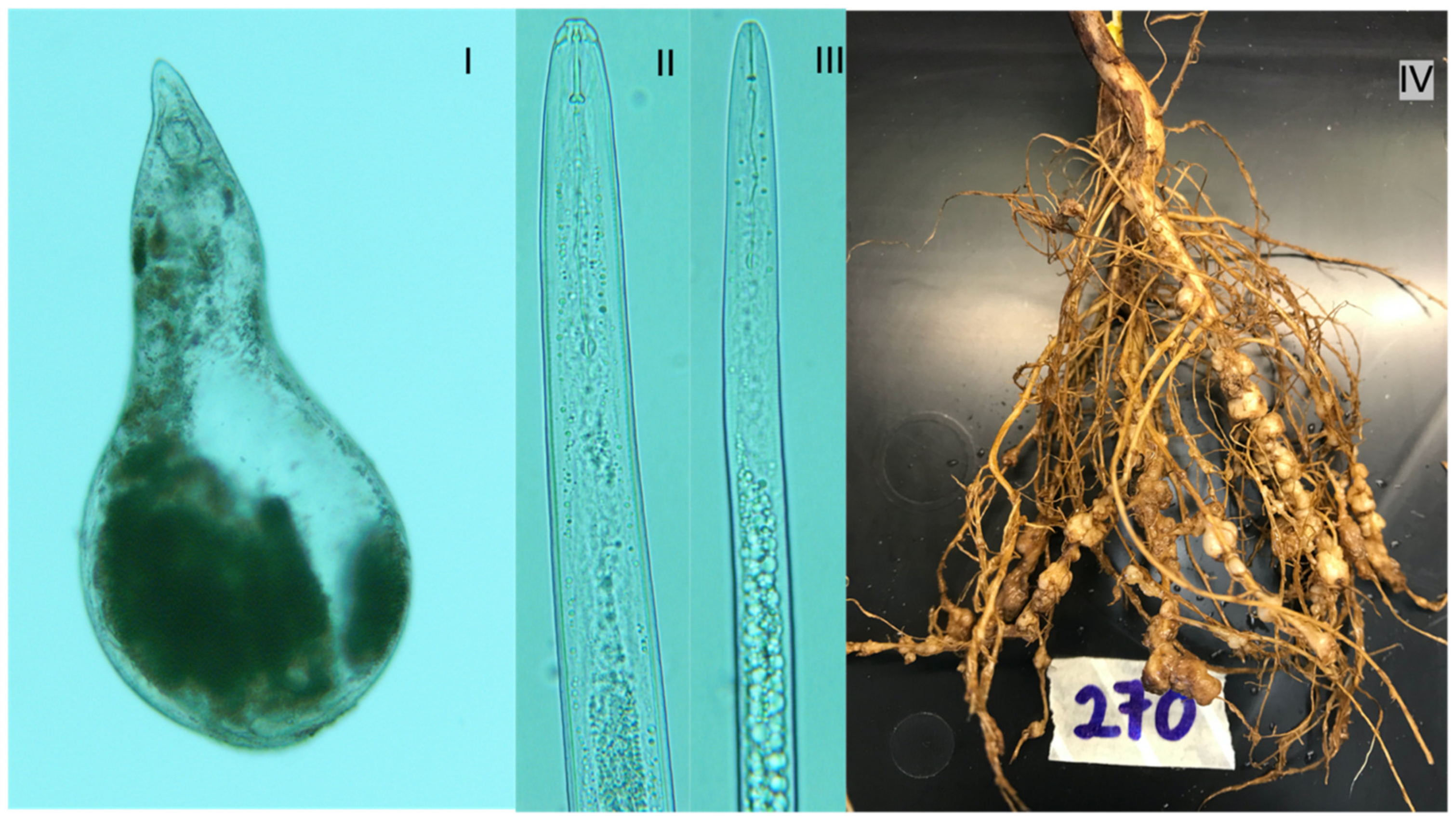
3.3. The Nematode Complex
3.3.1. Pea Cyst Nematodes
3.3.2. Root Knot Nematodes
3.3.3. Root Lesion Nematodes
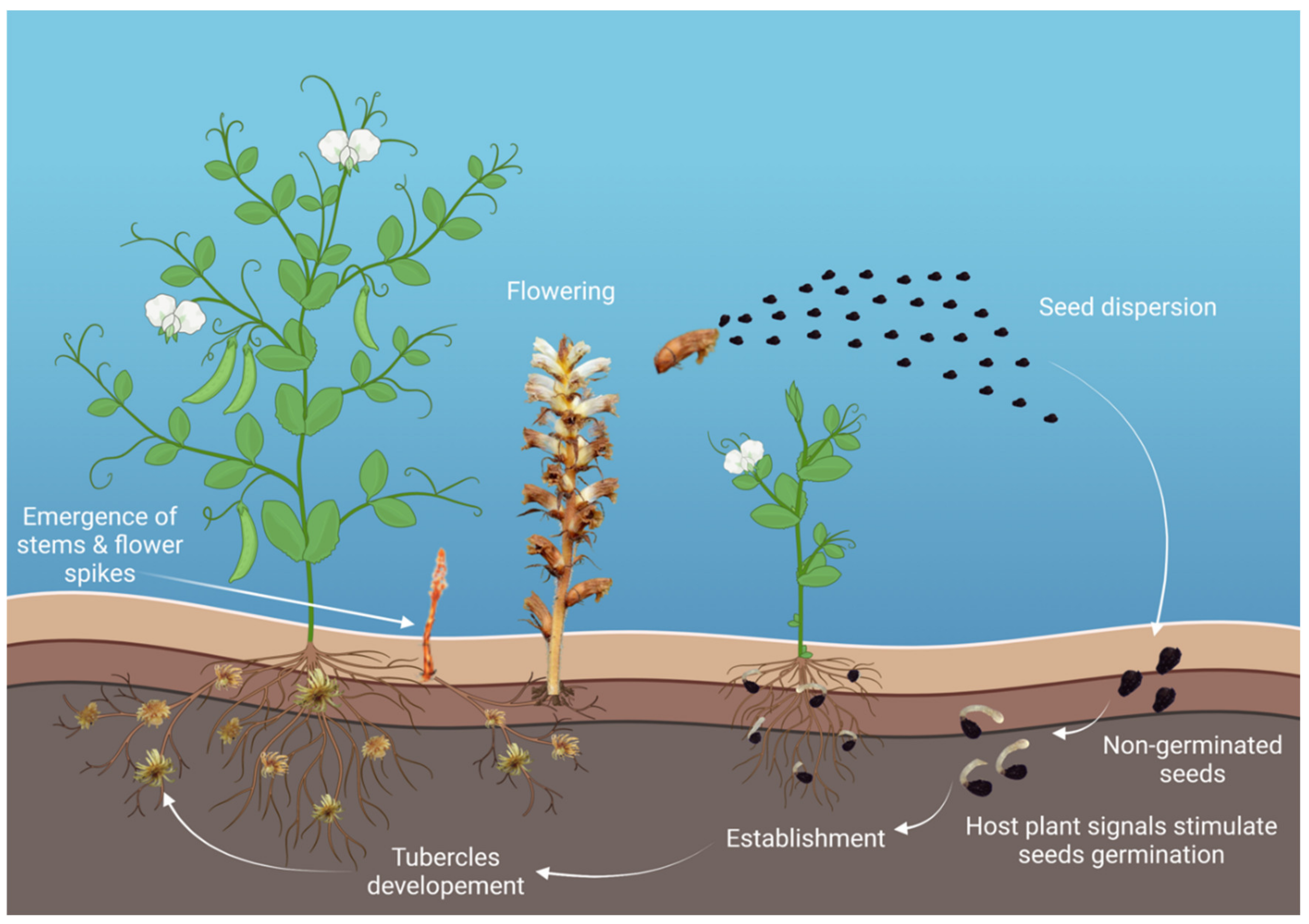
3.4. Root Parasitic Weeds: Broomrapes
4. Breeding Enabling Approaches for Disease Resistance
4.1. Phenotyping
4.1.1. Field and Controlled Condition Phenotyping
4.1.2. High-Throughput Phenotyping
4.1.3. Innovative Rhizotrons for Rhizosphere Phenotyping
4.2. Trait Discovery and Pre-Breeding for Resistance Breeding
4.3. Genotyping: Genomic Tools and Genetic Breeding Approaches
5. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Smýkal, P.; Aubert, G.; Burstin, J.; Coyne, C.J.; Ellis, N.T.; Flavell, A.J.; Ford, R.; Hýbl, M.; Macas, J.; Neumann, P.; et al. Pea (Pisum sativum L.) in the genomic era. Agronomy 2012, 2, 74–115. [Google Scholar] [CrossRef]
- Rubiales, D.; Ambrose, M.J.; Domoney, C.; Burstin, J. Pea. In Genetics, Genomics and Breeding of Cool Season Grain Legumes; Pérez-de-la-Vega, M., Torres, A.M., Cubero, J.I., Kole, C., Eds.; Science Publishers: Enfield, NH, USA, 2011; pp. 1–49. ISBN 978-1-57808-765-5. [Google Scholar]
- Trněný, O.; Brus, J.; Hradilová, I.; Rathore, A.; Das, R.R.; Kopecký, P.; Coyne, C.J.; Reeves, P.; Richards, C.; Smýkal, P. Molecular evidence for two domestication events in the pea crop. Genes 2018, 9, 535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, A.K.; Rubiales, D.; Wang, Y.; Fang, P.; Sun, T.; Liu, N.; Xu, P. Omics resources and omics-enabled approaches for achieving high productivity and improved quality in pea (Pisum sativum L.). Theor. Appl. Genet. 2021, 134, 755–776. [Google Scholar] [CrossRef] [PubMed]
- Coyne, C.J.; Kumar, S.; von Wettberg, E.J.B.; Marques, E.; Berger, J.D.; Redden, R.J.; Ellis, T.N.; Brus, J.; Zablatzká, L.; Smýkal, P. Potential and limits of exploitation of crop wild relatives for pea, lentil, and chickpea improvement. Legum. Sci. 2020, 2, e36. [Google Scholar] [CrossRef] [Green Version]
- Pratap, A.; Das, A.; Kumar, S.; Gupta, S. Current Perspectives on Introgression Breeding in Food Legumes. Front. Plant Sci. 2021, 11, 589189. [Google Scholar] [CrossRef] [PubMed]
- Cobos, M.J.; Satovic, Z.; Rubiales, D.; Fondevilla, S. Er3 gene, conferring resistance to powdery mildew in pea, is located in pea LGIV. Euphytica 2018, 214, 203. [Google Scholar] [CrossRef]
- Rubiales, D.; Fondevilla, S.; Fernández-Aparicio, M. Development of pea breeding lines with resistance to Orobanche crenata derived from pea landraces and wild Pisum spp. Agronomy 2021, 11, 36. [Google Scholar] [CrossRef]
- Rubiales, D.; Osuna-Caballero, S.; González-Bernal, M.J.; Cobos, M.J.; Flores, F. Pea Breeding Lines Adapted to Autumn Sowings in Broomrape Prone Mediterranean Environments. Agronomy 2021, 11, 769. [Google Scholar] [CrossRef]
- Das, A.; Parihar, A.K.; Saxena, D.; Singh, D.; Singha, K.D.; Kushwaha, K.P.S.; Chand, R.; Bal, R.S.; Chandra, S.; Gupta, S. Deciphering genotype-by- Environment interaction for targeting test environments and rust resistant genotypes in field pea (Pisum sativum L.). Front. Plant Sci. 2019, 10, 825. [Google Scholar] [CrossRef]
- Varshney, R.K.; Barmukh, R.; Roorkiwal, M.; Qi, Y.; Kholova, J.; Tuberosa, R.; Reynolds, M.P.; Tardieu, F.; Siddique, K.H. Breeding custom-designed crops for improved drought adaptation. Adv. Genet. 2021, 2, e202100017. [Google Scholar] [CrossRef]
- Rubiales, D.; González-Bernal, M.J.; Warkentin, T.; Bueckert, R.; Vaz Patto, M.C.; McPhee, K.; McGee, R.; Smýkal, P. Advances in pea breeding. In Achieving Sustainable Cultivation of Vegetables; Hochmuth, G., Ed.; Burleig Dodds Science Publishing Limited: Cambridge, UK, 2019; pp. 575–606. [Google Scholar] [CrossRef]
- FAOSTAT, Statistical Database. Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/faostat (accessed on 29 June 2022).
