

Detection of Superior Rice Genotypes and Yield Stability under Different Nitrogen Levels Using AMMI Model and Stability Statistics
 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Genotypic Variability of GY under Different N Environments
2.2. Combined AMMI Analysis of Variance of the GY and the Decomposition of GEI Effect
2.3. Environmental Effect on the Performance of the Genotypes
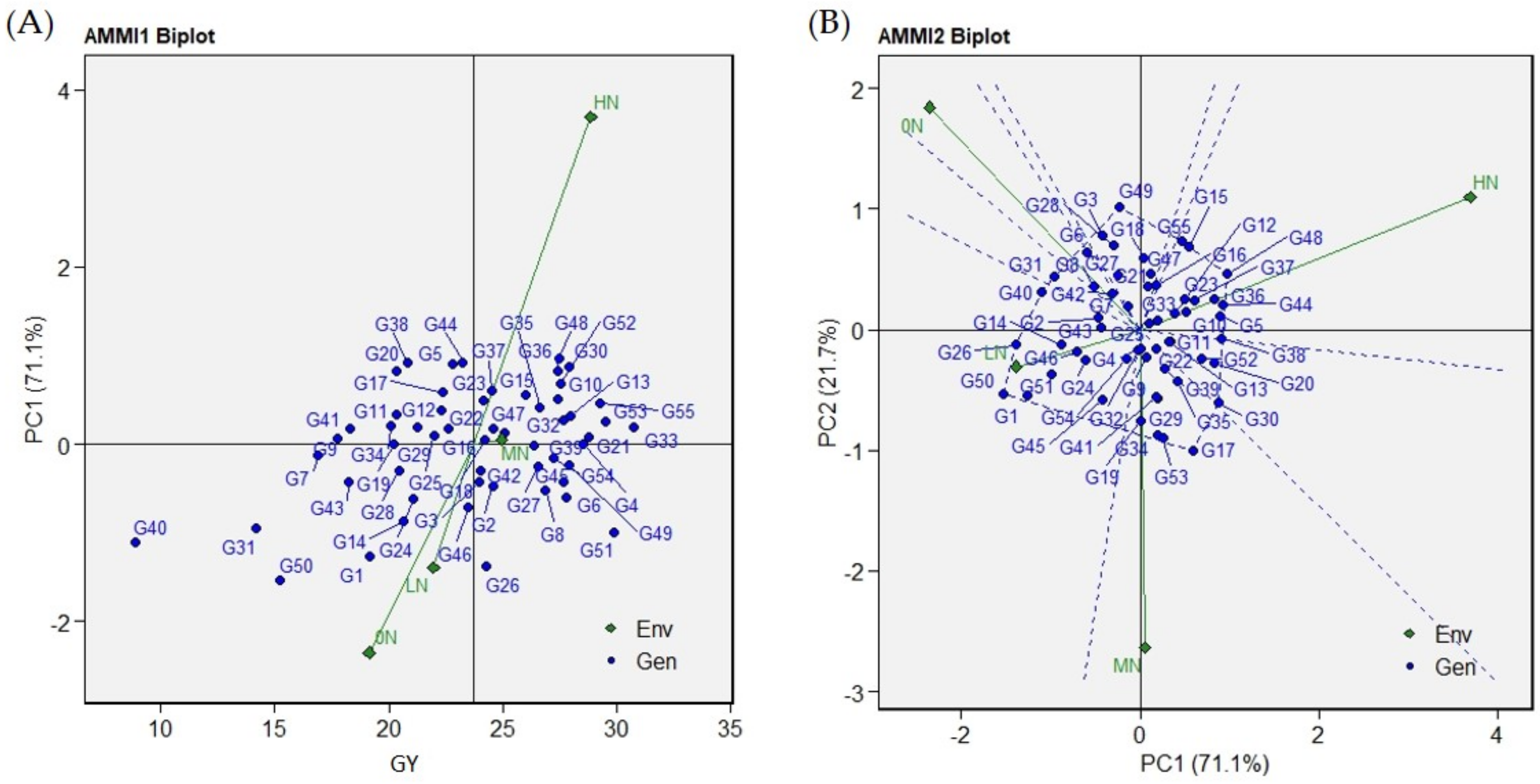
2.4. Graphical Representation of Genotypes and N-Environments in the AMMI Biplots
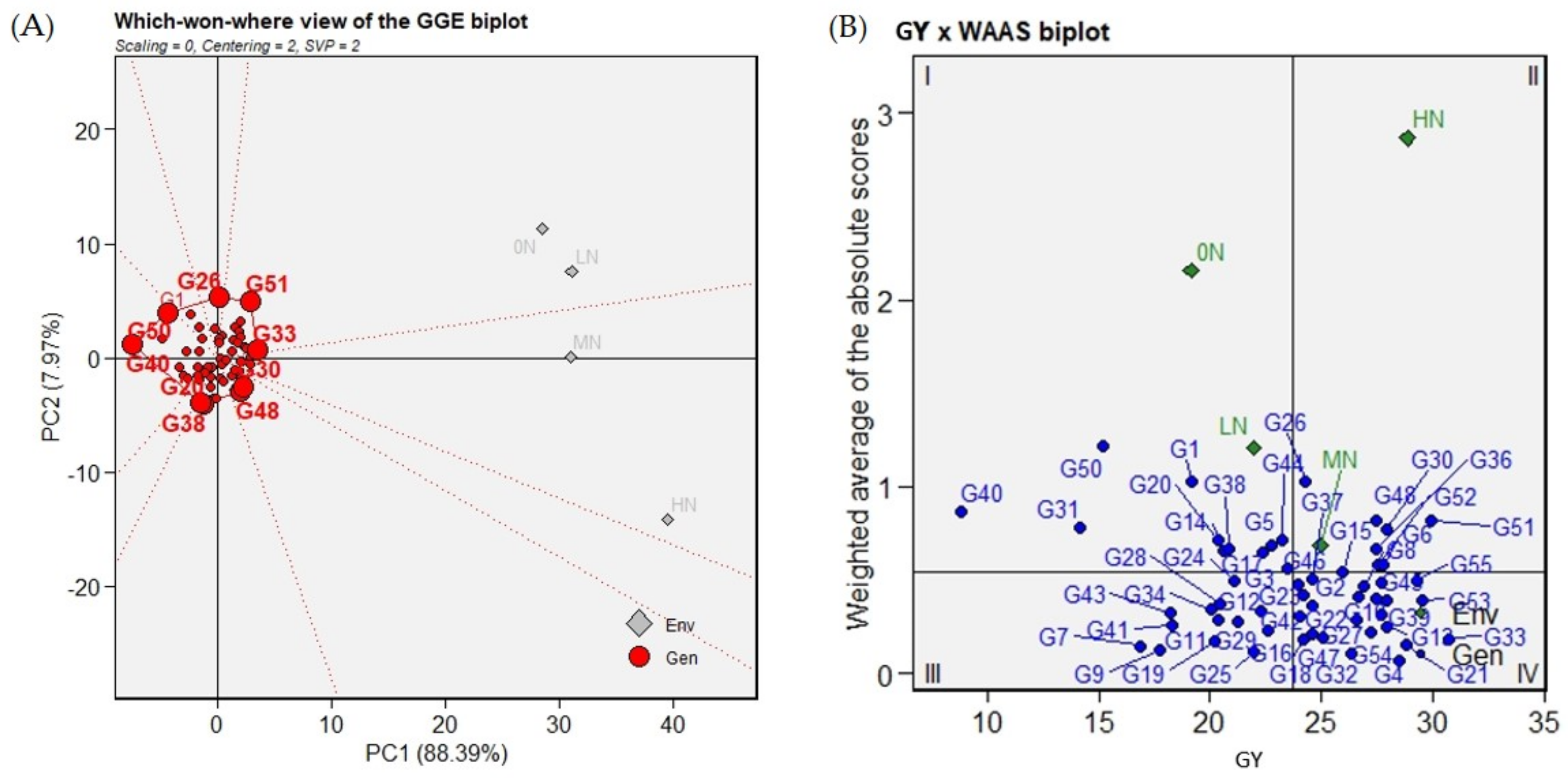
2.5. Which-Won-Where GGE Biplot Analysis
2.6. Superior Genotypes Selection Based on GY Means and Stability Parameters
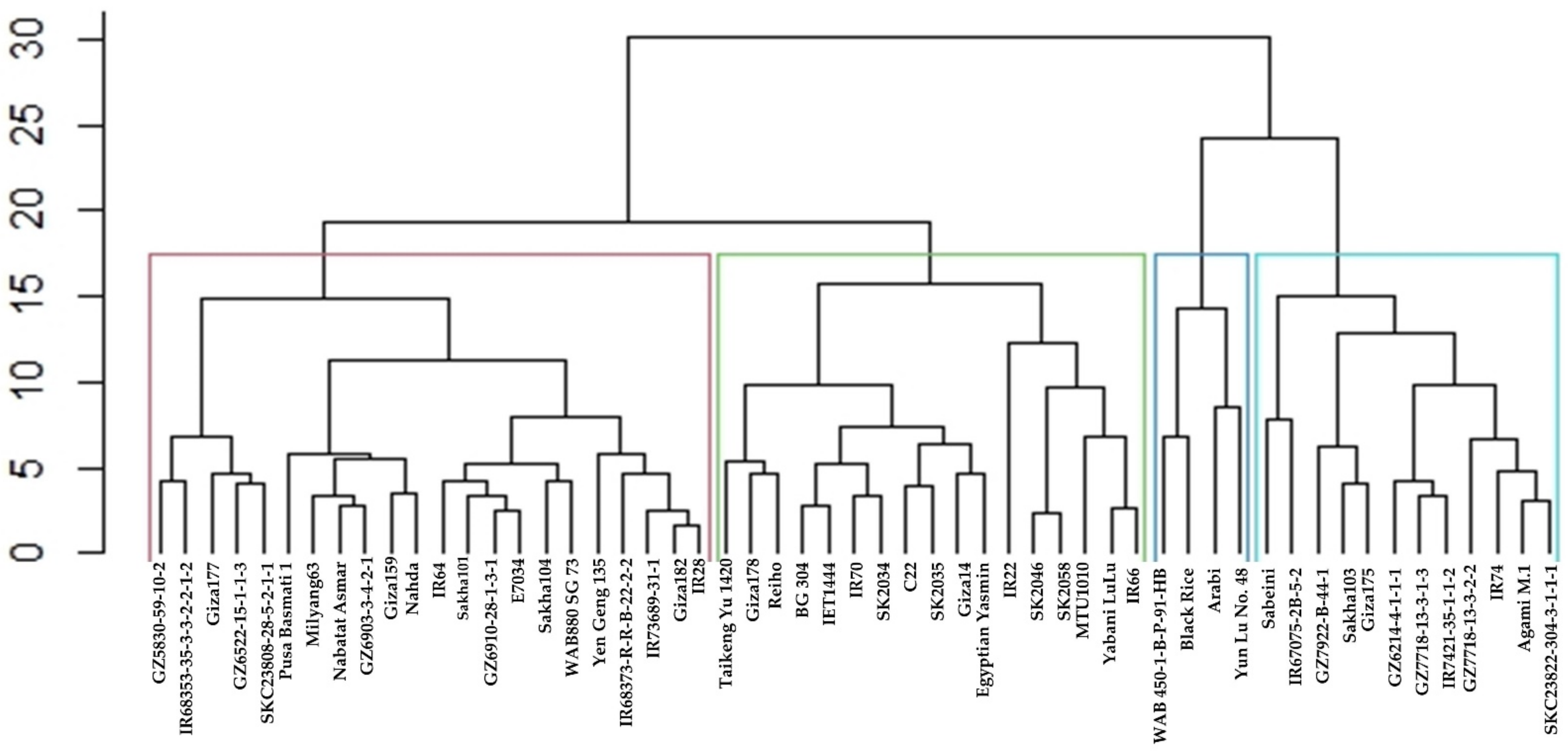
2.7. Cluster Analysis and Dendrogram Based on the Stability Statistics Values
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Experimental Location and Soil Properties
4.3. Field Experiments and Environmental Conditions
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shaheen, S.M.; Antoniadis, V.; Shahid, M.; Yang, Y.; Abdelrahman, H.; Zhang, T.; Hassan, N.E.; Bibi, I.; Niazi, N.K.; Younis, S.A. Sustainable applications of rice feedstock in agro-environmental and construction sectors: A global perspective. Renew. Sustain. Energy Rev. 2022, 153, 111791. [Google Scholar] [CrossRef]
- Naeem, E.; Abd El-Megeed, T.; Emadeldin, Y.; Abushady, A.M.; Abdelrahman, M. Injected Anhydrous Ammonia Is More Effective Than Broadcast Urea as a Source of Nitrogen for Drill Seeded Rice. Agronomy 2022, 12, 942. [Google Scholar] [CrossRef]
- Fageria, N. Yield physiology of rice. J. Plant Nutr. 2007, 30, 843–879. [Google Scholar] [CrossRef]
- Abdel-Rahman, M.; El-Denary, M.; Ammar, M.; Abdelkhalik, A.; Draz, A.; Dora, S. QTL validation for grain yield and nitrogen use efficiency under different nitrogen levels in rice. Egypt. J. Genet. Cytol. 2015, 44, 235–251. [Google Scholar] [CrossRef] [Green Version]
