

Novel Insights into the Nature of Intraspecific Genome Size Diversity in Cannabis sativa L.

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Results and Discussion
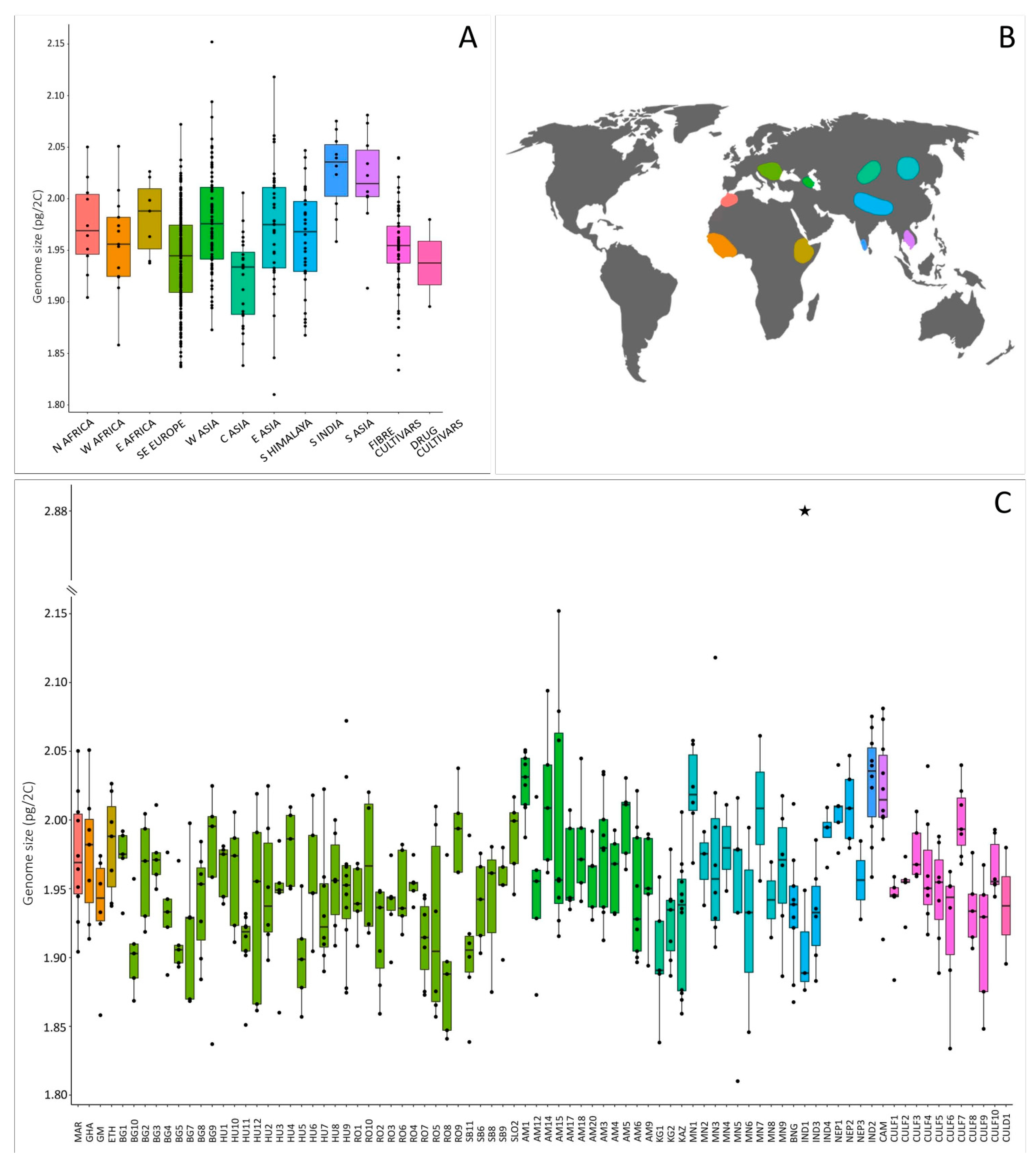
2.1. Genome Size in Cannabis: Evidence of Intraspecific Variation
2.2. Potential Factors Influencing Genome Size Variation in Cannabis
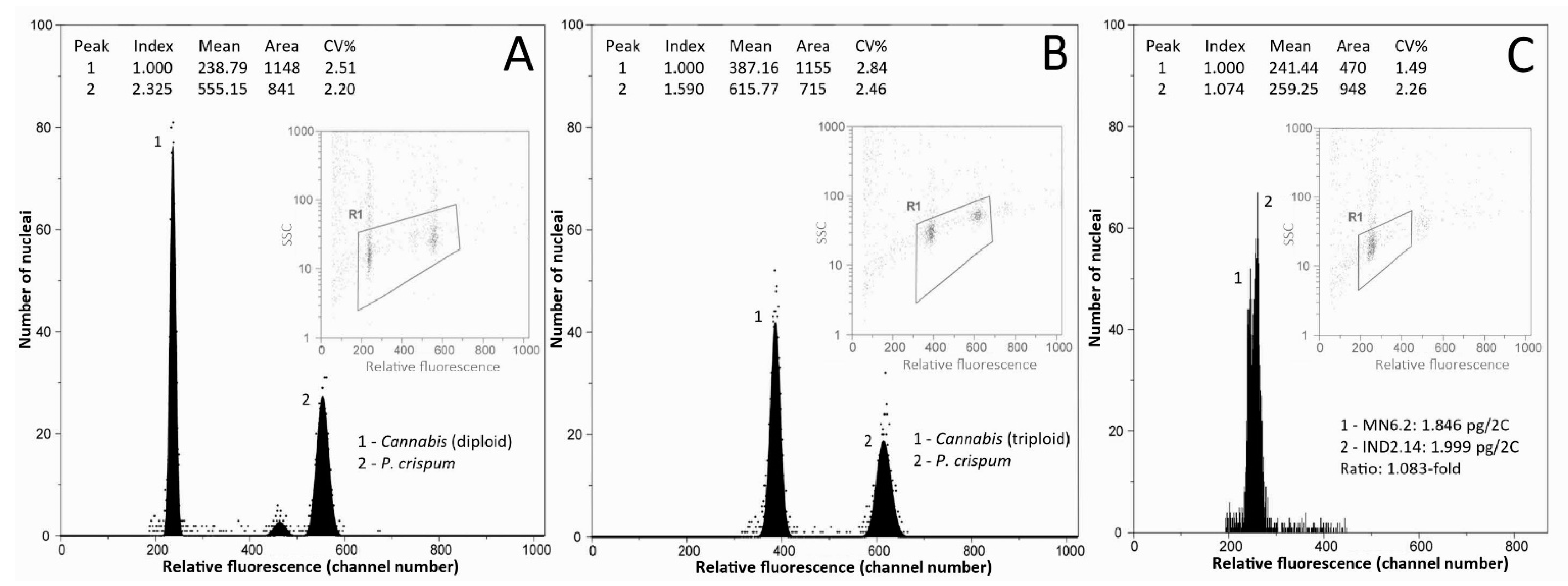
2.3. Events of Polyploidy in Cannabis Are Extremely Rare
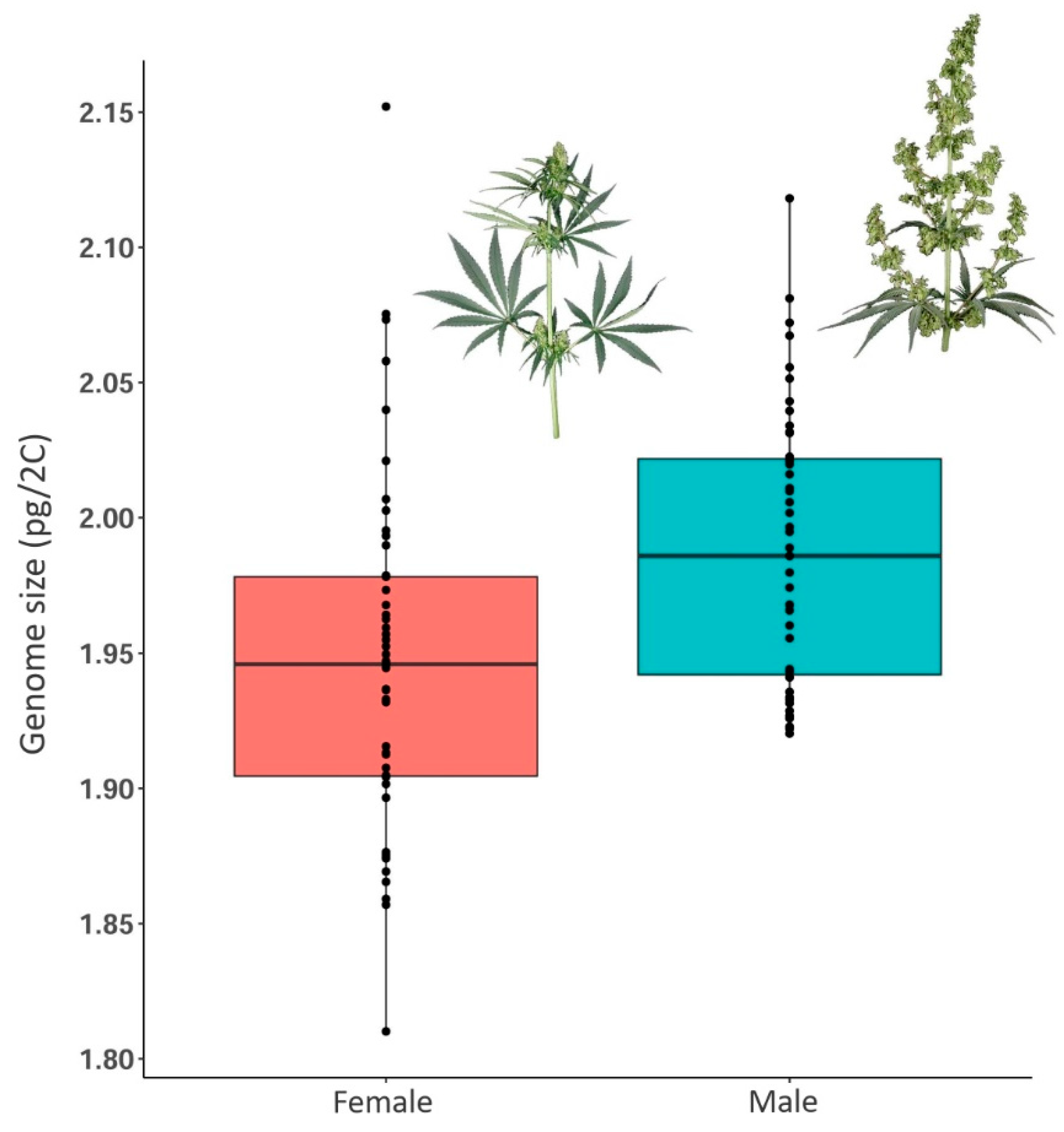
2.4. Differences in Genome Size Values of Male and Female Cannabis Individuals
2.5. Sex Determination in Cannabis Using Flow Cytometry
3. Materials and Methods
3.1. Plant Sampling and Cultivation
3.2. Flow Cytometry Measurements
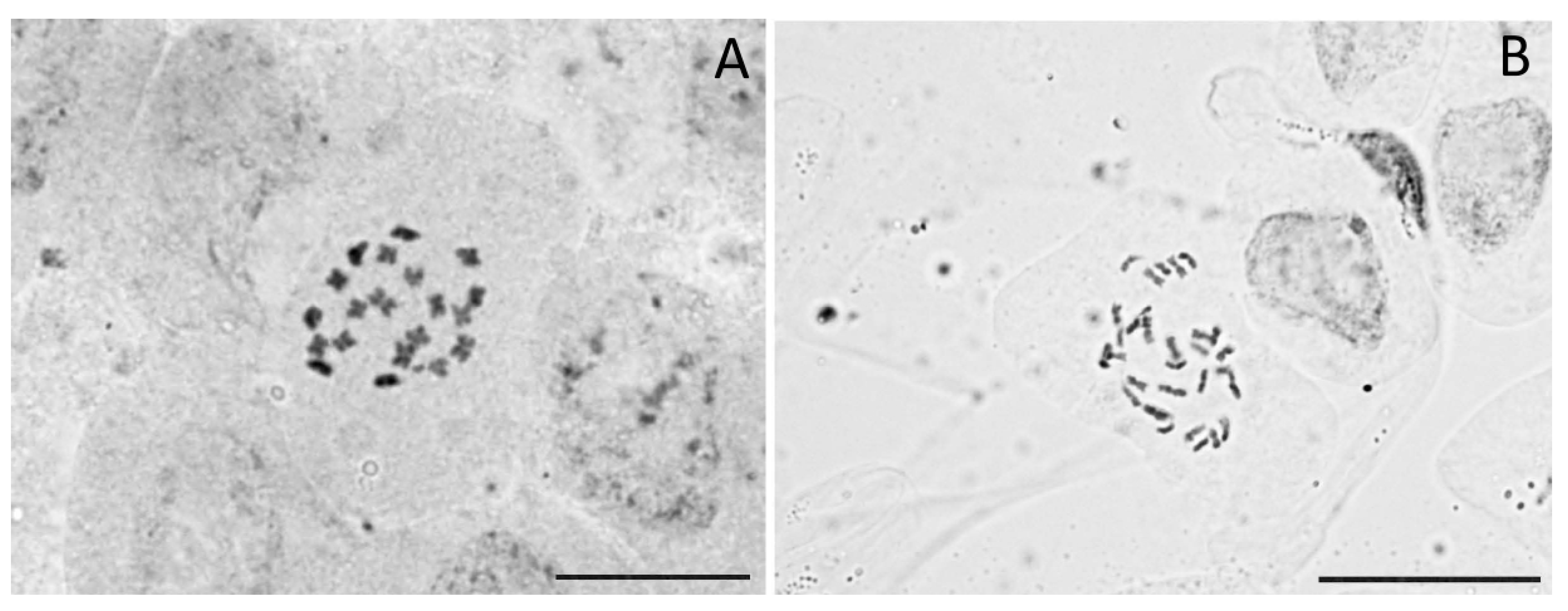
3.3. Chromosome Counts
3.4. Sex Determination Using Male-Associated Marker and Flow Cytometry
3.5. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clarke, R.; Merlin, M. Cannabis: Evolution and Ethnobotany, 1st ed.; University of California Press: Berkley, MI, USA; Los Angeles, CA, USA; London, UK, 2013; ISBN 9780520270480. [Google Scholar]
- Gedik, G.; Avinc, O. Hemp fiber as a sustainable raw material source for textile industry: Can we use its potential for more eco-friendly production? In Sustainability in the Textile and Apparel Industries. Sustainable Textiles: Production, Processing, Manufacturing & Chemistry; Muthu, S., Gardetti, M., Eds.; Springer: Cham, Switzerland, 2020; pp. 87–109. ISBN 9783030385415. [Google Scholar]
- Rupasinghe, H.P.V.; Davis, A.; Kumar, S.K.; Murray, B.; Zheljazkov, V.D. Industrial hemp (Cannabis sativa subsp. sativa) as an emerging source for value-added functional food ingredients and nutraceuticals. Molecules 2020, 25, 4078. [Google Scholar]
- Gomez, F.P.; Hu, J.; Clarke, M.A. Cannabis as a Feedstock for the Production of Chemicals, Fuels, and Materials: A Review of Relevant Studies to Date. Energy Fuels 2021, 35, 5538–5557. [Google Scholar] [CrossRef]
- McPartland, J.M.; Hegman, W.; Long, T. Cannabis in Asia: Its center of origin and early cultivation, based on a synthesis of subfossil pollen and archaeobotanical studies. Veg. Hist. Archaeobot. 2019, 28, 691–702. [Google Scholar] [CrossRef]
- Jin, J.J.; Yang, M.Q.; Fritsch, P.W.; van Velzen, R.; Li, D.Z.; Yi, T.S. Born migrators: Historical biogeography of the cosmopolitan family Cannabaceae. J. Syst. Evol. 2020, 58, 461–473. [Google Scholar] [CrossRef]
- Ren, G.; Zhang, X.; Li, Y.; Ridout, K.; Serrano-Serrano, M.L.; Yang, Y.; Liu, A.; Ravikanth, G.; Nawaz, M.A.; Mumtaz, A.S.; et al. Large-scale whole-genome sequencing unravels the domestication history of Cannabis sativa. Sci. Adv. 2021, 7, eabg2286. [Google Scholar] [CrossRef]
- Kovalchuk, I.; Pellino, M.; Rigault, P.; van Velzen, R.; Ebersbach, J.; Ashnest, J.R.; Mau, M.; Schranz, M.E.; Alcorn, J.; Laprairie, R.B.; et al. The genomics of Cannabis and its close relatives. Annu. Rev. Plant Biol. 2020, 71, 20.1–20.27. [Google Scholar] [CrossRef] [Green Version]
- Hesami, M.; Pepe, M.; Alizadeh, M.; Rakei, A.; Baiton, A.; Phineas Jones, A.M. Recent advances in cannabis biotechnology. Ind. Crops Prod. 2020, 158, 113026. [Google Scholar] [CrossRef]
- Small, E. Evolution and Classification of Cannabis sativa (Marijuana, Hemp) in Relation to Human Utilization. Bot. Rev. 2015, 81, 189–294. [Google Scholar] [CrossRef]
- Hurgobin, B.; Tamiru-Oli, M.; Welling, M.T.; Doblin, M.S.; Bacic, A.; Whelan, J.; Lewsey, M.G. Recent advances in Cannabis sativa genomics research. New Phytol. 2021, 230, 73–89. [Google Scholar] [CrossRef]
- Watts, S.; McElroy, M.; Migicovsky, Z.; Maassen, H.; van Velzen, R.; Myles, S. Cannabis labelling is associated with genetic variation in terpene synthase genes. Nat. Plants 2021, 7, 1330–1334. [Google Scholar] [CrossRef]