- Hickey, L.T.; Hafeez, A.N.; Robinson, H.; Jackson, S.A.; Leal-Bertioli, S.C.M.; Tester, M.; Gao, C.; Godwin, I.D.; Hayes, B.J.; Wulff, B.B. Breeding crops to feed 10 billion. Nat. Biotechnol. 2019, 37, 744–754. [Google Scholar] [CrossRef] [Green Version]
- Varshney, R.K.; Bohra, A.; Roorkiwal, M.; Barmukh, R.; Cowling, W.A.; Chitikineni, A.; Lam, H.M.; Hickey, L.T.; Croser, J.S.; Bayer, P.E.; et al. Fast-forward breeding for a food-secure world. Trends Genet. 2021, 37, 1124–1136. [Google Scholar] [CrossRef]
- Smýkal, P.; Varshney, R.K.; Singh, V.K.; Coyne, C.J.; Domoney, C.; Kejnovský, E.; Warkentin, T. From Mendel’s discovery on pea to today’s plant genetics and breeding: Commemorating the 150th anniversary of the reading of Mendel’s discovery. Theor. Appl. Genet. 2016, 129, 2267–2280. [Google Scholar] [CrossRef]
- Graham, P.H.; Vance, C.P. Update on Legume Utilization Legumes: Importance and Constraints to Greater Use. Plant Physiol. 2003, 131, 872–877. [Google Scholar] [CrossRef] [Green Version]
- Kreplak, J.; Madoui, M.A.; Cápal, P.; Novák, P.; Labadie, K.; Aubert, G.; Gali, K.K. A reference genome for pea provides insight into legume genome evolution. Nat. Genet. 2019, 51, 1411–1422. [Google Scholar] [CrossRef]
- Zander, P.; Amjath-Babu, T.S.; Preissel, S.; Reckling, M.; Bues, A.; Schläfke, N.; Kuhlman, T.; Bachinger, J.; Uthes, S.; Stoddard, F.; et al. Grain legume decline and potential recovery in European agriculture: A review. Agron. Sustain. Dev. 2016, 36, 26. [Google Scholar] [CrossRef] [Green Version]
- Ojiewo, C.; Monyo, E.; Desmae, H.; Boukar, O.; Mukankusi-Mugisha, C.; Thudi, M.; Pandey, M.K.; Saxena, R.K.; Gaur, P.M.; Chaturvedi, S.K.; et al. Genomics, genetics and breeding of tropical legumes for better livelihoods of smallholder farmers. Plant Breed. 2019, 138, 487–499. [Google Scholar] [CrossRef]
- Döring, T.F.; Rosslenbroich, D.; Giese, C.; Athmann, M.; Watson, C.; Vágó, I.; Kátai, J.; Tállai, M.; Bruns, C. Disease suppressive soils vary in resilience to stress. Appl. Soil Ecol. 2020, 149, 103482. [Google Scholar] [CrossRef]
- Vishwakarma, K.; Kumar, N.; Shandilya, C.; Mohapatra, S.; Bhayana, S.; Varma, A. Revisiting Plant–Microbe Interactions and Microbial Consortia Application for Enhancing Sustainable Agriculture: A Review. Front. Microbiol. 2020, 11, 560406. [Google Scholar] [CrossRef]
- Müller, D.B.; Vogel, C.; Bai, Y.; Vorholt, J.A. The plant microbiota: Systems-level insights and perspectives. Annu. Rev. Genet. 2016, 50, 211–234. [Google Scholar] [CrossRef] [Green Version]
- Latati, M.; Aouiche, A.; Tellah, S.; Laribi, A.; Benlahrech, S.; Kaci, G.; Ouarem, F.; Ounane, S.M. Intercropping maize and common bean enhances microbial carbon and nitrogen availability in low phosphorus soil under Mediterranean conditions. Eur. J. Soil Biol. 2017, 80, 9–18. [Google Scholar] [CrossRef]
- Hamel, C.; Gan, Y.T.; Sokolski, S.; Bainard, L.D. High frequency cropping of pulses modifies soil nitrogen level and the rhizosphere bacterial microbiome in 4-year rotation systems of the semiarid prairie. Appl. Soil Ecol. 2018, 126, 47–56. [Google Scholar] [CrossRef]
- Wille, L.; Messmer, M.M.; Studer, B.; Hohmann, P. Insights to plant–microbe interactions provide opportunities to improve resistance breeding against root diseases in grain legumes. Plant Cell Environ. 2019, 42, 20–40. [Google Scholar] [CrossRef] [Green Version]
- Chaudhari, D.; Rangappa, K.; Das, A.; Layek, J.; Basavaraj, S.; Kandpal, B.K.; Shouche, Y.; Rahi, P. Pea (Pisum sativum L.) plant shapes its rhizosphere microbiome for nutrient uptake and stress amelioration in acidic soils of the North-East region of India. Front. Microbiol. 2020, 11, 968. [Google Scholar] [CrossRef]
- Dolgikh, E.A.; Kusakin, P.G.; Kitaeva, A.B.; Tsyganova, A.V.; Kirienko, A.N.; Leppyanen, I.V.; Dolgikh, A.V.; Ilina, E.L.; Demchenko, K.N.; Tikhonovich, I.A.; et al. Mutational analysis indicates that abnormalities in rhizobial infection and subsequent plant cell and bacteroid differentiation in pea (Pisum sativum) nodules coincide with abnormal cytokinin responses and localization. Ann. Bot. 2020, 125, 905–923. [Google Scholar] [CrossRef]
- Rodriguez-Carres, M.; White, G.; Tsuchiya, D.; Taga, M.; VanEtten, H.D. The supernumerary chromosome of Nectria haematococca that carries pea-pathogenicity-related genes also carries a trait for pea rhizosphere competitiveness. Appl. Environ. Microbiol. 2008, 74, 3849–3856. [Google Scholar] [CrossRef] [Green Version]
- Belimov, A.A.; Shaposhnikov, A.I.; Syrova, D.S.; Kichko, A.A.; Guro, P.V.; Yuzikhin, O.S.; Azarova, T.S.; Sazanova, A.L.; Sekste, E.A.; Litvinskiy, V.A.; et al. The role of symbiotic microorganisms, nutrient uptake and rhizosphere bacterial community in response of pea (Pisum sativum L.) genotypes to elevated Al concentrations in soil. Plants 2020, 9, 1801. [Google Scholar] [CrossRef]
- Bani, M.; Cimmino, A.; Evidente, A.; Rubiales, D.; Rispail, N. Pisatin involvement in the variation of inhibition of Fusarium oxysporum f. sp. pisi spore germination by root exudates of Pisum spp. germplasm. Plant Pathol. 2018, 67, 1046–1054. [Google Scholar] [CrossRef]
- Oyserman, B.O.; Flores, S.S.; Griffioen, T.; Pan, X.; van der Wijk, E.; Pronk, L.; Lokhorst, W.; Nurfikari, A.; Paulson, J.N.; Movassagh, M.; et al. Disentangling the genetic basis of rhizosphere microbiome assembly in tomato. Nat. Commun. 2022, 13, 3228. [Google Scholar] [CrossRef]
- Whipps, J.M. Microbial interactions and biocontrol in the rhizosphere. J. Exp. Bot. 2001, 52, 487–511. [Google Scholar] [CrossRef]
- Williamson-Benavides, B.A.; Sharpe, R.M.; Nelson, G.; Bodah, E.T.; Porter, L.D.; Dhingra, A. Identification of Root Rot Resistance QTLs in Pea Using Fusarium solani f. sp. pisi-Responsive Differentially Expressed Genes. Front. Genet. 2021, 12, 1379. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Aparicio, M.; García-Garrido, J.M.; Ocampo, J.A.; Rubiales, D. Colonisation of field pea roots by arbuscular mycorrhizal fungi reduces Orobanche and Phelipanche species seed germination. Weed Res. 2010, 50, 262–268. [Google Scholar] [CrossRef]
- Harkes, P.; Van Steenbrugge, J.J.M.; Van Den Elsen, S.J.J.; Suleiman, A.K.A.; De Haan, J.J.; Holterman, M.H.M.; Helder, J. Shifts in the active rhizobiome paralleling low Meloidogyne chitwoodi densities in fields under prolonged organic soil management. Front. Plant Sci. 2020, 10, 1697. [Google Scholar] [CrossRef] [PubMed]
- Hawes, M.; Allen, C.; Turgeon, B.G.; Curlango-Rivera, G.; Minh Tran, T.; Huskey, D.A.; Xiong, Z. Root border cells and their role in plant defense. Annu. Rev. Phytopathol. 2016, 54, 143–161. [Google Scholar] [CrossRef] [PubMed]
- Camborde, L.; Kiselev, A.; Pel, M.J.C.; Le Ru, A.; Jauneau, A.; Pouzet, C.; Dumas, B.; Gaulin, E. An oomycete effector targets a plant RNA helicase involved in root development and defense. New Phytol. 2021, 233, 2232–2248. [Google Scholar] [CrossRef] [PubMed]
- Wille, L.; Messmer, M.M.; Bodenhausen, N.; Studer, B.; Hohmann, P. Heritable Variation in Pea for Resistance Against a Root Rot Complex and Its Characterization by Amplicon Sequencing. Front. Plant Sci. 2020, 11, 542153. [Google Scholar] [CrossRef]
- Tosi, M.; Mitter, E.K.; Gaiero, J.; Dunfield, K. It takes three to tango: The importance of microbes, host plant, and soil management to elucidate manipulation strategies for the plant microbiome. Can. J. Microbiol. 2020, 66, 413–433. [Google Scholar] [CrossRef]
- Dubey, S.; Sharma, S. Rhizospheric Engineering by Plant-Mediated Indirect Selection of Microbiome for Agricultural Sustainability. Crit. Rev. Plant Sci. 2021, 40, 379–397. [Google Scholar] [CrossRef]
- Hartmann, A.; Rothballer, M.; Schmid, M. Lorenz Hiltner, a pioneer in rhizosphere microbial ecology and soil bacteriology research. Plant Soil 2008, 312, 7–14. [Google Scholar] [CrossRef]
- Lucas, M.R.; Huynh, B.L.; da Silva Vinholes, P.; Cisse, N.; Drabo, I.; Ehlers, J.D.; Roberts, P.A.; Close, T.J. Association studies and legume synteny reveal haplotypes determining seed size in Vigna unguiculata. Front. Plant Sci. 2013, 4, 95. [Google Scholar] [CrossRef] [Green Version]
- Niks, R.E.; Parlevliet, J.E.; Lindhout, P.; Bai, Y. Breeding Crops with Resistance to Diseases and Pests; Wageningen Academic Publishers: Wageningen, The Netherlands, 2019. [Google Scholar]
- Rubiales, D. Developing pest- and disease-resistant cultivars of grain legumes. In Achieving Sustainable Cultivation of Grain Legumes. Volume 1: Advances in Breeding and Cultivation Techniques; Sivasankar, S., Bergvinson, D., Gaur, P., Kumar, S., Beebe, S., Tamò, M., Eds.; Burleig Dodds Science Publishing: Cambridge, UK, 2018; ISBN 978-1-78676-136-1. [Google Scholar]
- Raaijmakers, J.M.; Paulitz, T.C.; Steinberg, C.; Alabouvette, C.; Moënne-Loccoz, Y. The rhizosphere: A playground and battlefield for soilborne pathogens and beneficial microorganisms. Plant Soil 2009, 321, 341–361. [Google Scholar] [CrossRef] [Green Version]
- O’Keeffe, K.R.; Carbone, I.; Jones, C.D.; Mitchell, C.E. Plastic potential: How the phenotypes and adaptations of pathogens are influenced by microbial interactions within plants. Curr. Opin. Plant Biol. 2017, 38, 78–83. [Google Scholar] [CrossRef]
- Lyu, D.; Msimbira, L.A.; Nazari, M.; Antar, M.; Pagé, A.; Shah, A.; Monjezi, N.; Zajonc, J.; Tanney, C.A.; Backer, R.; et al. The coevolution of plants and microbes underpins sustainable agriculture. Microorganisms 2021, 9, 1036. [Google Scholar] [CrossRef]
- Infantino, A.; Kharrat, M.; Riccioni, L.; Coyne, C.J.; McPhee, K.E.; Grünwald, N.J. Screening techniques and sources of resistance to root diseases in cool season food legumes. Euphytica 2006, 147, 201–221. [Google Scholar] [CrossRef]