- Ju, X.-T.; Xing, G.-X.; Chen, X.-P.; Zhang, S.-L.; Zhang, L.-J.; Liu, X.-J.; Cui, Z.-L.; Yin, B.; Christie, P.; Zhu, Z.-L. Reducing environmental risk by improving N management in intensive Chinese agricultural systems. Proc. Natl. Acad. Sci. USA 2009, 106, 3041–3046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnamurthy, S.; Sharma, P.; Sharma, D.; Ravikiran, K.; Singh, Y.; Mishra, V.; Burman, D.; Maji, B.; Mandal, S.; Sarangi, S. Identification of mega-environments and rice genotypes for general and specific adaptation to saline and alkaline stresses in India. Sci. Rep. 2017, 7, 7968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oladosu, Y.; Rafii, M.Y.; Abdullah, N.; Magaji, U.; Miah, G.; Hussin, G.; Ramli, A. Genotype× Environment interaction and stability analyses of yield and yield components of established and mutant rice genotypes tested in multiple locations in Malaysia. Acta Agric. Scand. Sect. B—Soil Plant Sci. 2017, 67, 590–606. [Google Scholar] [CrossRef]
- Mehana, M.; Abdelrahman, M.; Emadeldin, Y.; Rohila, J.S.; Karthikeyan, R. Impact of genetic improvements of rice on its water use and effects of climate variability in Egypt. Agriculture 2021, 11, 865. [Google Scholar] [CrossRef]
- Bocianowski, J.; Tratwal, A.; Nowosad, K. Genotype by environment interaction for main winter triticale varieties characteristics at two levels of technology using additive main effects and multiplicative interaction model. Euphytica 2021, 217, 26. [Google Scholar] [CrossRef]
- Chairi, F.; Aparicio, N.; Serret, M.D.; Araus, J.L. Breeding effects on the genotype× environment interaction for yield of durum wheat grown after the Green Revolution: The case of Spain. Crop J. 2020, 8, 623–634. [Google Scholar] [CrossRef]
- Khan, M.M.H.; Rafii, M.Y.; Ramlee, S.I.; Jusoh, M.; Al Mamun, M. AMMI and GGE biplot analysis for yield performance and stability assessment of selected Bambara groundnut (Vigna subterranea L. Verdc.) genotypes under the multi-environmental trials (METs). Sci. Rep. 2021, 11, 22791. [Google Scholar] [CrossRef]