- Hazekamp, A.; Fischedick, J.T. Cannabis—From cultivar to chemovar. Drug Test. Anal. 2012, 4, 660–667. [Google Scholar] [CrossRef]
- Bautista, J.L.; Yu, S.; Tian, L. Flavonoids in Cannabis sativa: Biosynthesis, Bioactivities, and Biotechnology. ACS Omega 2021, 6, 5119–5123. [Google Scholar] [CrossRef]
- Mcpartland, J.M.; Small, E. A classification of endangered high-THC cannabis (Cannabis sativa subsp. indica) domesticates and their wild relatives. PhytoKeyes 2020, 144, 81–112. [Google Scholar] [CrossRef] [Green Version]
- Greilhuber, J.; Doležel, J.; Lysák, M.A.; Bennett, M.D. The origin, evolution and proposed stabilization of the terms “genome size” and “C-value” to describe nuclear DNA contents. Ann. Bot. 2005, 95, 255–260. [Google Scholar] [CrossRef]
- Bennett, M.D.; Leitch, I.J. Genome Size Evolution in Plants. In The Evolution of the Genome; Gregory, T.R., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2005; pp. 89–162. ISBN 9780123014634. [Google Scholar]
- Pellicer, J.; López-Pujol, J.; Aixarch, M.; Garnatje, T.; Vallès, J.; Hidalgo, O. Detecting introgressed populations in the Iberian endemic Centaurea podospermifolia through genome size. Plants 2021, 10, 1492. [Google Scholar] [CrossRef]
- Fernández, P.; Hidalgo, O.; Juan, A.; Leitch, I.J.; Leitch, A.R.; Palazzesi, L.; Pegoraro, L.; Viruel, J.; Pellicer, J. Genome Insights into Autopolyploid Evolution: A Case Study in Senecio doronicum (Asteraceae) from the Southern Alps. Plants 2022, 11, 1235. [Google Scholar] [CrossRef]
- González, G.E.; Poggio, L. Intragenomic conflict between knob heterochromatin and b chromosomes is the key to understand genome size variation along altitudinal clines in maize. Plants 2021, 10, 1859. [Google Scholar] [CrossRef]
- Costich, D.E.; Meagher, T.R.; Yurkow, J.E. A rapid means of sex identification in Silene latifolia by use of flow cytometry. Plant Mol. Biol. Report. 1991, 9, 359–370. [Google Scholar] [CrossRef]
- Doležel, J.; Göhde, W. Sex determination in dioecious plants Melandrium album and M. rubrum using high-resolution flow cytometry. Cytometry 1995, 19, 103–106. [Google Scholar] [CrossRef]
- Zhang, S.J.; Liu, L.; Yang, R.; Wang, X. Genome Size Evolution Mediated by Gypsy Retrotransposons in Brassicaceae. Genom. Proteom. Bioinforma 2020, 18, 321–332. [Google Scholar] [CrossRef]
- Becher, H.; Powell, R.F.; Brown, M.R.; Metherell, C.; Pellicer, J.; Leitch, I.J.; Twyford, A.D. The nature of intraspecific and interspecific genome size variation in taxonomically complex eyebrights. Ann. Bot. 2021, 128, 639–651. [Google Scholar] [CrossRef] [PubMed]
- Devos, K.M.; Brown, J.K.M.; Bennetzen, J.L. Genome size reduction through illegitimate recombination counteracts genome expansion in Arabidopsis. Genome Res. 2002, 12, 1075–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Achigan-Dako, E.G.; Fuchs, J.; Ahanchede, A.; Blattner, F.R. Flow cytometric analysis in Lagenaria siceraria (Cucurbitaceae) indicates correlation of genome size with usage types and growing elevation. Plant Syst. Evol. 2008, 276, 9–19. [Google Scholar] [CrossRef]
- Benor, S.; Fuchs, J.; Blattner, F.R. Genome size variation in Corchorus olitorius (Malvaceae s.l.) and its correlation with elevation and phenotypic traits. Genome 2011, 54, 575–585. [Google Scholar] [CrossRef]
- García-Fernández, A.; Iriondo, J.M.; Vallès, J.; Orellana, J.; Escudero, A. Ploidy level and genome size of locally adapted populations of Silene ciliata across an altitudinal gradient. Plant Syst. Evol. 2012, 298, 139–146. [Google Scholar] [CrossRef]