- Foster, K.; You, M.P.; Nietschke, B.; Edwards, N.; Barbetti, M.J. Soilborne root disease pathogen complexes drive widespread decline of subterranean clover pastures across diverse climatic zones. Crop Pasture Sci. 2017, 68, 33–44. [Google Scholar] [CrossRef]
- Rubiales, D.; Fondevilla, S.; Chen, W.; Gentzbittel, L.; Higgins, T.J.V.; Castillejo, M.A.; Singh, K.B.; Rispail, N. Achievements and Challenges in Legume Breeding for Pest and Disease Resistance. CRC. Crit. Rev. Plant Sci. 2015, 34, 195–236. [Google Scholar] [CrossRef] [Green Version]
- Zitnick-Anderson, K.; del Río Mendoza, L.E.; Forster, S.; Pasche, J.S. Associations among the communities of soil-borne pathogens, soil edaphic properties and disease incidence in the field pea root rot complex. Plant Soil 2020, 457, 339–354. [Google Scholar] [CrossRef]
- Di Pietro, A.; Madrid, M.P.; Caracuel, Z.; Delgado-Jarana, J.; Roncero, M.I.G. Fusarium oxysporum: Exploring the molecular arsenal of a vascular wilt fungus. Mol. Plant Pathol. 2003, 4, 315–325. [Google Scholar] [CrossRef]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual, 1st ed.; Blackwell Publishing: Ames, IA, USA, 2006; Volume 22, pp. 3–274. [Google Scholar]
- Bani, M.; Rubiales, D.; Rispail, N. A detailed evaluation method to identify sources of quantitative resistance to Fusarium oxysporum f. sp. pisi race 2 within a Pisum spp. germplasm collection. Plant Pathol. 2012, 61, 532–542. [Google Scholar] [CrossRef]
- Willsey, T.; Patey, J.; Vucurevich, C.; Chatterton, S.; Carcamo, H. Evaluation of foliar and seed treatments for integrated management of root rot and pea leaf weevil in field pea and faba bean. Crop Prot. 2021, 143, 105538. [Google Scholar] [CrossRef]
- Sampaio, A.M.; De Sousa Araújo, S.; Rubiales, D.; Patto, M.C.V. Fusarium wilt management in legume crops. Agronomy 2020, 10, 1073. [Google Scholar] [CrossRef]
- Kraft, J.M. Fusarium wilt of peas (a review). Agronomie 1994, 14, 561–567. [Google Scholar] [CrossRef] [Green Version]
- Mcphee, K.E.; Tullu, A.; Kraft, J.M.; Muehlbauer, F.J. Resistance to Fusarium Wilt Race 2 in the Pisum Core Collection. J. Am. Soc. Hortic. Sci. 1999, 124, 28–31. [Google Scholar] [CrossRef] [Green Version]
- Bani, M.; Pérez-de-Luque, A.; Rubiales, D.; Rispail, N. Physical and chemical barriers in root tissues contribute to quantitative resistance to Fusarium oxysporum f. sp. pisi in Pea. Front. Plant Sci. 2018, 9, 199. [Google Scholar] [CrossRef]
- Kraft, J.M.; Pfleger, F.L. Compendium of Pea Diseases and Pests, 2nd ed.; American Phytopathological Society (APS Press): St. Paul, MN, USA, 2001. [Google Scholar]
- Shubha, K.; Dhar, S.; Choudhary, H.; Dubey, S.C.; Sharma, R.K. Identification of resistant sources and inheritance of Fusarium wilt resistance in garden pea (Pisum sativum ssp. hortense). Indian J. Hortic. 2016, 73, 356–361. [Google Scholar] [CrossRef]
- Harveson, R.M.; Pasche, J.S.; Porter, L.; Chen, W.; Burrows, M. Compendium of Pea Diseases and Pests, 3rd ed.; American Phytopathological Society: St. Paul, MN, USA, 2021; ISBN 978-0-89054-655-0. [Google Scholar]
- Leslie, J.F.; Anderson, L.L.; Bowden, R.L.; Lee, Y.W. Inter- and intra-specific genetic variation in Fusarium. Int. J. Food Microbiol. 2007, 119, 25–32. [Google Scholar] [CrossRef]
- Coyne, C.J.; Porter, L.D.; Boutet, G.; Ma, Y.; McGee, R.J.; Lesné, A.; Baranger, A.; Pilet-Nayel, M.L. Confirmation of Fusarium root rot resistance QTL Fsp-Ps 2.1 of pea under controlled conditions. BMC Plant Biol. 2019, 19, 98. [Google Scholar] [CrossRef] [Green Version]
- Jha, U.C.; Bohra, A.; Pandey, S.; Parida, S.K. Breeding, genetics, and genomics approaches for improving Fusarium wilt resistance in major grain legumes. Front. Genet. 2020, 11, 1001. [Google Scholar] [CrossRef] [PubMed]
- Rispail, N.; Bani, M.; Rubiales, D. Resistance reaction of Medicago truncatula genotypes to Fusarium oxysporum: Effect of plant age, substrate and inoculation method. Crop Pasture Sci. 2015, 66, 506–515. [Google Scholar] [CrossRef]
- Sampaio, A.M.; Vitale, S.; Turrà, D.; Di Pietro, A.; Rubiales, D.; van Eeuwijk, F.; Vaz Patto, M.C. A diversity of resistance sources to Fusarium oxysporum f. sp. pisi found within grass pea germplasm. Plant Soil. 2021, 463, 19–38. [Google Scholar] [CrossRef]
- Leslie, J.F.; Summerell, B.A. Fusarium laboratory workshops--A recent history. Mycotoxin Res. 2006, 22, 73. [Google Scholar] [CrossRef]
- Wang, R.; Huang, J.; Liang, A.; Wang, Y.; Mur, L.A.J.; Wang, M.; Guo, S. Zinc and copper enhance cucumber tolerance to fusaric acid by mediating its distribution and toxicity and modifying the antioxidant system. Int. J. Mol. Sci. 2020, 21, 3370. [Google Scholar] [CrossRef]
- Wang, M.; Liu, W.; Yan, J.; Sun, P.; Chen, F.; Jiang, B.; Xie, D.; Lin, Y.; Peng, Q.; He, X. A Transcriptomic Analysis of Gene Expression in Chieh-Qua in Response to Fusaric Acid Stress. Horticulturae 2021, 7, 88. [Google Scholar] [CrossRef]
- Kraft, J.M.; Papavizas, G.C. Use of host resistance, Trichoderma, and fungicides to control soilborne diseases and increase seed yields of peas. Plant Dis. 1983, 11, 1234–1237. [Google Scholar] [CrossRef]
- Riaz, R.; Khan, A.; Khan, W.J.; Jabeen, Z.; Yasmin, H.; Naz, R.; Nosheen, A.; Hassan, M.N. Vegetable associated Bacillus spp. suppress the pea (Pisum sativum L.) root rot caused by Fusarium solani. Biol. Control 2021, 158, 104610. [Google Scholar] [CrossRef]
- Summerell, B.A.; Leslie, J.F. Fifty years of Fusarium: How could nine species have ever been enough? Fungal Divers. 2011, 50, 135–144. [Google Scholar] [CrossRef]
- Saremi, H.; Okhovvat, S.M.; Ashrafi, S.J. Fusarium diseases as the main soil borne fungal pathogen on plants and their control management with soil solarization in Iran. Afr. J. Biotechnol. 2011, 10, 18391–18398. [Google Scholar] [CrossRef]
- Dita, M.; Barquero, M.; Heck, D.; Mizubuti, E.S.G.; Staver, C.P. Fusarium wilt of banana: Current knowledge on epidemiology and research needs toward sustainable disease management. Front. Plant Sci. 2018, 871, 1468. [Google Scholar] [CrossRef] [Green Version]
- Grajal-Martìn, M.J.; Muehlbauer, F.J. Genomic location of the Fw gene for resistance to Fusarium wilt race 1 in peas. J. Hered. 2002, 93, 291–293. [Google Scholar] [CrossRef]
- McClendon, M.T.; Inglis, D.A.; McPhee, K.E.; Coyne, C.J. DNA markers linked to fusarium wilt race 1 resistance in pea. J. Am. Soc. Hortic. Sci. 2002, 127, 602–607. [Google Scholar] [CrossRef] [Green Version]
- Okubara, P.A.; Keller, K.E.; McClendon, M.T.; Inglis, D.A.; McPhee, K.E.; Coyne, C.J. Y15_999 Fw, a dominant SCAR marker linked to the Fusarium wilt race 1 (Fw) resistance gene in pea. Pisum Genet. 2015, 37, 30–33. [Google Scholar]
- Wohor, Z.O.; Rispail, N.; Rubiales, D. Evaluation of a Pisum spp. Germplasm Collection for the Resistance to Fusarium oxysporum f. sp. pisi Schlecht and Orobanche crenata Forsk. In Proceedings of the 9th International Conference on Legume Genetics and Genomics ICLGG, Dijon, France, 13 May 2019; Volume 71. [Google Scholar]
- McPhee, K.E.; Inglis, D.A.; Gundersen, B.; Coyne, C.J. Mapping QTL for Fusarium wilt Race 2 partial resistance in pea (Pisum sativum). Plant Breed. 2012, 131, 300–306. [Google Scholar] [CrossRef]
- Cheng, P.; Holdsworth, W.; Ma, Y.; Coyne, C.J.; Mazourek, M.; Grusak, M.A.; Fuchs, S.; McGee, R.J. Association mapping of agronomic and quality traits in USDA pea single-plant collection. Mol. Breed. 2015, 35, 1–13. [Google Scholar] [CrossRef]
- Sampaio, A.M.; Alves, M.L.; Pereira, P.; Valiollahi, E.; Santos, C.; Šatović, Z.; Rubiales, D.; Araújo, S.D.S.; van Eeuwijk, F.; Vaz Patto, M.C. Grass pea natural variation reveals oligogenic resistance to Fusarium oxysporum f. sp. pisi. Plant Genome 2021, 14, e20154. [Google Scholar] [CrossRef]
- Xue, A.G. Biological control of pathogens causing root rot complex in field pea using Clonostachys rosea strain ACM941. Phytopathology 2003, 93, 329–335. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.N. An Overview of Ecological and Habitat Aspects in the Genus Fusarium with Special Emphasis on the Soil- Borne Pathogenic Forms. Plant Pathol. 2007, 16, 97–120. [Google Scholar]
- Zitnick-Anderson, K.; Simons, K.; Pasche, J.S. Detection and qPCR quantification of seven Fusarium species associated with the root rot complex in field pea. Can. J. Plant Pathol. 2018, 40, 261–271. [Google Scholar] [CrossRef]
- Tu, J.C. Management of root rot diseases of peas, beans, and tomatoes. Can. J. Plant Pathol. 1992, 14, 92–99. [Google Scholar] [CrossRef]
- Sivachandra Kumar, N.T.; Cox, L.; Armstrong-Cho, C.; Banniza, S. Optimization of zoospore production and inoculum concentration of Aphanomyces euteiches for resistance screening of pea and lentil. Can. J. Plant Pathol. 2020, 42, 419–428. [Google Scholar] [CrossRef]
- Hossain, S.; Bergkvist, G.; Berglund, K.; Mårtensson, A.; Persson, P. Aphanomyces pea root rot disease and control with special reference to impact of Brassicaceae cover crops. Acta Agric. Scand. Sect. B Soil Plant Sci. 2012, 62, 477–487. [Google Scholar] [CrossRef]
- Wu, L.; Chang, K.F.; Conner, R.L.; Strelkov, S.; Fredua-Agyeman, R.; Hwang, S.F.; Feindel, D. Aphanomyces euteiches: A Threat to Canadian Field Pea Production. Engineering 2018, 4, 542–551. [Google Scholar] [CrossRef]
- Billard, E.; Quillévéré-Hamard, A.; Lavaud, C.; Pilet-Nayel, M.L.; Le May, C. Testing of life history traits of a soilborne pathogen in vitro: Do characteristics of oospores change according the strains of Aphanomyces euteiches and the host plant infected by the pathogen? J. Phytopathol. 2019, 167, 313–320. [Google Scholar] [CrossRef] [Green Version]