- Omrani, A.; Omrani, S.; Khodarahmi, M.; Shojaei, S.H.; Illés, Á.; Bojtor, C.; Mousavi, S.M.N.; Nagy, J. Evaluation of Grain Yield Stability in Some Selected Wheat Genotypes Using AMMI and GGE Biplot Methods. Agronomy 2022, 12, 1130. [Google Scholar] [CrossRef]
- Yohane, E.N.; Shimelis, H.; Laing, M.; Mathew, I.; Shayanowako, A. Genotype-by-environment interaction and stability analyses of grain yield in pigeonpea [Cajanus cajan (L.) Millspaugh]. Acta Agric. Scand. Sect. B—Soil Plant Sci. 2021, 71, 145–155. [Google Scholar] [CrossRef]
- Yue, H.; Gauch, H.G.; Wei, J.; Xie, J.; Chen, S.; Peng, H.; Bu, J.; Jiang, X. Genotype by Environment Interaction Analysis for Grain Yield and Yield Components of Summer Maize Hybrids across the Huanghuaihai Region in China. Agriculture 2022, 12, 602. [Google Scholar] [CrossRef]
- Abdelrahman, M.; Selim, M.E.; ElSayed, M.A.; Ammar, M.H.; Hussein, F.A.; ElKholy, N.K.; ElShamey, E.A.; Khan, N.; Attia, K.A. Developing Novel Rice Genotypes Harboring Specific QTL Alleles Associated with High Grain Yield under Water Shortage Stress. Plants 2021, 10, 2219. [Google Scholar] [CrossRef]
- Ferreira Coelho, I.; Peixoto, M.A.; Marcal, T.d.S.; Bernardeli, A.; Silva Alves, R.; de Lima, R.O.; Reis, E.F.d.; Bhering, L.L. Accounting for spatial trends in multi-environment diallel analysis in maize breeding. PLoS ONE 2021, 16, e0258473. [Google Scholar] [CrossRef]
- LI, C.-x.; Yun, S.; ZHANG, L.-l. Effects of long-term organic fertilization on soil microbiologic characteristics, yield and sustainable production of winter wheat. J. Integr. Agric. 2018, 17, 210–219. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Yuan, J.; Zhang, M.; Zhang, Y.; Wang, L.; Li, J. Long term effects of crop rotation and fertilization on crop yield stability in southeast China. Sci. Rep. 2022, 12, 14234. [Google Scholar] [CrossRef]