- Díez, C.M.; Gaut, B.S.; Meca, E.; Scheinvar, E.; Montes-Hernandez, S.; Eguiarte, L.E.; Tenaillon, M.I. Genome size variation in wild and cultivated maize along altitudinal gradients. New Phytol. 2013, 199, 264–276. [Google Scholar] [CrossRef] [Green Version]
- Bilinski, P.; Albert, P.S.; Berg, J.J.; Birchler, J.A.; Grote, M.N.; Lorant, A.; Quezada, J.; Swarts, K.; Yang, J.; Ross-Ibarra, J. Parallel altitudinal clines reveal trends in adaptive evolution of genome size in Zea mays. PLoS Genet. 2018, 14, e1007162. [Google Scholar] [CrossRef] [Green Version]
- Walker, D.J.; Moñino, I.; Correal, E. Genome size in Bituminaria bituminosa (L.) C.H. Stirton (Fabaceae) populations: Separation of “true” differences from environmental effects on DNA determination. Environ. Exp. Bot. 2006, 55, 258–265. [Google Scholar] [CrossRef]
- Pellicer, J.; Vallès, J.; Garcia, S.; Garnatje, T. Changes in genome size in a fragmented distribution area: The case of Artemisia crithmifolia L. (Asteraceae, Anthemideae). Caryologia 2009, 62, 152–160. [Google Scholar] [CrossRef] [Green Version]
- Cronk, Q.; Hidalgo, O.; Pellicer, J.; Percy, D.; Leitch, I.J. Salix transect of Europe: Variation in ploidy and genome size in willow-associated common nettle, Urtica dioica L. sens. lat., from Greece to arctic Norway. Biodivers. Data J. 2016, 4, e10003. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Y.; Hamernick, S.; Ortiz, J.B.; Watkins, E. DNA content and ploidy estimation of Festuca ovina accessions by flow cytometry. Crop Sci. 2020, 60, 2757–2767. [Google Scholar] [CrossRef]
- Small, E.; Cronquist, A. A Practical and Natural Taxonomy for Cannabis. Taxon 1976, 25, 405–435. [Google Scholar] [CrossRef]
- Hirata, K. Sex determination in hemp (Cannabis sativa L.). J. Genet. 1925, 19, 65–79. [Google Scholar] [CrossRef]
- Faux, A.M.; Berhin, A.; Dauguet, N.; Bertin, P. Sex chromosomes and quantitative sex expression in monoecious hemp (Cannabis sativa L.). Euphytica 2014, 196, 183–197. [Google Scholar] [CrossRef]
- Soorni, A.; Fatahi, R.; Haak, D.C.; Salami, S.A.; Bombarely, A. Assessment of Genetic Diversity and Population Structure in Iranian Cannabis Germplasm. Sci. Rep. 2017, 7, 15668. [Google Scholar] [CrossRef] [Green Version]
- Bocharkina, Y.V.; Alexandrov, O.S.; Razumova, O.V.; Karlov, G.I. The comparative analysis of the DNA repeat composition among Cannabis sativa L., Humulus lupulus L. and Humulus japonicus Siebold & Zucc. with heteromorphic sex chromosomes. Preprints 2021, 2021120040. [Google Scholar] [CrossRef]
- Grassa, C.J.; Weiblen, G.D.; Wenger, J.P.; Dabney, C.; Poplawski, S.G.; Motley, S.T.; Michael, T.P.; Schwartz, C.J. A new Cannabis genome assembly associates elevated cannabidiol (CBD) with hemp introgressed into marijuana. New Phytol. 2021, 230, 1665–1679. [Google Scholar] [CrossRef]
- Márquez, F.; Lozada, M.; Idaszkin, Y.L.; González-José, R.; Bigatti, G. Cannabis Varieties Can Be Distinguished by Achene Shape Using Geometric Morphometrics. Cannabis Cannabinoid Res. 2022, 7, 409–414. [Google Scholar] [CrossRef]
- Petit, J.; Salentijn, E.M.J.; Paulo, M.J.; Thouminot, C.; van Dinter, B.J.; Magagnini, G.; Gusovius, H.J.; Tang, K.; Amaducci, S.; Wang, S.; et al. Genetic Variability of Morphological, Flowering, and Biomass Quality Traits in Hemp (Cannabis sativa L.). Front. Plant Sci. 2020, 11, 102. [Google Scholar] [CrossRef] [Green Version]
- Vergara, D.; Feathers, C.; Huscher, E.L.; Holmes, B.; Haas, J.A.; Kane, N.C. Widely assumed phenotypic associations in Cannabis sativa lack a shared genetic basis. PeerJ 2021, 9, e10672. [Google Scholar] [CrossRef]
- Jin, D.; Dai, K.; Xie, Z.; Jie, C. Secondary metabolites profiled in Cannabis inflorescences, leaves, stem barks, and roots for medicinal purposes. Sci. Rep. 2020, 10, 3309. [Google Scholar] [CrossRef]