- Gaulin, E.; Madoui, M.A.; Bottin, A.; Jacquet, C.; Mathé, C.; Couloux, A.; Wincker, P.; Dumas, B. Transcriptome of Aphanomyces euteiches: New Oomycete putative pathogenicity factors and metabolic pathways. PLoS ONE 2008, 3, e1723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilet-Nayel, M.L.; Muehlbauer, F.J.; McGee, R.J.; Kraft, J.M.; Baranger, A.; Coyne, C.J. Consistent quantitative trait loci in pea for partial resistance to Aphanomyces euteiches isolates from the United States and France. Phytopathology 2005, 95, 1287–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, A.; Rani, M.; Lata, H.; Thakur, A.; Sharma, P.; Kumar, P.; Jayswal, D.K.; Rana, R.S. Global dimension of root rot complex in garden pea: Current status and breeding prospective. Crop Prot. 2022, 158, 106004. [Google Scholar] [CrossRef]
- Godebo, A.T.; Germida, J.J.; Walley, F.L. Isolation, identification, and assessment of soil bacteria as biocontrol agents of pea root rot caused by Aphanomyces euteiches. Can. J. Soil Sci. 2020, 100, 206–216. [Google Scholar] [CrossRef] [Green Version]
- Godebo, A.T.; Wee, N.M.J.; Yost, C.K.; Walley, F.L.; Germida, J.J. A Meta-Analysis to Determine the State of Biological Control of Aphanomyces Root Rot. Front. Mol. Biosci. 2021, 8, 777042. [Google Scholar] [CrossRef]
- Lagerlöf, J.; Ayuke, F.; Heyman, F.; Meijer, J. Effects of biocontrol bacteria and earthworms on Aphanomyces euteiches root-rot and growth of peas (Pisum sativum) studied in a pot experiment. Acta Agric. Scand. Sect. B–Soil Plant Sci. 2020, 70, 427–436. [Google Scholar] [CrossRef]
- Hossain, S.; Bergkvist, G.; Glinwood, R.; Berglund, K.; Mårtensson, A.; Hallin, S.; Persson, P. Brassicaceae cover crops reduce Aphanomyces pea root rot without suppressing genetic potential of microbial nitrogen cycling. Plant Soil 2015, 392, 227–238. [Google Scholar] [CrossRef] [Green Version]
- McGee, R.J.; Coyne, C.J.; Pilet-Nayel, M.L.; Moussart, A.; Tivoli, B.; Baranger, A.; Hamon, C.; Vandemark, G.; McPhee, K. Registration of pea germplasm lines partially resistant to Aphanomyces root rot for breeding fresh or freezer pea and dry pea types. J. Plant Regist. 2012, 6, 203–207. [Google Scholar] [CrossRef]
- Pilet-Nayel, M.L.; Muehlbauer, F.J.; McGee, R.J.; Kraft, J.M.; Baranger, A.; Coyne, C.J. Quantitative trait loci for partial resistance to Aphanomyces root rot in pea. Theor. Appl. Genet. 2002, 106, 28–39. [Google Scholar] [CrossRef]
- Hamon, C.; Coyne, C.J.; McGee, R.J.; Lesné, A.; Esnault, R.; Mangin, P.; Hervé, M.; Le Goff, I.; Deniot, G.; Roux-Duparque, M.; et al. QTL meta-analysis provides a comprehensive view of loci controlling partial resistance to Aphanomyces euteiches in four sources of resistance in pea. BMC Plant Biol. 2013, 13, 45. [Google Scholar] [CrossRef] [Green Version]
- Lavaud, C.; Lesné, A.; Piriou, C.; Le Roy, G.; Boutet, G.; Moussart, A.; Poncet, C.; Delourme, R.; Baranger, A.; Pilet-Nayel, M.L. Validation of QTL for resistance to Aphanomyces euteiches in different pea genetic backgrounds using near-isogenic lines. Theor. Appl. Genet. 2015, 128, 2273–2288. [Google Scholar] [CrossRef]
- Desgroux, A.; L’Anthoëne, V.; Roux-Duparque, M.; Rivière, J.P.; Aubert, G.; Tayeh, N.; Moussart, A.; Mangin, P.; Vetel, P.; Piriou, C.; et al. Genome-wide association mapping of partial resistance to Aphanomyces euteiches in pea. BMC Genom. 2016, 17, 124. [Google Scholar] [CrossRef] [Green Version]
- Desgroux, A.; Baudais, V.N.; Aubert, V.; Le Roy, G.; de Larambergue, H.; Miteul, H.; Aubert, G.; Boutet, G.; Duc, G.; Baranger, A.; et al. Comparative genome-wide-association mapping identifies common loci controlling root system architecture and resistance to Aphanomyces euteiches in pea. Front. Plant Sci. 2018, 8, 2195. [Google Scholar] [CrossRef] [Green Version]
- Bonhomme, M.; Fariello, M.I.; Navier, H.; Hajri, A.; Badis, Y.; Miteul, H.; Samac, D.A.; Dumas, B.; Baranger, A.; Jacquet, C.; et al. A local score approach improves GWAS resolution and detects minor QTL: Application to Medicago truncatula quantitative disease resistance to multiple Aphanomyces euteiches isolates. Heredity 2019, 123, 517–531. [Google Scholar] [CrossRef]
- Wu, L.; Fredua-Agyeman, R.; Hwang, S.F.; Chang, K.F.; Conner, R.L.; McLaren, D.L.; Strelkov, S.E. Mapping QTL associated with partial resistance to Aphanomyces root rot in pea (Pisum sativum L.) using a 13.2 K SNP array and SSR markers. Theor. Appl. Genet. 2021, 134, 2965–2990. [Google Scholar] [CrossRef]
- Hadwiger, L.A. Pea-Fusarium solani interactions contributions of a system toward understanding disease resistance. Phytopathology 2008, 98, 372–379. [Google Scholar] [CrossRef] [Green Version]
- Baćanović-Šišić, J.; Šišić, A.; Schmidt, J.H.; Finckh, M.R. Identification and characterization of pathogens associated with root rot of winter peas grown under organic management in Germany. Eur. J. Plant Pathol. 2018, 151, 745–755. [Google Scholar] [CrossRef]
- Fernandez, M.R.; Huber, D.; Basnyat, P.; Zentner, R.P. Impact of agronomic practices on populations of Fusarium and other fungi in cereal and noncereal crop residues on the Canadian Prairies. Soil Tillage Res. 2008, 100, 60–71. [Google Scholar] [CrossRef]
- Bodah, E.T.; Porter, L.D.; Chaves, B.; Dhingra, A. Evaluation of pea accessions and commercial cultivars for fusarium root rot resistance. Euphytica 2016, 208, 63–72. [Google Scholar] [CrossRef]
- Singh, B.P.; Singh, G.; Krishna, K.; Nayak, S.C.; Srinivasa, N. Management of Fungal Pathogens in Pulses: Current Status and Future Challenges; Springer Nature: Cham, Switzerland, 2020; Volume 11224, pp. 35947–35948. [Google Scholar] [CrossRef]
- Mazzola, M. Mechanisms of natural soil suppressiveness to soilborne diseases. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2002, 81, 557–564. [Google Scholar] [CrossRef]
- Jha, P.K.; Jalali, B.L. Biocontrol of pea root rot incited by Fusarium solani f. sp. pisi with rhizosphere mycoflora. Indian Phytopathol. 2006, 59, 41–43. [Google Scholar]
- Panth, M.; Hassler, S.C.; Baysal-Gurel, F. Methods for Management of Soilborne Diseases in Crop Production. Agriculture 2020, 10, 16. [Google Scholar] [CrossRef] [Green Version]
- Grünwald, N.J.; Coffman, V.A.; Kraft, J.M. Sources of Partial Resistance to Fusarium Root Rot in the Pisum Core Collection. Plant Dis. 2003, 87, 1197–1200. [Google Scholar] [CrossRef] [Green Version]
- Li, W.J.; Feng, J.; Chang, K.F.; Conner, R.L.; Hwang, S.F.; Strelkov, S.E.; Gossen, B.D.; McLaren, D.L. Microsatellite DNA markers indicate quantitative trait loci controlling resistance to pea root rot caused by Fusarium avenaceum (Corda ex Fries) Sacc. Plant Pathol. J. 2012, 11, 114–119. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Hwang, R.; Chang, K.F.; Conner, R.L.; Hwang, S.F.; Strelkov, S.E.; Gossen, B.D.; McLaren, D.L.; Xue, A.G. Identification of microsatellite markers linked to quantitative trait loci controlling resistance to Fusarium root rot in field pea. Can. J. Plant Sci. 2011, 91, 199–204. [Google Scholar] [CrossRef]
- Williamson-Benavides, B.A.; Sharpe, R.M.; Nelson, G.; Bodah, E.T.; Porter, L.D.; Dhingra, A. Identification of Fusarium solani f. sp. pisi (Fsp) Responsive Genes in Pisum sativum. Front. Genet. 2020, 11, 950. [Google Scholar] [CrossRef]
- Grünwald, N.J.; Chen, W.; Larsen, R.C. Pea Diseases and their Management. In Diseases of Fruits and Vegetables; Springer: Dordrech, The Netherlands, 2004; Volume 2, pp. 301–331. [Google Scholar] [CrossRef]
- Nel, W.J.; Duong, T.A.; Wingfield, B.D.; Wingfield, M.J.; de Beer, Z.W. A new genus and species for the globally important, multihost root pathogen Thielaviopsis basicola. Plant Pathol. 2018, 67, 871–882. [Google Scholar] [CrossRef]
- Abawi, G.S.; Widmer, T.L. Impact of soil health management practices on soilborne pathogens, nematodes and root diseases of vegetable crops. Appl. Soil Ecol. 2000, 15, 37–47. [Google Scholar] [CrossRef]
- Hood, M.E.; Shew, H.D. Pathogenesis of Thielaviopsis basicola on a susceptible and a resistant cultivar of burley tobacco. Phytopathology 1996, 86, 38–44. [Google Scholar] [CrossRef]
- Niu, C.; Lister, H.E.; Nguyen, B.; Wheeler, T.A.; Wright, R.J. Resistance to Thielaviopsis basicola in the cultivated a genome cotton. Theor. Appl. Genet. 2008, 117, 1313–1323. [Google Scholar] [CrossRef] [PubMed]
- Keijer, J. The initial steps of the infection process in Rhizoctonia solani. In Rhizoctonia Species: Taxonomy, Molecular Biology, Ecology, Pathology and Disease Control; Springer: Dordrecht, The Netherlands, 1996; pp. 149–162. [Google Scholar]
- Sharma-Poudyal, D.; Paulitz, T.C.; Porter, L.D.; Sharma-Poudyal, D. Characterization and pathogenicity of Rhizoctonia and Rhizoctonia-like spp. From pea crops in the Columbia Basin of Oregon and Washington. Plant Dis. 2015, 99, 604–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beniwal, S.P.S.; Ahmed, S.; Gorfu, D. Wilt/root rot diseases of chickpea in Ethiopia. Trop. Pest Manag. 1992, 38, 48–51. [Google Scholar] [CrossRef]
- Ketta, H.; Elkhateeb, N.; Saleh, M.; Kamel, S. Efficiency Assessment of Combinations Between Rhizobium leguminosarum and Trichoderma spp. for Controlling of Pea (Pisum sativum L.) Damping-off Disease. Egypt. J. Phytopathol. 2021, 49, 1–14. [Google Scholar] [CrossRef]
- Uwaremwe, C.; Yue, L.; Liu, Y.; Tian, Y.; Zhao, X.; Wang, Y.; Xie, Z.; Zhang, Y.; Cui, Z.; Wang, R. Molecular identification and pathogenicity of Fusarium and Alternaria species associated with root rot disease of wolfberry in Gansu and Ningxia provinces, China. Plant Pathol. 2021, 70, 397–406. [Google Scholar] [CrossRef]
- Wille, L.; Kurmann, M.; Messmer, M.M.; Studer, B.; Hohmann, P. Untangling the Pea Root Rot Complex Reveals Microbial Markers for Plant Health. Front. Plant Sci. 2021, 12, 737820. [Google Scholar] [CrossRef]
- Flower, K.C.; Hüberli, D.; Collins, S.J.; Thomas, G.; Ward, P.R.; Cordingley, N. Progression of plant-parasitic nematodes and foliar and root diseases under no-tillage with different crop rotations. Soil Tillage Res. 2019, 191, 18–28. [Google Scholar] [CrossRef]