- Wricke, G. Uber eine Methode zur Erfassung der okologischen Streubreite in Feldverzuchen. Z. Pflanz. 1962, 47, 92–96. [Google Scholar]
- Shukla, G. Some statistical aspects of partitioning genotype-environmental components of variability. Heredity 1972, 29, 237–245. [Google Scholar] [CrossRef]
- Francis, T.; Kannenberg, L. Yield stability studies in short-season maize. I. A descriptive method for grouping genotypes. Can. J. Plant Sci. 1978, 58, 1029–1034. [Google Scholar] [CrossRef]
- Roemer, J. Sinde die ertagdreichen Sorten ertagissicherer. Mitt DLG 1917, 32, 87–89. [Google Scholar]
- Plaisted, R.; Peterson, L. A technique for evaluating the ability of selections to yield consistently in different locations or seasons. Am. Potato J. 1959, 36, 381–385. [Google Scholar] [CrossRef]
- Plaisted, R. A shorter method for evaluating the ability of selections to yield consistently over locations. Am. Potato J. 1960, 37, 166–172. [Google Scholar] [CrossRef]
- Finlay, K.W.; Wilkinson, G. The analysis of adaptation in a plant-breeding programme. Aust. J. Agric. Res. 1963, 14, 742–754. [Google Scholar] [CrossRef] [Green Version]
- Zobel, R.W.; Wright, M.J.; Gauch, H.G., Jr. Statistical analysis of a yield trial. Agron. J. 1988, 80, 388–393. [Google Scholar] [CrossRef]
- Yan, W.; Cornelius, P.L.; Crossa, J.; Hunt, L. Two types of GGE biplots for analyzing multi-environment trial data. Crop Sci. 2001, 41, 656–663. [Google Scholar] [CrossRef] [Green Version]
- Agahi, K.; Ahmadi, J.; Oghan, H.A.; Fotokian, M.H.; Orang, S.F. Analysis of genotype× environment interaction for seed yield in spring oilseed rape using the AMMI model. Crop Breed. Appl. Biotechnol. 2020, 20, e26502012. [Google Scholar] [CrossRef]
- Katsenios, N.; Sparangis, P.; Leonidakis, D.; Katsaros, G.; Kakabouki, I.; Vlachakis, D.; Efthimiadou, A. Effect of genotype× Environment interaction on yield of maize Hybrids in Greece Using AMMI Analysis. Agronomy 2021, 11, 479. [Google Scholar] [CrossRef]