- Busta, L.; Dweikat, I.; Sato, S.J.; Qu, H.; Xue, Y.; Zhou, B.; Gan, L.; Yu, B.; Clemente, T.E.; Cahoon, E.B.; et al. Chemical and genetic variation in feral Cannabis sativa populations across the Nebraska climate gradient. Phytochemistry 2022, 200, 113206. [Google Scholar] [CrossRef] [PubMed]
- Di Sotto, A.; Gullì, M.; Acquaviva, A.; Tacchini, M.; Di Simone, S.C.; Chiavaroli, A.; Recinella, L.; Leone, S.; Brunetti, L.; Orlando, G.; et al. Phytochemical and pharmacological profiles of the essential oil from the inflorescences of the Cannabis sativa L. Ind. Crop. Prod. 2022, 183, 114980. [Google Scholar] [CrossRef]
- Sharma, K.; Rana, R.; Ashawat, M.S. Preliminary Phytochemical Screening of Leaves, Stems and Roots of wild Cannabis sativa. Res. J. Pharmacogn. Phytochem. 2022, 14, 19–22. [Google Scholar] [CrossRef]
- Kubešová, M.; Moravcová, L.; Suda, J.; Jarošík, V.; Pyšek, P. Naturalized plants have smaller genomes than their non-invading relatives: A flow cytometric analysis of the Czech alien flora. Preslia 2010, 82, 81–96. [Google Scholar]
- Sakamoto, K.; Akiyama, Y.; Fukui, K.; Kamada, H.; Satoh, S. Characterization; Genome Sizes and Morphology of Sex Chromosomes in Hemp (Cannabis sativa L.). Cytologia (Tokyo) 1998, 63, 459–464. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.-J.; Uchiyama, H.; Sekita, S.; Goda, Y.; Satake, M. Analytical studies on Cannabis sativa L. introduced into Japan. Part III. The range variation of genome size in C. sativa. Foods Food Ingred. J. Jpn. 2003, 208, 382–386. [Google Scholar]
- Parsons, J.L.; Martin, S.L.; James, T.; Golenia, G.; Boudko, E.A.; Hepworth, S.R. Polyploidization for the Genetic Improvement of Cannabis sativa. Front. Plant Sci. 2019, 10, 476. [Google Scholar] [CrossRef]
- Sharma, V.; Srivastava, D.K.; Gupta, R.C.; Singh, B. Abnormal Meiosis in Tetraploid (4x) Cannabis sativa (L.) from Lahaul-Spiti (Cold Desert Higher Altitude Himalayas)-A Neglected But Important Herb. J. Biol. Chem. Chron. 2015, 2, 38–42. [Google Scholar]
- Small, E. Interfertility and chromosomal uniformity in Cannabis. Can. J. Bot. 1972, 50, 1947–1949. [Google Scholar] [CrossRef]
- Bagheri, M.; Mansouri, H. Effect of Induced Polyploidy on Some Biochemical Parameters in Cannabis sativa L. Appl. Biochem. Biotechnol. 2015, 175, 2366–2375. [Google Scholar] [CrossRef]
- Mansouri, H.; Bagheri, M. Induction of Polyploidy and Its Effect on Cannabis sativa L. In Cannabis sativa L.—Botany and Biotechnology; Chandra, S., Lata, H., ElSohly, M.A., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 365–383. ISBN 9783319545646. [Google Scholar]
- Galán-Ávila, A.; García-Fortea, E.; Prohens, J.; Herraiz, F.J. Development of a Direct in vitro Plant Regeneration Protocol From Cannabis sativa L. Seedling Explants: Developmental Morphology of Shoot Regeneration and Ploidy Level of Regenerated Plants. Front. Plant Sci. 2020, 11, 645. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, L.E.; Brand, M.H.; Lubell-Brand, J.D. Production of tetraploid and triploid hemp. HortScience 2020, 55, 1703–1707. [Google Scholar] [CrossRef]
- Lynch, R.C.; Vergara, D.; Tittes, S.; White, K.; Schwartz, C.J.; Gibbs, M.J.; Ruthenburg, T.C.; DeCesare, K.; Land, D.P.; Kane, N.C. Genomic and Chemical Diversity in Cannabis. CRC. Crit. Rev. Plant Sci. 2016, 35, 349–363. [Google Scholar] [CrossRef]
- McPhee, H.C. The genetics of sex in Hemp. J. Agric. Res. 1926, 31, 935–943. [Google Scholar]