- El_Komy, M.H.; Hassouna, M.G.; Abou-Taleb, E.M.; Al-Sarar, A.S.; Abobakr, Y. A mixture of Azotobacter, Azospirillum, and Klebsiella strains improves root-rot disease complex management and promotes growth in sunflowers in calcareous soil. Eur. J. Plant Pathol. 2020, 156, 713–726. [Google Scholar] [CrossRef]
- Khan, M.R.; Ashraf, S.; Rasool, F.; Salati, K.M.; Mohiddin, F.A.; Haque, Z. Field performance of Trichoderma species against wilt disease complex of chickpea caused by Fusarium oxysporum f. sp. ciceri and Rhizoctonia solani. Turk. J. Agric. For. 2014, 38, 447–454. [Google Scholar] [CrossRef]
- Hashem, A.H.; Abdelaziz, A.M.; Askar, A.A.; Fouda, H.M.; Khalil, A.M.A.; Abd-Elsalam, K.A.; Khaleil, M.M. Bacillus megaterium-mediated synthesis of selenium nanoparticles and their antifungal activity against Rhizoctonia solani in faba bean plants. J. Fungi 2021, 7, 195. [Google Scholar] [CrossRef]
- Cubeta, M.A.; Thomas, E.; Dean, R.A.; Jabaji, S.; Neate, S.M.; Tavantzis, S.; Toda, T.; Vilgalys, R.; Bharathan, N.; Fedorova-Abrams, N.; et al. Draft genome sequence of the plant-pathogenic soil fungus Rhizoctonia solani anastomosis group 3 strain Rhs1AP. Genome Announc. 2014, 2, e01072-14. [Google Scholar] [CrossRef] [Green Version]
- Zhong, J.; Chen, C.Y.; Gao, B.D. Genome sequence of a novel mycovirus of Rhizoctonia solani, a plant pathogenic fungus. Virus Genes 2015, 51, 167–170. [Google Scholar] [CrossRef]
- Chen, Y.; Su, J.E.; Qin, X.Y.; Fan, Z.Y.; Zhang, X.H.; Yu, Q.; Xia, Z.Y.; Zou, C.M.; Zhao, G.K.; Lin, Z.L. A novel putative betapartitivirus isolated from the plant-pathogenic fungus Rhizoctonia solani. Arch. Virol. 2020, 165, 1697–1701. [Google Scholar] [CrossRef]
- Akhter, W.; Bhuiyan, M.K.A.; Sultana, F.; Hossain, M.M. Integrated effect of microbial antagonist, organic amendment and fungicide in controlling seedling mortality (Rhizoctonia solani) and improving yield in pea (Pisum sativum L.). Comptes Rendus Biol. 2015, 338, 21–28. [Google Scholar] [CrossRef]
- Wang, Y.; Fristensky, B. Transgenic canola lines expressing pea defense gene DRR206 have resistance to aggressive blackleg isolates and to Rhizoctonia solani. Mol. Breed. 2001, 8, 263–271. [Google Scholar] [CrossRef]
- Dölfors, F.; Holmquist, L.; Dixelius, C.; Tzelepis, G. A LysM effector protein from the basidiomycete Rhizoctonia solani contributes to virulence through suppression of chitin-triggered immunity. Mol. Genet. Genom. 2019, 294, 1211–1218. [Google Scholar] [CrossRef]
- Liu, Y.; Hassan, S.; Kidd, B.N.; Garg, G.; Mathesius, U.; Singh, K.B.; Anderson, J.P. Ethylene signaling is important for isoflavonoid-mediated resistance to Rhizoctonia solani in roots of Medicago truncatula. Mol. Plant-Microbe Interact. 2017, 30, 691–700. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, K.L.; Martin, F.N.; de Cock, A.W.A.M.; Lévesque, C.A.; Spies, C.F.J.; Okubara, P.A.; Paulitz, T.C. Molecular detection and quantification of pythium species: Evolving taxonomy, new tools, and challenges. Plant Dis. 2013, 97, 4–20. [Google Scholar] [CrossRef] [Green Version]
- Kageyama, K. Molecular taxonomy and its application to ecological studies of Pythium species. J. Gen. Plant Pathol. 2014, 80, 314–326. [Google Scholar] [CrossRef]
- Khalil, S.A.M.; Nehal, S.-M.; Nadia, G.-G.; Mokhtar, M.-K. Field approaches of chemical inducers and bioagents for controlling root diseases incidence of pea (Pisum sativum L.) under field conditions. Plant Pathol. J. 2020, 19, 166–175. [Google Scholar] [CrossRef]
- Wu, W.; Ogawa, F.; Ochiai, M.; Yamada, K.; Fukui, H. Common strategies to control pythium disease. Rev. Agric. Sci. 2020, 8, 58–69. [Google Scholar] [CrossRef]
- Kraft, J.M.; Haware, M.P.; Jiménez-Díaz, R.M.; Bayaa, B.; Harrabi, M. Screening techniques and sources of resistance to root rots and wilts in cool season food legumes. Euphytica 1993, 73, 27–39. [Google Scholar] [CrossRef] [Green Version]
- Alcala, A.V.C.; Paulitz, T.C.; Schroeder, K.L.; Porter, L.D.; Derie, M.L.; du Toit, L.J. Pythium species associated with damping-off of pea in certified organic fields in the Columbia Basin of central Washington. Plant Dis. 2016, 100, 916–925. [Google Scholar] [CrossRef] [Green Version]
- Klepadlo, M.; Balk, C.S.; Vuong, T.D.; Dorrance, A.E.; Nguyen, H.T. Molecular characterization of genomic regions for resistance to Pythium ultimum var. ultimum in the soybean cultivar Magellan. Theor. Appl. Genet. 2019, 132, 405–417. [Google Scholar] [CrossRef]
- Urrea, K.; Rupe, J.; Chen, P.; Rothrock, C.S. Characterization of seed rot resistance to Pythium aphanidermatum in soybean. Crop Sci. 2017, 57, 1394–1403. [Google Scholar] [CrossRef]
- Lin, F.; Wani, S.H.; Collins, P.J.; Wen, Z.; Li, W.; Zhang, N.; McCoy, A.G.; Bi, Y.; Tan, R.; Zhang, S.; et al. QTL mapping and GWAS for identification of loci conferring partial resistance to Pythium sylvaticum in soybean (Glycine max (L.) Merr). Mol. Breed. 2020, 40, 1–11. [Google Scholar] [CrossRef]
- Navarro, F.; Sass, M.E.; Nienhuis, J. Identification and confirmation of quantitative trait loci for root rot resistance in snap bean. Crop Sci. 2008, 48, 962–972. [Google Scholar] [CrossRef]
- Arora, H.; Sharma, A.; Sharma, S.; Haron, F.F.; Gafur, A.; Sayyed, R.Z.; Datta, R. Pythium damping-off and root rot of Capsicum annuum L.: Impacts, diagnosis, and management. Microorganisms 2021, 9, 823. [Google Scholar] [CrossRef]
- Ai, G.; Yang, K.; Ye, W.; Tian, Y.; Du, Y.; Zhu, H.; Li, T.; Xia, Q.; Shen, D.; Peng, H.; et al. Prediction and characterization of RXLR effectors in Pythium species. Mol. Plant-Microbe Interact. 2020, 33, 1046–1058. [Google Scholar] [CrossRef]
- Trudgill, D.L.; Blok, V.C. Apomictic, Polyphagous Root-Knot Nematodes: Exceptionally Successful and Damaging Biotrophic Root Pathogens. Annu. Rev. Phytopathol. 2001, 39, 53–77. [Google Scholar] [CrossRef] [PubMed]
- Mesa-Valle, C.M.; Garrido-Cardenas, J.A.; Cebrian-Carmona, J.; Talavera, M.; Manzano-Agugliaro, F. Global research on plant nematodes. Agronomy 2020, 10, 1148. [Google Scholar] [CrossRef]
- Castillo, P.; Navas-Cortés, J.A.; Landa, B.B.; Jiménez-Díaz, R.M.; Vovlas, N. Plant-parasitic nematodes attacking chickpea and their in planta interactions with rhizobia and phytopathogenic fungi. Plant Dis. 2008, 92, 840–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, J.T.; Haegeman, A.; Danchin, E.G.J.; Gaur, H.S.; Helder, J.; Jones, M.G.K.; Kikuchi, T.; Manzanilla-López, R.; Palomares-Rius, J.E.; Wesemael, W.M.L.; et al. Top 10 plant-parasitic nematodes in molecular plant pathology. Mol. Plant Pathol. 2013, 14, 946–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobosz, R.; Krawczyk, R. Meloidogyne hapla development on growing legume plants–Short Communication. Plant Prot. Sci. 2019, 55, 274–277. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.K.; Hodda, M.; Ash, G.J. Plant-parasitic nematodes of potential phytosanitary importance, their main hosts and reported yield losses. EPPO Bull. 2013, 43, 334–374. [Google Scholar] [CrossRef]
- Kimpinski, J.; Sturz, A.V. Managing crop root zone ecosystems for prevention of harmful and encouragement of beneficial nematodes. Soil Tillage Res. 2003, 72, 213–221. [Google Scholar] [CrossRef]
- Zhang, S.; Cui, S.; McLaughlin, N.B.; Liu, P.; Hu, N.; Liang, W.; Wu, D.; Liang, A. Tillage effects outweigh seasonal effects on soil nematode community structure. Soil Tillage Res. 2019, 192, 233–239. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, S.; Li, H.; Wang, R.; Zhang, K.Q.; Xu, J. Fungi–nematode interactions: Diversity, ecology, and biocontrol prospects in agriculture. J. Fungi 2020, 6, 206. [Google Scholar] [CrossRef]
- Li, X.; Liu, C.; Zhao, H.; Gao, F.; Ji, G.; Hu, F.; Li, H. Similar positive effects of beneficial bacteria, nematodes and earthworms on soil quality and productivity. Appl. Soil Ecol. 2018, 130, 202–208. [Google Scholar] [CrossRef]
- Di Vito, M.; Greco, N. Control of food legume nematodes in the Mediterranean Basin 1. EPPO Bull. 1994, 24, 489–494. [Google Scholar] [CrossRef]
- Sillero, J.C.; Villegas-Fernández, A.M.; Thomas, J.; Rojas-Molina, M.M.; Emeran, A.A.; Fernández-Aparicio, M.; Rubiales, D. Faba bean breeding for disease resistance. Field Crops Res. 2010, 115, 297–307. [Google Scholar] [CrossRef] [Green Version]
- Kosterin, O.E. Prospects of the use of wild relatives for pea breeding. Russ. J. Genet. Appl. Res. 2016, 6, 233–243. [Google Scholar] [CrossRef]
- Zwart, R.S.; Thudi, M.; Channale, S.; Manchikatla, P.K.; Varshney, R.K.; Thompson, J.P. Resistance to Plant-Parasitic Nematodes in Chickpea: Current Status and Future Perspectives. Front. Plant Sci. 2019, 10, 966. [Google Scholar] [CrossRef] [Green Version]
- Vovlas, A.; Santoro, S.; Radicci, V.; Leonetti, P.; Castillo, P.; Palomares-Rius, J.E. Host-suitability of black medick (Medicago lupulina L.) and additional molecular markers for identification of the pea cyst nematode Heterodera goettingiana. Eur. J. Plant Pathol. 2017, 149, 193–199. [Google Scholar] [CrossRef]
- Jones, M.G.K. Host cell responses to endoparasitic nematode attack: Structure and function of giant cells and syncytia. Ann. Appl. Biol. 1981, 97, 353–372. [Google Scholar] [CrossRef]
- Munawar, M.; Yevtushenko, D.P.; Castillo, P. Integrative taxonomy, distribution, and host associations of Geocenamus brevidens and Quinisulcius capitatus from southern Alberta, Canada. J. Nematol. 2021, 53, 1–17. [Google Scholar] [CrossRef]
- Whitehead, A.G.; Bromilow, R.H.; Tite, D.J.; Finch, P.H.; Fraser, J.E.; French, E.M. Incorporation of granular nematicides in soil to control pea cyst-nematode, Heterodera goettingiana. Ann. Appl. Biol. 1979, 92, 81–91. [Google Scholar] [CrossRef]
- Green, C.D.; Williamson, K.; Dennis, E.B.; McBurney, T. The effect of oxamyl on the growth of peas attacked by pea cyst nematode. Ann. Appl. Biol. 1981, 97, 303–309. [Google Scholar] [CrossRef]