- Mafouasson, H.N.A.; Gracen, V.; Yeboah, M.A.; Ntsomboh-Ntsefong, G.; Tandzi, L.N.; Mutengwa, C.S. Genotype-by-environment interaction and yield stability of maize single cross hybrids developed from tropical inbred lines. Agronomy 2018, 8, 62. [Google Scholar] [CrossRef] [Green Version]
- Gauch, H.G., Jr.; Piepho, H.P.; Annicchiarico, P. Statistical analysis of yield trials by AMMI and GGE: Further considerations. Crop Sci. 2008, 48, 866–889. [Google Scholar] [CrossRef]
- Nassar, R.; Huehn, M. Studies on estimation of phenotypic stability: Tests of significance for nonparametric measures of phenotypic stability. Biometrics 1987, 43, 45–53. [Google Scholar] [CrossRef]
- Kang, M. A rank-sum method for selecting high-yielding, stable corn genotypes. Cereal Res. Commun. 1988, 16, 113–115. [Google Scholar]
- Fox, P.; Skovmand, B.; Thompson, B.; Braun, H.-J.; Cormier, R. Yield and adaptation of hexaploid spring triticale. Euphytica 1990, 47, 57–64. [Google Scholar] [CrossRef]
- Thennarasu, K. On Certain Non-Parametric Procedures for Studying Genotype-Environment Inertactions and Yield Stability; IARI, Division of Agricultural Statistics: New Delhi, India, 1995. [Google Scholar]
- Farshadfar, E. Incorporation of AMMI stability value and grain yield in a single non-parametric index (GSI) in bread wheat. Pak. J. Biol. Sci. 2008, 11, 1791. [Google Scholar] [CrossRef] [Green Version]
- Olivoto, T.; Lúcio, A.D.C. metan: An R package for multi-environment trial analysis. Methods Ecol. Evol. 2020, 11, 783–789. [Google Scholar] [CrossRef] [Green Version]
- Mastrodomenico, A.T.; Haegele, J.W.; Seebauer, J.R.; Below, F.E. Yield stability differs in commercial maize hybrids in response to changes in plant density, nitrogen fertility, and environment. Crop Sci. 2018, 58, 230–241. [Google Scholar] [CrossRef] [Green Version]