- Westergaard, M. The Mechanism of Sex Determination in Dioecious Flowering Plants. Adv. Genet. 1958, 9, 217–281. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, K.; Abe, T.; Matsuyama, T.; Yoshida, S.; Ohmido, N.; Fukui, K.; Satoh, S. RAPD markers encoding retrotransposable elements are linked to the male sex in Cannabis sativa L. Genome 2005, 48, 931–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandolino, G.; Carboni, A.; Forapani, S.; Faeti, V.; Ranalli, P. Identification of DNA markers linked to the male sex in dioecious hemp (Cannabis sativa L.). Theor. Appl. Genet. 1999, 98, 86–92. [Google Scholar] [CrossRef]
- Flachowsky, H.; Schumann, E.; Weber, W.E.; Peil, A. Application of AFLP for the detection of sex-specific markers in hemp. Plant Breed. 2001, 120, 305–309. [Google Scholar] [CrossRef]
- Törjék, O.; Bucherna, N.; Kiss, E.; Homoki, H.; Finta-Korpelová, Z.; Bócsa, I.; Nagy, I.; Heszky, L.E. Novel male-specific molecular markers (MADC5, MADC6) in hemp. Euphytica 2002, 127, 209–218. [Google Scholar] [CrossRef]
- Peil, A.; Flachowsky, H.; Schumann, E.; Weber, W.E. Sex-linked AFLP markers indicate a pseudoautosomal region in hemp (Cannabis sativa L.). Theor. Appl. Genet. 2003, 107, 102–109. [Google Scholar] [CrossRef]
- Sakamoto, K.; Shimomura, K.; Komeda, Y.; Kamada, H.; Satoh, S. A Male-Associated DNA Sequence in a Dioecious Plant, Cannabis sativa L. Plant Cell Physiol. 1995, 36, 1549–1554. [Google Scholar] [CrossRef] [Green Version]
- Toth, J.A.; Stack, G.M.; Cala, A.R.; Carlson, C.H.; Wilk, R.L.; Crawford, J.L.; Viands, D.R.; Philippe, G.; Smart, C.D.; Rose, J.K.C.; et al. Development and validation of genetic markers for sex and cannabinoid chemotype in Cannabis sativa L. GCB Bioenergy 2020, 12, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Mandolino, G.; Carboni, A. Potential of marker-assisted selection in hemp genetic improvement. Euphytica 2004, 140, 107–120. [Google Scholar] [CrossRef]
- Doležel, J.; Greilhuber, J.; Lucretti, S.; Meister, A.; Lysák, M.A.; Nardi, L.; Obermayer, R. Plant genome size estimation by flow cytometry: Inter-laboratory comparison. Ann. Bot. 1998, 82, 17–26. [Google Scholar] [CrossRef]
- Šmarda, P.; Bureš, P. Intraspecific DNA content variability in Festuca pallens on different geographical scales and ploidy levels. Ann. Bot. 2006, 98, 665–678. [Google Scholar] [CrossRef] [Green Version]
- Slovák, M.; Vít, P.; Urfus, T.; Suda, J. Complex pattern of genome size variation in a polymorphic member of the Asteraceae. J. Biogeogr. 2009, 36, 372–384. [Google Scholar] [CrossRef]
- Suda, J.; Weiss-Schneeweiss, H.; Tribsch, A.; Schneeweiss, G.M.; Trávníček, P.; Schönswetter, P. Complex distribution patterns of di-, tetra-, and hexaploid cytotypes in the European high mountain plant Senecio carniolicus (Asteraceae). Am. J. Bot. 2007, 94, 1391–1401. [Google Scholar] [CrossRef] [Green Version]
- Cires, E.; Cuesta, C.; Revilla, M.Á.; Fernández Prieto, J.A. Intraspecific genome size variation and morphological differentiation of Ranunculus parnassifolius (Ranunculaceae), an Alpine-Pyrenean-Cantabrian polyploid group. Biol. J. Linn. Soc. 2010, 101, 251–271. [Google Scholar] [CrossRef] [Green Version]
- Vít, P.; Krak, K.; Trávníček, P.; Douda, J.; Lomonosova, M.N.; Mandák, B. Genome size stability across Eurasian Chenopodium species (Amaranthaceae). Bot. J. Linn. Soc. 2016, 182, 637–649. [Google Scholar] [CrossRef] [Green Version]
- Kolano, B.; Siwinska, D.; Pando, L.G.; Szymanowska-Pulka, J.; Maluszynska, J. Genome size variation in Chenopodium quinoa (Chenopodiaceae). Plant Syst. Evol. 2012, 298, 251–255. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Liu, L.; Wang, Y.; Fan, G.; Zhang, S.; Wang, Y.; Liao, K. Determination of genome size and chromosome ploidy of selected taxa from Prunus armeniaca by flow cytometry. Sci. Hortic. (Amsterdam) 2020, 261, 108987. [Google Scholar] [CrossRef]