- Dopierata, U.; Giebel, J. Herbicides can influence the level of pea infestation by Heterodera goettingiana. J. Plant Prot. 2002, 42, 337–341. [Google Scholar]
- Di Vito, M.; Perrino, P. Reaction of Pisum spp. to the attacks of Heterodera goettingiana. Nematol. Mediterr. 1978, 6, 113–118. [Google Scholar]
- Bleve-Zacheo, T.; Melillo, M.T.; Zacheo, G. Syncytia development in germplasm pea accessions infected with Heterodera goettingiana. Nematol Mediterr. 1990, 18, 93–102. [Google Scholar]
- Veronico, P.; Melillo, M.T.; Saponaro, C.; Leonetti, P.; Picardi, E.; Jones, J.T. A polygalacturonase-inhibiting protein with a role in pea defence against the cyst nematode Heterodera goettingiana. Mol. Plant Pathol. 2011, 12, 275–287. [Google Scholar] [CrossRef]
- Veronico, P.; Giannino, D.; Melillo, M.T.; Leone, A.; Reyes, A.; Kennedy, M.W.; Bleve-Zacheo, T. A novel lipoxygenase in pea roots. Its function in wounding and biotic stress. Plant Physiol. 2006, 141, 1045–1055. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.B.; Sikora, R.A.; Greco, N.; Di Vito, M.; Caubel, G. Screening techniques and sources of resistance to nematodes in cool season food legumes. Euphytica 1993, 73, 59–66. [Google Scholar] [CrossRef]
- Castillo, P.; Navas-Cortés, J.A.; Gomar-Tinoco, D.; Di Vito, M.; Jiménez-Díaz, R.M. Interactions between Meloidogyne artiellia, the Cereal and Legume Root-Knot Nematode, and Fusarium oxysporum f. sp. ciceris Race 5 in Chickpea. Phytopathology 2003, 93, 1513–1523. [Google Scholar] [CrossRef] [Green Version]
- Sturz, A.V.; Christie, B.R. Beneficial microbial allelopathies in the root zone: The management of soil quality and plant disease with rhizobacteria. Soil Tillage Res. 2003, 72, 107–123. [Google Scholar] [CrossRef]
- Haque, Z.; Khan, M.R. Identification of multi-facial microbial isolates from the rice rhizosphere and their biocontrol activity against Rhizoctonia solani AG1-IA. Biol. Control 2021, 161, 104640. [Google Scholar] [CrossRef]
- Pandey, R.; Kalra, A.; Tandon, S.; Mehrotra, N.; Singh, H.N.; Kumar, S. Essential oils as potent source of nematicidal compounds. J. Phytopathol. 2000, 148, 501–502. [Google Scholar] [CrossRef]
- Sharma, A.; Haseeb, A.; Abuzar, S. Screening of field pea (Pisum sativum) selections for their reactions to root-knot nematode (Meloidogyne incognita). J. Zhejiang Univ. Sci. B. 2006, 7, 209–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gautam, N.K.; Marla, S.S.; Mirza, N.; Khan, Z.; Singh, B.; Wankhede, D.P.; Gawade, B.H. Evaluation of field pea accessions for root-knot nematode resistance and possible role of NADP dependent malic enzyme gene in host resistance. Indian J. Genet. Plant Breed. 2017, 77, 4. [Google Scholar] [CrossRef]
- Youssef, M.; El-Nagdi, W. Differential responses of certain field pea and cowpea cultivars to root-knot nematode, Meloidogyne incognita for commercial release. Bull. Natl. Res. Cent. 2019, 43, 178. [Google Scholar] [CrossRef]
- Hewezi, T.; Baum, T.J. Manipulation of plant cells by cyst and root-knot nematode effectors. Mol. Plant-Microbe Interact. 2013, 26, 9–16. [Google Scholar] [CrossRef]
- Clément, M.; Ketelaar, T.; Rodiuc, N.; Banora, M.Y.; Smertenko, A.; Engler, G.; Abad, P.; Hussey, P.J.; de Almeida Engler, J. Actin-depolymerizing factor2-mediated actin dynamics are essential for root-knot nematode infection of Arabidopsis. Plant Cell 2009, 21, 2963–2979. [Google Scholar] [CrossRef] [Green Version]
- Abad, P.; Gouzy, J.; Aury, J.M.; Castagnone-Sereno, P.; Danchin, E.G.; Deleury, E.; Perfus-Barbeoch, L.; Anthouard, V.; Artiguenave, F.; Blok, V.C.; et al. Genome sequence of the metazoan plant-parasitic nematode Meloidogyne incognita. Nat. Biotechnol. 2008, 26, 909–915. [Google Scholar] [CrossRef] [Green Version]
- Upadhaya, A.; Yan, G.; Pasche, J. Reproduction Ability and Growth Effect of Pin Nematode, Paratylenchus nanus, With Selected Field Pea Cultivars. Plant Dis. 2019, 103, 2520–2526. [Google Scholar] [CrossRef]
- Reen, R.A.; Mumford, M.H.; Thompson, J.P. Novel Sources of Resistance to Root-Lesion Nematode (Pratylenchus thornei) in a New Collection of Wild Cicer Species (C. reticulatum and C. echinospermum) to Improve Resistance in Cultivated Chickpea (C. arietinum). Phytopathology 2019, 109, 1270–1279. [Google Scholar] [CrossRef]
- Kandel, S.L.; Smiley, R.W.; Garland-Campbell, K.; Elling, A.A.; Huggins, D.; Paulitz, T.C. Spatial distribution of root lesion nematodes (Pratylenchus spp.) in a long-term no-till cropping system and their relationship with soil and landscape properties. Eur. J. Plant Pathol. 2018, 150, 1011–1021. [Google Scholar] [CrossRef]
- Smiley, R. Root-lesion Nematodes Affecting Dryland Cereals in the Semiarid Pacific Northwest USA. Plant Dis. 2021, 105, 3324–3343. [Google Scholar] [CrossRef]
- Taylor, S.P.; Hollaway, G.J.; Hunt, C.H. Effect of field crops on population densities of Pratylenchus neglectus and P. thornei in Southeastern Australia; Part 1: P. neglectus. J. Nematol. 2000, 32, 591–599. [Google Scholar]
- Thompson, J.P. Treatments to eliminate root-lesion nematode (Pratylenchus thornei Sher & Allen) from a vertisol. Nematologica 1990, 36, 123–127. [Google Scholar]
- Taylor, S.P.; Vanstone, V.A.; Ware, A.H.; McKay, A.C.; Szot, D.; Russ, M.H. Measuring yield loss in cereals caused by root lesion nematodes (Pratylenchus neglectus and P. thornei) with and without nematicide. Aust. J. Agric. Res. 1999, 50, 617–627. [Google Scholar] [CrossRef]
- Thompson, J.P.; Reen, R.A.; Clewett, T.G.; Sheedy, J.G.; Kelly, A.M.; Gogel, B.J.; Knights, E.J. Hybridisation of Australian chickpea cultivars with wild Cicer spp. increases resistance to root-lesion nematodes (Pratylenchus thornei and P. neglectus). Australas. Plant Pathol. 2011, 40, 601–611. [Google Scholar] [CrossRef]
- Rubiales, D.; Fernández-Aparicio, M. Parasitic weed: Broomrape. In Compendium of Pea Diseases and Pests, 3rd ed.; Harveson, R.M., Ed.; American Phytopathological Society: St. Paul, MN, USA, 2021; pp. 67–69. ISBN 978-0-89054-655-0. [Google Scholar]
- Rubiales, D.; Fernández-Aparicio, M. Innovations in parasitic weeds management in legume crops. A review. Agron. Sustain. Dev. 2012, 32, 433–449. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Flores, F.; Rubiales, D. The effect of Orobanche crenata infection severity in faba bean, field pea, and grass pea productivity. Front. Plant Sci. 2016, 7, 1409. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Yoneyama, K.; Rubiales, D. The role of strigolactones in host specificity of Orobanche and Phelipanche seed germination. Seed Sci. Res. 2011, 21, 55–61. [Google Scholar] [CrossRef] [Green Version]
- Rubiales, D.; Moreno, M.T.; Sillero, J.C. Search for resistance to crenate broomrape (Orobanche crenata Forsk.) in pea germplasm. Genet. Resour. Crop Evol. 2005, 52, 853–861. [Google Scholar] [CrossRef]
- Rispail, N.; Dita, M.A.; González-Verdejo, C.; Pérez-de-Luque, A.; Castillejo, M.A.; Prats, E.; Román, B.; Jorrín, J.; Rubiales, D. Plant resistance to parasitic plants: Molecular approaches to an old foe. New Phytol. 2007, 173, 703–712. [Google Scholar] [CrossRef] [Green Version]
- Pérez-de-Luque, A.; Moreno, M.T.; Rubiales, D. Host plant resistance against broomrapes (Orobanche spp.): Defence reactions and mechanisms of resistance. Ann. Appl. Biol. 2008, 152, 131–141. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Rubiales, D. Advances in understanding plant root response to weedy root parasites. In Improving Crop Root Function, 1st ed.; Gregory, P., Ed.; Burleig Dodds Science Publishing Limited: Cambridge, UK, 2021; pp. 215–230. [Google Scholar] [CrossRef]
- Rubiales, D.; Fernández-Aparicio, M.; Wegmann, K.; Joel, D.M. Revisiting strategies for reducing the seedbank of Orobanche and Phelipanche spp. Weed Res. 2009, 49, 23–33. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Sillero, J.C.; Rubiales, D. Intercropping with cereals reduces infection by Orobanche crenata in legumes. Crop Prot. 2007, 26, 1166–1172. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Emeran, A.A.; Rubiales, D. Control of Orobanche crenata in legumes intercropped with fenugreek (Trigonella foenum-graecum). Crop Prot. 2008, 27, 653–659. [Google Scholar] [CrossRef]
- Rubiales, D. Parasitic plants, wild relatives and the nature of resistance. New Phytol. 2003, 160, 459–461. [Google Scholar] [CrossRef] [PubMed]
- Mabrouk, Y.; Zourgui, L.; Sifi, B.; Delavault, P.; Simier, P.; Belhadj, O. Some compatible Rhizobium leguminosarum strains in peas decrease infections when parasitised by Orobanche crenata. Weed Res. 2007, 47, 44–53. [Google Scholar] [CrossRef]
- Mabrouk, Y.; Mejri, S.; Belhadj, O. Biochemical mechanisms of induced resistance by rhizobial lipopolysaccharide in pea against crenate broomrape. Rev. Bras. Bot. 2016, 39, 107–114. [Google Scholar] [CrossRef]
- Louarn, J.; Carbonne, F.; Delavault, P.; Bécard, G.; Rochange, S. Reduced Germination of Orobanche cumana Seeds in the Presence of Arbuscular Mycorrhizal Fungi or Their Exudates. PLoS ONE 2012, 7, e49273. [Google Scholar] [CrossRef] [Green Version]
- Rubiales, D.; Fernández-Aparicio, M.; Pérez-de-Luque, A.; Castillejo, M.A.; Prats, E.; Sillero, J.C.; Rispail, N.; Fondevilla, S. Breeding approaches for crenate broomrape (Orobanche crenata Forsk.) management in pea (Pisum sativum L.). Pest Manag. Sci. 2009, 65, 553–559. [Google Scholar] [CrossRef]
- Pérez-de-Luque, A.; Jorrín, J.; Cubero, J.I.; Rubiales, D. Orobanche crenata resistance and avoidance in pea (Pisum spp.) operate at different developmental stages of the parasite. Weed Res. 2005, 45, 379–387. [Google Scholar] [CrossRef]
- Rubiales, D. Broomrape threat to agriculture. Outlooks Pest Manag. 2020, 31, 141–144. [Google Scholar] [CrossRef]