- Stomph, T.; Dordas, C.; Baranger, A.; de Rijk, J.; Dong, B.; Evers, J.; Gu, C.; Li, L.; Simon, J.; Jensen, E.S. Designing intercrops for high yield, yield stability and efficient use of resources: Are there principles? Adv. Agron. 2020, 160, 1–50. [Google Scholar]
- Zewdu, Z.; Abebe, T.; Mitiku, T.; Worede, F.; Dessie, A.; Berie, A.; Atnaf, M. Performance evaluation and yield stability of upland rice (Oryza sativa L.) varieties in Ethiopia. Cogent Food Agric. 2020, 6, 1842679. [Google Scholar] [CrossRef]
- Zaid, I.U.; Zahra, N.; Habib, M.; Naeem, M.K.; Asghar, U.; Uzair, M.; Latif, A.; Rehman, A.; Ali, G.M.; Khan, M.R. Estimation of Genetic Variances and Stability Components of Yield-Related Traits of Green Super Rice at Multi-Environmental Conditions in Pakistan. Agronomy 2022, 12, 1157. [Google Scholar] [CrossRef]
- Gauch, H., Jr.; Zobel, R.W. Optimal replication in selection experiments. Crop Sci. 1996, 36, 838–843. [Google Scholar] [CrossRef]
- George, N.; Lundy, M. Quantifying genotype× environment effects in long-term common wheat yield trials from an agroecologically diverse production region. Crop Sci. 2019, 59, 1960–1972. [Google Scholar] [CrossRef]
- Gauch, H., Jr. A simple protocol for AMMI analysis of yield trials. Crop Sci. 2013, 53, 1860–1869. [Google Scholar] [CrossRef]
- Pour-Aboughadareh, A.; Barati, A.; Koohkan, S.A.; Jabari, M.; Marzoghian, A.; Gholipoor, A.; Shahbazi-Homonloo, K.; Zali, H.; Poodineh, O.; Kheirgo, M. Dissection of genotype-by-environment interaction and yield stability analysis in barley using AMMI model and stability statistics. Bull. Natl. Res. Cent. 2022, 46, 19. [Google Scholar] [CrossRef]
- Da Silva, K.J.; Teodoro, P.E.; da Silva, M.J.; Teodoro, L.P.R.; Cardoso, M.J.; Godinho, V.d.P.C.; Mota, J.H.; Simon, G.A.; Tardin, F.D.; da Silva, A.R. Identification of mega-environments for grain sorghum in Brazil using GGE biplot methodology. Agron. J. 2021, 113, 3019–3030. [Google Scholar] [CrossRef]