- Šlenker, M.; Zozomová-Lihová, J.; Mandáková, T.; Kudoh, H.; Zhao, Y.; Soejima, A.; Yahara, T.; Skokanová, K.; Španiel, S.; Fls, K.M. Morphology and genome size of the widespread weed Cardamine occulta: How it differs from cleistogamic C. kokaiensis and other closely related taxa in Europe and Asia. Bot. J. Linn. Soc. 2018, 187, 456–482. [Google Scholar] [CrossRef]
- Pisupati, R.; Vergara, D.; Kane, N.C. Diversity and evolution of the repetitive genomic content in Cannabis sativa. BMC Genom. 2018, 19, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnable, P.S.; Ware, D.; Fulton, R.S.; Stein, J.C.; Wei, F.; Pasternak, S.; Liang, C.; Zhang, J.; Fulton, L.; Graves, T.A.; et al. The B73 maize genome: Complexity, diversity, and dynamics. Science 2009, 326, 1112–1115. [Google Scholar] [CrossRef] [PubMed]
- Quesneville, H. Twenty years of transposable element analysis in the Arabidopsis thaliana genome. Mob. DNA 2020, 11, 28. [Google Scholar] [CrossRef] [PubMed]
- Schmuths, H.; Meister, A.; Horres, R.; Bachmann, K. Genome size variation among accessions of Arabidopsis thaliana. Ann. Bot. 2004, 93, 317–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loureiro, J.; Rodriguez, E.; Doležel, J.; Santos, C. Flow cytometric and microscopic analysis of the effect of tannic acid on plant nuclei and estimation of DNA content. Ann. Bot. 2006, 98, 515–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, S.; Kaushik, S.; Raina, S.N. Estimation of nuclear DNA content and its variation among Indian Tea accessions by flow cytometry. Physiol. Mol. Biol. Plants 2019, 25, 339–346. [Google Scholar] [CrossRef]
- Noirot, M.; Barre, P.; Louarn, J.; Duperray, C.; Hamon, S. Nucleus-cytosol interactions—A source of stoichiometric error in flow cytometric estimation of nuclear DNA content in plants. Ann. Bot. 2000, 86, 309–316. [Google Scholar] [CrossRef] [Green Version]
- Noirot, M.; Barre, P.; Duperray, C.; Louarn, J.; Hamon, S. Effects of caffeine and chlorogenic acid on propidium iodide accessibility to DNA: Consequences on genome size evaluation in coffee tree. Ann. Bot. 2003, 92, 259–264. [Google Scholar] [CrossRef] [Green Version]
- Bennett, M.D.; Price, H.J.; Johnston, J.S. Anthocyanin inhibits propidium iodide DNA fluorescence in Euphorbia pulcherrima: Implications for genome size variation and flow cytometry. Ann. Bot. 2008, 101, 777–790. [Google Scholar] [CrossRef]
- Price, H.J.; Hodnett, G.; Johnston, J.S. Sunflower (Helianthus annuus) leaves contain compounds that reduce nuclear propidium iodide fluorescence. Ann. Bot. 2000, 86, 929–934. [Google Scholar] [CrossRef] [Green Version]
- Akagi, T.; Jung, K.; Masuda, K.; Shimizu, K.K. Polyploidy before and after domestication of crop species. Curr. Opin. Plant Biol. 2022, 69, 102255. [Google Scholar] [CrossRef]
- Salman-Minkov, A.; Sabath, N.; Mayrose, I. Whole-genome duplication as a key factor in crop domestication. Nat. Plants 2016, 2, 16115. [Google Scholar] [CrossRef]
- Peng, Y.; Yan, H.; Guo, L.; Deng, C.; Wang, C.; Wang, Y.; Kang, L.; Zhou, P.; Yu, K.; Dong, X.; et al. Reference genome assemblies reveal the origin and evolution of allohexaploid oat. Nat. Genet. 2022, 54, 1248–1258. [Google Scholar] [CrossRef]
- Peng, J.H.; Sun, D.; Nevo, E. Domestication evolution, genetics and genomics in wheat. Mol. Breed. 2011, 28, 281–301. [Google Scholar] [CrossRef]
- Yang, J.; Moeinzadeh, M.H.; Kuhl, H.; Helmuth, J.; Xiao, P.; Haas, S.; Liu, G.; Zheng, J.; Sun, Z.; Fan, W.; et al. Haplotype-resolved sweet potato genome traces back its hexaploidization history. Nat. Plants 2017, 3, 696–703. [Google Scholar] [CrossRef]
- Qi, X.; An, H.; Hall, T.E.; Di, C.; Blischak, P.D.; McKibben, M.T.W.; Hao, Y.; Conant, G.C.; Pires, J.C.; Barker, M.S. Genes derived from ancient polyploidy have higher genetic diversity and are associated with domestication in Brassica rapa. New Phytol. 2021, 230, 372–386. [Google Scholar] [CrossRef]