- Valderrama, M.R.; Román, B.; Satovic, Z.; Rubiales, D.; Cubero, J.I.; Torres, A.M. Locating quantitative trait loci associated with Orobanche crenata resistance in pea. Weed Res. 2004, 44, 323–328. [Google Scholar] [CrossRef]
- Fondevilla, S.; Fernández-Aparicio, M.; Satovic, Z.; Emeran, A.A.; Torres, A.M.; Moreno, M.T.; Rubiales, D. Identification of quantitative trait loci for specific mechanisms of resistance to Orobanche crenata Forsk. in pea (Pisum sativum L.). Mol. Breed. 2010, 25, 259–272. [Google Scholar] [CrossRef] [Green Version]
- Fondevilla, S.; Flores, F.; Emeran, A.A.; Kharrat, M.; Rubiales, D. High productivity of dry pea genotypes resistant to crenate broomrape in Mediterranean environments. Agron. Sustain. Dev. 2017, 37, 61. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Aparicio, M.; Flores, F.; Rubiales, D. Recognition of root exudates by seeds of broomrape (Orobanche and Phelipanche) species. Ann. Bot. 2009, 103, 423–431. [Google Scholar] [CrossRef] [Green Version]
- Dita, M.A.; Die, J.V.; Román, B.; Krajinski, F.; Küster, H.; Moreno, M.T.; Cubero, J.I.; Rubiales, D. Gene expression profiling of Medicago truncatula roots in response to the parasitic plant Orobanche crenata. Weed Res. 2009, 49, 66–80. [Google Scholar] [CrossRef]
- Castillejo, M.Á.; Maldonado, A.M.; Dumas-Gaudot, E.; Fernández-Aparicio, M.; Susín, R.; Rubiales, D.; Jorrín, J.V. Differential expression proteomics to investigate responses and resistance to Orobanche crenata in Medicago truncatula. BMC Genom. 2009, 10, 294. [Google Scholar] [CrossRef]
- Castillejo, M.Á.; Amiour, N.; Dumas-Gaudot, E.; Rubiales, D.; Jorrín, J.V. A proteomic approach to studying plant response to crenate broomrape (Orobanche crenata) in pea (Pisum sativum). Phytochemistry 2004, 65, 1817–1828. [Google Scholar] [CrossRef]
- Rubiales, D. Can we breed for durable resistance to broomrapes? Phytopathol. Mediterr. 2018, 57, 170–185. [Google Scholar]
- Varshney, R.K.; Kudapa, H.; Pazhamala, L.; Chitikineni, A.; Thudi, M.; Bohra, A.; Gaur, P.M.; Janila, P.; Fikre, A.; Kimurto, P.; et al. Translational Genomics in Agriculture: Some Examples in Grain Legumes. CRC. Crit. Rev. Plant Sci. 2015, 34, 169–194. [Google Scholar] [CrossRef] [Green Version]
- Thudi, M.; Palakurthi, R.; Schnable, J.C.; Chitikineni, A.; Dreisigacker, S.; Mace, E.; Srivastava, R.K.; Satyavathi, C.T.; Odeny, D.; Tiwari, V.K.; et al. Genomic resources in plant breeding for sustainable agriculture. J. Plant Physiol. 2021, 257, 153351. [Google Scholar] [CrossRef]
- Tivoli, B.; Baranger, A.; Avila, C.M.; Banniza, S.; Barbetti, M.; Chen, W.; Davidson, J.; Lindeck, K.; Kharrat, M.; Rubiales, D.; et al. Screening techniques and sources of resistance to foliar diseases caused by major necrotrophic fungi in grain legumes. Euphytica 2006, 147, 223–253. [Google Scholar] [CrossRef]
- Burud, I.; Lange, G.; Lillemo, M.; Bleken, E.; Grimstad, L.; From, P.J. Exploring robots and UAVs as phenotyping tools in plant breeding. IFAC-PapersOnLine 2017, 50, 11479–11484. [Google Scholar] [CrossRef]
- Chen, C.Y.; Butts, C.L.; Dang, P.M.; Wang, M.L. Advances in phenotyping of functional traits. In Phenomics in Crop Plants: Trends, options and Limitations; Springer: New Delhi, India, 2015; pp. 163–180. [Google Scholar]
- Cazzola, F.; Bermejo, C.J.; Guindon, M.F.; Cointry, E. Speed breeding in pea (Pisum sativum L.), an efficient and simple system to accelerate breeding programs. Euphytica 2020, 216, 178. [Google Scholar] [CrossRef]
- Rubiales, D.; Pérez-de-Luque, A.; Fernández-Aparico, M.; Sillero, J.C.; Román, B.; Kharrat, M.; Khalil, S.; Joel, D.M.; Riches, C. Screening techniques and sources of resistance against parasitic weeds in grain legumes. Euphytica 2006, 147, 187–199. [Google Scholar] [CrossRef]
- Furbank, R.T.; Tester, M. Phenomics–technologies to relieve the phenotyping bottleneck. Trends Plant Sci. 2011, 16, 635–644. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Rubiales, D. Characterisation of resistance to crenate broomrape (Orobanche crenata Forsk.) in Lathyrus cicera L. Euphytica 2010, 173, 77–84. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Moral, A.; Kharrat, M.; Rubiales, D. Resistance against broomrapes (Orobanche and Phelipanche spp.) in faba bean (Vicia faba) based in low induction of broomrape seed germination. Euphytica 2012, 186, 897–905. [Google Scholar] [CrossRef]
- Marzougui, A.; Rajendran, A.; Mattinson, D.S.; Ma, Y.; McGee, R.J.; Garcia-Perez, M.; Ficklin, S.P.; Sankaran, S. Evaluation of biogenic markers-based phenotyping for resistance to Aphanomyces root rot in field pea. Inf. Process. Agric. 2022, 9, 1–10. [Google Scholar] [CrossRef]
- Divyanth, L.G.; Marzougui, A.; Gonzalez-Bernal, M.J.; McGee, R.J.; Rubiales, D.; Sankaran, S. Evaluation of effective class-balancing techniques for CNN-based assessment of Aphanomyces root rot resistance in pea. Sensors 2022, 22, 7237. [Google Scholar] [CrossRef]
- Araus, J.L.; Kefauver, S.C.; Zaman-Allah, M.; Olsen, M.S.; Cairns, J.E. Translating high-throughput phenotyping into genetic gain. Trends Plant Sci. 2018, 23, 451–466. [Google Scholar] [CrossRef] [Green Version]
- Araus, J.L.; Cairns, J.E. Field high-throughput phenotyping: The new crop breeding frontier. Trends Plant Sci. 2014, 19, 52–61. [Google Scholar] [CrossRef]
- Araus, J.L.; Kefauver, S.C. Breeding to adapt agriculture to climate change: Affordable phenotyping solutions. Curr. Opin. Plant Biol. 2018, 45, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Maeda, M.; Chang, A.; Bhandari, M.; Ashapure, A.; Landivar-Bowles, J. The potential of remote sensing and artificial intelligence as tools to improve the resilience of agriculture production systems. Curr. Opin. Biotechnol. 2021, 70, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Quirós Vargas, J.J.; Zhang, C.; Smitchger, J.A.; McGee, R.J.; Sankaran, S. Phenotyping of plant biomass and performance traits using remote sensing techniques in pea (Pisum sativum L.). Sensors 2019, 19, 2031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, L.; Van De Zedde, R.; Kowalchuk, G. Recent developments and potential of robotics in plant eco-phenotyping. Emerg. Top. Life Sci. 2021, 5, 289–300. [Google Scholar] [CrossRef]
- Nguyen, G.N.; Norton, S.L.; Rosewarne, G.M.; James, L.E.; Slater, A.T. Automated phenotyping for early vigour of field pea seedlings in controlled environment by colour imaging technology. PLoS ONE 2018, 13, e0207788. [Google Scholar] [CrossRef]
- Rispail, N.; Rubiales, D. Rapid and efficient estimation of pea resistance to the soil-borne pathogen Fusarium oxysporum by infrared imaging. Sensors 2015, 15, 3988–4000. [Google Scholar] [CrossRef] [Green Version]
- Bohar, R.; Chitkineni, A.; Varshney, R.K. Genetic molecular markers to accelerate genetic gains in crops. Biotechniques 2020, 69, 158–160. [Google Scholar] [CrossRef]
- Moreira, F.F.; Oliveira, H.R.; Volenec, J.J.; Rainey, K.M.; Brito, L.F. Integrating High-Throughput Phenotyping and Statistical Genomic Methods to Genetically Improve Longitudinal Traits in Crops. Front. Plant Sci. 2020, 11, 681. [Google Scholar] [CrossRef]
- Bates, G.H. A Device for the Observation of Root Growth in the Soil. Nature 1937, 139, 966–967. [Google Scholar] [CrossRef]
- Kuijken, R.C.P.; van Eeuwijk, F.A.; Marcelis, L.F.M.; Bouwmeester, H.J. Root phenotyping: From component trait in the lab to breeding. J. Exp. Bot. 2015, 66, 5389–5401. [Google Scholar] [CrossRef] [Green Version]
- Cabrera, J.; Conesa, C.M.; del Pozo, J.C. May the dark be with roots: A perspective on how root illumination may bias in vitro research on plant–environment interactions. New Phytol. 2022, 233, 1988–1997. [Google Scholar] [CrossRef]
- Yee, M.O.; Kim, P.; Li, Y.; Singh, A.K.; Northen, T.R.; Chakraborty, R. Specialized Plant Growth Chamber Designs to Study Complex Rhizosphere Interactions. Front. Microbiol. 2021, 12, 507. [Google Scholar] [CrossRef]
- Jeudy, C.; Adrian, M.; Baussard, C.; Bernard, C.; Bernaud, E.; Bourion, V.; Busset, H.; Cabrera-Bosquet, L.; Cointault, F.; Han, S.; et al. RhizoTubes as a new tool for high throughput imaging of plant root development and architecture: Test, comparison with pot grown plants and validation. Plant Methods 2016, 12, 31. [Google Scholar] [CrossRef]
- Taylor, H.M.; Upchurch, D.R.; McMichael, B.L. Applications and limitations of rhizotrons and minirhizotrons for root studies. Plant Soil 1990, 129, 29–35. [Google Scholar] [CrossRef]
- Klepper, B.; Kaspar, T.C. Rhizotrons: Their Development and Use in Agricultural Research. Agron. J. 1994, 86, 745–753. [Google Scholar] [CrossRef]
- Bodner, G.; Nakhforoosh, A.; Arnold, T.; Leitner, D. Hyperspectral imaging: A novel approach for plant root phenotyping. Plant Methods 2018, 14, 84. [Google Scholar] [CrossRef] [Green Version]
- Rahman, G.; Sohag, H.; Chowdhury, R.; Wahid, K.A.; Dinh, A.; Arcand, M.; Vail, S. SoilCam: A fully automated minirhizotron using multispectral imaging for root activity monitoring. Sensors 2020, 20, 787. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Watson, A.; Gonzalez-Navarro, O.E.; Ramirez-Gonzalez, R.H.; Yanes, L.; Mendoza-Suárez, M.; Simmonds, J.; Wells, R.; Rayner, T.; Green, P.; et al. Speed breeding in growth chambers and glasshouses for crop breeding and model plant research. Nat. Protoc. 2018, 13, 2944–2963. [Google Scholar] [CrossRef] [Green Version]
- Bhatta, M.; Sandro, P.; Smith, M.R.; Delaney, O.; Voss-Fels, K.P.; Gutierrez, L.; Hickey, L.T. Need for speed: Manipulating plant growth to accelerate breeding cycles. Curr. Opin. Plant Biol. 2021, 60, 101986. [Google Scholar] [CrossRef]