- Šimoníková, D.; Nĕmečková, A.; Čížková, J.; Brown, A.; Swennen, R.; Doležel, J.; Hřibová, E. Chromosome painting in cultivated bananas and their wild relatives (Musa spp.) reveals differences in chromosome structure. Int. J. Mol. Sci. 2020, 21, 7915. [Google Scholar] [CrossRef]
- Hesami, M.; Baiton, A.; Alizadeh, M.; Pepe, M.; Torkamaneh, D.; Phineas Jones, A.M. Advances and Perspectives in Tissue Culture and Genetic Engineering of Cannabis. Int. J. Mol. Sci. 2021, 22, 5671. [Google Scholar] [CrossRef]
- Romo, A.; Hidalgo, O.; Boratyński, A.; Sobierajska, K.; Jasińska, A.K.; Vallès, J.; Garnatje, T. Genome size and ploidy levels in highly fragmented habitats: The case of western Mediterranean Juniperus (Cupressaceae) with special emphasis on J. thurifera L. Tree Genet. Genomes 2013, 9, 587–599. [Google Scholar] [CrossRef]
- Marie, D.; Brown, S.C. A cytometric exercise in plant DNA histograms, with 2C values for 70 species. Biol. Cell 1993, 78, 41–51. [Google Scholar] [CrossRef]
- Vagera, J.; Paulíková, D.; Doležel, J. The Development of Male and Female Regenerants by In Vitro Androgenesis in Dioecious Plant Melandrium album. Ann. Bot. 1994, 73, 455–459. [Google Scholar] [CrossRef]
- Błocka-Wandas, M.; Sliwinska, E.; Grabowska-Joachimiak, A.; Musial, K.; Joachimiak, A.J. Male gametophyte development and two different DNA classes of pollen grains in Rumex acetosa L., a plant with an XX/XY1Y2 sex chromosome system and a female-biased sex ratio. Sex. Plant Reprod. 2007, 20, 171–180. [Google Scholar] [CrossRef]
- Grabowska-Joachimiak, A.; Kula, A.; Książczyk, T.; Chojnicka, J.; Sliwinska, E.; Joachimiak, A.J. Chromosome landmarks and autosome-sex chromosome translocations in Rumex hastatulus, a plant with XX/XY1Y2 sex chromosome system. Chromosom. Res. 2015, 23, 187–197. [Google Scholar] [CrossRef] [Green Version]
- Sousa, A.; Fuchs, J.; Renner, S.S. Molecular cytogenetics (FISH, GISH) of Coccinia grandis: A ca. 3 myr-old species of cucurbitaceae with the largest Y/autosome divergence in flowering plants. Cytogenet. Genome Res. 2013, 139, 107–118. [Google Scholar] [CrossRef] [Green Version]
- Charlesworth, D. Plant sex determination and sex chromosomes. Heredity (Edinb.) 2002, 88, 94–101. [Google Scholar] [CrossRef]
- Razumova, O.V.; Divashuk, M.G.; Alexandrov, O.S.; Karlov, G.I. GISH painting of the Y chromosomes suggests advanced phases of sex chromosome evolution in three dioecious Cannabaceae species (Humulus lupulus, H. japonicus, and Cannabis sativa). Protoplasma 2022. [Google Scholar] [CrossRef]
- Obermayer, R.; Leitch, I.J.; Hanson, L.; Bennett, M.D. Nuclear DNA C-values in 30 species double the familial representation in pteridophytes. Ann. Bot. 2002, 90, 209–217. [Google Scholar] [CrossRef] [Green Version]
- Doležel, J.; Binarová, P.; Lucretti, S. Analysis of Nuclear DNA content in plant cells by Flow cytometry. Biol. Plant. 1989, 31, 113–120. [Google Scholar] [CrossRef]
- Ebihara, A.; Ishikawa, H.; Matsumoto, S.; Lin, S.J.; Iwatsuki, K.; Takamiya, M.; Watano, Y.; Ito, M. Nuclear DNA, chloroplast DNA, and ploidy analysis clarified biological complexity of the Vandenboschia radicans complex (Hymenophyllaceae) in Japan and adjacent areas. Am. J. Bot. 2005, 92, 1535–1547. [Google Scholar] [CrossRef]
- Loureiro, J.; Rodriguez, E.; Doležel, J.; Santos, C. Two new nuclear isolation buffers for plant DNA flow cytometry: A test with 37 species. Ann. Bot. 2007, 100, 875–888. [Google Scholar] [CrossRef] [Green Version]
- Pellicer, J.; Powell, R.F.; Leitch, I.J. The Application of Flow Cytometry for Estimating Genome Size, Ploidy Level Endopolyploidy, and Reproductive Modes in Plants. In Molecular Plant Taxonomy: Methods and Protocols; Besse, P., Ed.; Springer: New York, NY, USA, 2021; pp. 325–361. ISBN 9781071609972. [Google Scholar]
- Doležel, J.; Greilhuber, J.; Suda, J. Estimation of nuclear DNA content in plants using flow cytometry. Nat. Protoc. 2007, 2, 2233–2244. [Google Scholar] [CrossRef]
- Doyle, J.; Doyle, J.L. Genomic plant DNA preparation from fresh tissue-CTAB method. Phytochem. Bull. 1987, 19, 1–15. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | No. ind. | DF | Sum Sq. | Mean Sq. | F Value | p Value |
|---|---|---|---|---|---|---|
| Accessions | 482 | |||||
| Accession | 83 | 0.5206 | 0.006272 | 3.386 | <0.001 | |
| Residuals | 398 | 0.7372 | 0.001852 | |||
| Distribution area | 482 | |||||
| Distribution area | 11 | 0.2185 | 0.019863 | 8.983 | <0.001 | |
| Residuals | 470 | 1.0393 | 0.002211 | |||
| Sex | 96 | |||||
| Sex | 1 | 0.0397 | 0.03965 | 11.62 | <0.001 | |
| Residuals | 94 | 0.3208 | 0.00341 |
| Female Genome Size (pg/2C) | Male Genome Size (pg/2C) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Accession | No. ind. | Mean | SD 1 | Min. | Max. | No. ind. | Mean | SD 1 | Min. | Max. | Difference |
| AM15 | 4 | 2.021 | 0.106 | 1.916 | 2.152 | 3 | 1.942 | 0.014 | 1.927 | 1.956 | 0.079 |
| AM3 | 3 | 1.927 | 0.013 | 1.913 | 1.937 | 1 | 1.989 | / | 1.989 | 1.989 | 0.061 |
| BG3 | 1 | 1.950 | / | 1.950 | 1.950 | 2 | 1.986 | 0.036 | 1.960 | 2.011 | 0.036 |
| CAM | 4 | 1.999 | 0.066 | 1.913 | 2.073 | 6 | 2.029 | 0.034 | 1.986 | 2.081 | 0.030 |
| HU11 | 3 | 1.913 | 0.017 | 1.902 | 1.932 | 2 | 1.923 | 0.000 | 1.922 | 1.923 | 0.010 |
| HU9 | 6 | 1.939 | 0.034 | 1.875 | 1.968 | 4 | 1.997 | 0.068 | 1.920 | 2.072 | 0.058 |
| IND2 | 2 | 2.035 | 0.057 | 1.995 | 2.075 | 6 | 2.036 | 0.030 | 1.980 | 2.067 | 0.001 |
| CUL7 | 5 | 2.003 | 0.027 | 1.973 | 2.040 | 2 | 1.989 | 0.030 | 1.968 | 2.011 | 0.014 |
| KAZ | 6 | 1.914 | 0.045 | 1.869 | 1.963 | 5 | 1.957 | 0.031 | 1.933 | 2.006 | 0.043 |
| MAR | 3 | 1.938 | 0.031 | 1.904 | 1.964 | 2 | 1.973 | 0.067 | 1.926 | 2.021 | 0.036 |
| MN3 | 2 | 1.927 | 0.028 | 1.908 | 1.947 | 5 | 1.997 | 0.080 | 1.922 | 2.118 | 0.069 |
| MN5 | 4 | 1.925 | 0.080 | 1.810 | 1.979 | 1 | 2.016 | / | 2.016 | 2.016 | 0.091 |
| RO2 | 2 | 1.882 | 0.032 | 1.859 | 1.905 | 0 | / | / | / | / | / |
| RO3 | 1 | 1.897 | / | 1.897 | 1.897 | 5 | 1.947 | 0.016 | 1.932 | 1.974 | 0.051 |
| RO5 | 4 | 2.021 | 0.106 | 1.916 | 2.152 | 3 | 1.942 | 0.014 | 1.927 | 1.956 | 0.079 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balant, M.; Rodríguez González, R.; Garcia, S.; Garnatje, T.; Pellicer, J.; Vallès, J.; Vitales, D.; Hidalgo, O. Novel Insights into the Nature of Intraspecific Genome Size Diversity in Cannabis sativa L. Plants 2022, 11, 2736. https://doi.org/10.3390/plants11202736
Balant M, Rodríguez González R, Garcia S, Garnatje T, Pellicer J, Vallès J, Vitales D, Hidalgo O. Novel Insights into the Nature of Intraspecific Genome Size Diversity in Cannabis sativa L. Plants. 2022; 11(20):2736. https://doi.org/10.3390/plants11202736
Chicago/Turabian StyleBalant, Manica, Roi Rodríguez González, Sònia Garcia, Teresa Garnatje, Jaume Pellicer, Joan Vallès, Daniel Vitales, and Oriane Hidalgo. 2022. "Novel Insights into the Nature of Intraspecific Genome Size Diversity in Cannabis sativa L." Plants 11, no. 20: 2736. https://doi.org/10.3390/plants11202736
APA StyleBalant, M., Rodríguez González, R., Garcia, S., Garnatje, T., Pellicer, J., Vallès, J., Vitales, D., & Hidalgo, O. (2022). Novel Insights into the Nature of Intraspecific Genome Size Diversity in Cannabis sativa L. Plants, 11(20), 2736. https://doi.org/10.3390/plants11202736