- Rubiales, D.; Pérez-de-Luque, A.; Joel, D.M.; Alcántara, C.; Sillero, J.C. Characterization of resistance in chickpea to crenate broomrape (Orobanche crenata). Weed Sci. 2003, 51, 702–707. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Kisugi, T.; Xie, X.; Rubiales, D.; Yoneyama, K. Low strigolactone root exudation: A novel mechanism of broomrape (Orobanche and Phelipanche spp.) resistance available for faba bean breeding. J. Agric. Food Chem. 2014, 62, 7063–7071. [Google Scholar] [CrossRef]
- Gurney, A.L.; Grimanelli, D.; Kanampiu, F.; Hoisington, D.; Scholes, J.D.; Press, M.C. Novel sources of resistance to Striga hermonthica in Tripsacum dactyloides, a wild relative of maize. New Phytol. 2003, 160, 557–568. [Google Scholar] [CrossRef] [Green Version]
- Gurney, A.L.; Slate, J.; Press, M.C.; Scholes, J.D. A novel form of resistance in rice to the angiosperm parasite Striga hermonthica. New Phytol. 2006, 169, 199–208. [Google Scholar] [CrossRef]
- Kavuluko, J.; Kibe, M.; Sugut, I.; Kibet, W.; Masanga, J.; Mutinda, S.; Wamalwa, M.; Magomere, T.; Odeny, D.; Runo, S. GWAS provides biological insights into mechanisms of the parasitic plant (Striga) resistance in sorghum. BMC Plant Biol. 2021, 21, 392. [Google Scholar] [CrossRef]
- Esquinas-Alcázar, J. Protecting crop genetic diversity for food security: Political, ethical and technical challenges. Nat. Rev. Genet. 2005, 6, 946–953. [Google Scholar] [CrossRef]
- Bariana, H.S.; Bansal, U.K. Breeding for Disease Resistance. Encycl. Appl. Plant Sci. 2016, 3, 69–76. [Google Scholar] [CrossRef]
- Varshney, R.K.; Glaszmann, J.C.; Leung, H.; Ribaut, J.M. More genomic resources for less-studied crops. Trends Biotechnol. 2010, 28, 452–460. [Google Scholar] [CrossRef] [Green Version]
- Varshney, R.K.; Dubey, A. Novel genomic tools and modern genetic and breeding approaches for crop improvement. J. Plant Biochem. Biotechnol. 2009, 18, 127–138. [Google Scholar] [CrossRef] [Green Version]
- Annicchiarico, P.; Nazzicari, N.; Pecetti, L.; Romani, M.; Ferrari, B.; Wei, Y.; Brummer, E.C. GBS-Based Genomic Selection for Pea Grain Yield under Severe Terminal Drought. Plant Genome 2017, 10, plantgenome2016.07.0072. [Google Scholar] [CrossRef]
- Leonforte, A.; Sudheesh, S.; Cogan, N.O.I.; Salisbury, P.A.; Nicolas, M.E.; Materne, M.; Forster, J.W.; Kaur, S. SNP marker discovery, linkage map construction and identification of QTLs for enhanced salinity tolerance in field pea (Pisum sativum L.). BMC Plant Biol. 2013, 13, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aryamanesh, N.; Zeng, Y.; Byrne, O.; Hardie, D.C.; Al-Subhi, A.M.; Khan, T.; Siddique, K.H.M.; Yan, G. Identification of genome regions controlling cotyledon, pod wall/seed coat and pod wall resistance to pea weevil through QTL mapping. Theor. Appl. Genet. 2014, 127, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Dachapak, S.; Somta, P.; Naito, K.; Tomooka, N.; Kaga, A.; Srinives, P. Detection of quantitative trait loci for salt tolerance in zombi pea [Vigna vexillata (L.) A. Rich]. Euphytica 2019, 215, 208. [Google Scholar] [CrossRef]
- Barilli, E.; Carrillo-Perdomo, E.; Cobos, M.J.; Kilian, A.; Carling, J.; Rubiales, D. Identification of potential candidate genes controlling pea aphid tolerance in a Pisum fulvum high-density integrated DArTseq SNP-based genetic map. Pest Manag. Sci. 2020, 76, 1731–1742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, X.F.; Li, Y.B.; Han, X.; Ahmad, W.; Li, Q. Using high-throughput sequencing quantitatively to investigate soil nematode community composition in a steppe-forest ecotone. Appl. Soil Ecol. 2020, 152, 103562. [Google Scholar] [CrossRef]
- Nasiri, J.; Haghnazari, A.; Saba, J. Genetic diversity among varieties and wild species accessions of pea (Pisum sativum L.) based on SSR markers. Afr. J. Biotechnol. 2009, 8, 3405–3417. [Google Scholar] [CrossRef]
- Siol, M.; Jacquin, F.; Chabert-Martinello, M.; Smýkal, P.; Le Paslier, M.C.; Aubert, G.; Burstin, J. Patterns of genetic structure and linkage disequilibrium in a large collection of pea germplasm. G3 2017, 7, 2461–2471. [Google Scholar] [CrossRef] [Green Version]
- Varshney, R.K. Exciting journey of 10 years from genomes to fields and markets: Some success stories of genomics-assisted breeding in chickpea, pigeonpea and groundnut. Plant Sci. 2016, 242, 98–107. [Google Scholar] [CrossRef] [Green Version]
- Kankanala, P.; Nandety, R.S.; Mysore, K.S. Genomics of Plant Disease Resistance in Legumes. Front. Plant Sci. 2019, 10, 1345. [Google Scholar] [CrossRef]
- Le Signor, C.; Aimé, D.; Bordat, A.; Belghazi, M.; Labas, V.; Gouzy, J.; Young, N.D.; Prosperi, J.M.; Leprince, O.; Thompson, R.D.; et al. Genome-wide association studies with proteomics data reveal genes important for synthesis, transport and packaging of globulins in legume seeds. New Phytol. 2017, 214, 1597–1613. [Google Scholar] [CrossRef] [Green Version]
- Zitnick-Anderson, K.; Oladzadabbasabadi, A.; Jain, S.; Modderman, C.; Osorno, J.M.; McClean, P.E.; Pasche, J.S. Sources of Resistance to Fusarium solani and Associated Genomic Regions in Common Bean Diversity Panels. Front. Genet. 2020, 11, 475. [Google Scholar] [CrossRef]
- Kwon, S.J.; Smýkal, P.; Hu, J.; Wang, M.; Kim, S.J.; McGee, R.J.; McPhee, K.; Coyne, C.J. User-friendly markers linked to Fusarium wilt race 1 resistance Fw gene for marker-assisted selection in pea. Plant Breed. 2013, 132, 642–648. [Google Scholar] [CrossRef]
- Shirasawa, K.; Sasaki, K.; Hirakawa, H.; Isobe, S. Genomic region associated with pod color variation in pea (Pisum sativum). G3 Genes Genomes Genet. 2021, 11, jkab081. [Google Scholar] [CrossRef]
- Dinglasan, E.; Periyannan, S.; Hickey, L.T. Harnessing adult-plant resistance genes to deploy durable disease resistance in crops. Essays Biochem. 2022, 66, 571–580. [Google Scholar]
- Castillejo, M.Á.; Fernández-Aparicio, M.; Rubiales, D. Proteomic analysis by two-dimensional differential in gel electrophoresis (2D DIGE) of the early response of Pisum sativum to Orobanche crenata. J. Exp. Bot. 2012, 63, 107–119. [Google Scholar] [CrossRef]
- Castillejo, M.Á.; Bani, M.; Rubiales, D. Understanding pea resistance mechanisms in response to Fusarium oxysporum through proteomic analysis. Phytochemistry 2015, 115, 44–58. [Google Scholar] [CrossRef]
- Burstin, J.; Salloignon, P.; Chabert-Martinello, M.; Magnin-Robert, J.B.; Siol, M.; Jacquin, F.; Chauveau, A.; Pont, C.; Aubert, G.; Delaitre, C.; et al. Genetic diversity and trait genomic prediction in a pea diversity panel. BMC Genom. 2015, 16, 105. [Google Scholar] [CrossRef]
- Annicchiarico, P.; Nazzicari, N.; Laouar, M.; Thami-Alami, I.; Romani, M.; Pecetti, L. Development and proof-of-concept application of genome-enabled selection for pea grain yield under severe terminal drought. Int. J. Mol. Sci. 2020, 21, 2414. [Google Scholar] [CrossRef] [Green Version]
- Bohra, A.; Pandey, M.K.; Jha, U.C.; Singh, B.; Singh, I.P.; Datta, D.; Chaturvedi, S.K.; Nadarajan, N.; Varshney, R.K. Genomics-assisted breeding in four major pulse crops of developing countries: Present status and prospects. Theor. Appl. Genet. 2014, 127, 1263–1291. [Google Scholar] [CrossRef] [Green Version]
- Varshney, R.K.; Tuberosa, R. Translational genomics in crop breeding for biotic stress resistance: An introduction. Transl. Genom. Crop Breed. Vol. I Biot. Stress 2013, 1, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Zargar, S.M.; Raatz, B.; Sonah, H.; Muslima, N.; Bhat, J.A.; Dar, Z.A.; Agrawal, G.K.; Rakwal, R. Recent advances in molecular marker techniques: Insight into QTL mapping, GWAS and genomic selection in plants. J. Crop Sci. Biotechnol. 2015, 18, 293–308. [Google Scholar] [CrossRef]
- Sagan, M.; Huguet, T.; Duc, G. Phenotypic characterization and classification of nodulation mutants of pea (Pisum sativum L.). Plant Sci. 1994, 100, 59–70. [Google Scholar] [CrossRef]
- Dalmais, M.; Schmidt, J.; Le Signor, C.; Moussy, F.; Burstin, J.; Savois, V.; Aubert, G.; Brunaud, V.; De Oliveira, Y.; Guichard, C.; et al. UTILLdb, a Pisum sativum in silicoforward and reverse genetics tool. Genome Biol. 2008, 9, R43. [Google Scholar] [CrossRef] [Green Version]
- Tayeh, N.; Aubert, G.; Pilet-Nayel, M.L.; Lejeune-Hénaut, I.; Warkentin, T.D.; Burstin, J. Genomic tools in pea breeding programs: Status and perspectives. Front. Plant Sci. 2015, 6, 1037. [Google Scholar] [CrossRef] [Green Version]
- Gupta, M.; Dubey, S.; Jain, D.; Chandran, D. The Medicago truncatula sugar transport protein 13 and its Lr67res-like variant confer powdery mildew resistance in legumes via defense modulation. Plant Cell Physiol. 2021, 62, 650–667. [Google Scholar] [CrossRef]
- Benhamou, N.; Garand, C. Cytological analysis of defense-related mechanisms induced in pea root tissues in response to colonization by nonpathogenic Fusarium oxysporum Fo47. Phytopathology 2001, 91, 730–740. [Google Scholar] [CrossRef] [Green Version]
- Gali, K.K.; Tar’an, B.; Madoui, M.A.; van der Vossen, E.; van Oeveren, J.; Labadie, K.; Berges, H.; Bendahmane, A.; Lachagari, R.V.; Burstin, J.; et al. Development of a sequence-based reference physical map of pea (Pisum sativum L.). Front. Plant Sci. 2019, 10, 323. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wohor, O.Z.; Rispail, N.; Ojiewo, C.O.; Rubiales, D. Pea Breeding for Resistance to Rhizospheric Pathogens. Plants 2022, 11, 2664. https://doi.org/10.3390/plants11192664
Wohor OZ, Rispail N, Ojiewo CO, Rubiales D. Pea Breeding for Resistance to Rhizospheric Pathogens. Plants. 2022; 11(19):2664. https://doi.org/10.3390/plants11192664
Chicago/Turabian StyleWohor, Osman Z., Nicolas Rispail, Chris O. Ojiewo, and Diego Rubiales. 2022. "Pea Breeding for Resistance to Rhizospheric Pathogens" Plants 11, no. 19: 2664. https://doi.org/10.3390/plants11192664
APA StyleWohor, O. Z., Rispail, N., Ojiewo, C. O., & Rubiales, D. (2022). Pea Breeding for Resistance to Rhizospheric Pathogens. Plants, 11(19), 2664. https://doi.org/10.3390/plants11192664