- Bakare, M.A.; Kayondo, S.I.; Aghogho, C.I.; Wolfe, M.D.; Parkes, E.Y.; Kulakow, P.; Egesi, C.; Rabbi, I.Y.; Jannink, J.-L. Exploring genotype by environment interaction on cassava yield and yield related traits using classical statistical methods. PLoS ONE 2022, 17, e0268189. [Google Scholar] [CrossRef]
- Omar, M.; Rabie, H.A.; Mowafi, S.A.; Othman, H.T.; El-Moneim, D.A.; Alharbi, K.; Mansour, E.; Ali, M. Multivariate Analysis of Agronomic Traits in Newly Developed Maize Hybrids Grown under Different Agro-Environments. Plants 2022, 11, 1187. [Google Scholar] [CrossRef]
- Anila, M.; Mahadeva Swamy, H.; Kale, R.; Bhadana, V.; Anantha, M.; Hajira, S.; Balachiranjeevi, C.; Ayyappa Dass, M.; Bhaskar, S.; Dilip, T. Breeding lines of the Indian mega-rice variety, MTU 1010, possessing protein kinase OsPSTOL (Pup1), show better root system architecture and higher yield in soils with low phosphorus. Mol. Breed. 2018, 38, 147. [Google Scholar] [CrossRef]
- Koutroubas, S.; Ntanos, D. Genotypic differences for grain yield and nitrogen utilization in Indica and Japonica rice under Mediterranean conditions. Field Crops Res. 2003, 83, 251–260. [Google Scholar] [CrossRef]
- Olivoto, T.; Lúcio, A.D.; da Silva, J.A.; Marchioro, V.S.; de Souza, V.Q.; Jost, E. Mean performance and stability in multi-environment trials I: Combining features of AMMI and BLUP techniques. Agron. J. 2019, 111, 2949–2960. [Google Scholar] [CrossRef]
- Ahakpaz, F.; Abdi, H.; Neyestani, E.; Hesami, A.; Mohammadi, B.; Mahmoudi, K.N.; Abedi-Asl, G.; Noshabadi, M.R.J.; Ahakpaz, F.; Alipour, H. Genotype-by-environment interaction analysis for grain yield of barley genotypes under dryland conditions and the role of monthly rainfall. Agric. Water Manag. 2021, 245, 106665. [Google Scholar] [CrossRef]
- Pour-Aboughadareh, A.; Sanjani, S.; Nikkhah-Chamanabad, H.; Mehrvar, M.R.; Asadi, A.; Amini, A. Identification of salt-tolerant barley genotypes using multiple-traits index and yield performance at the early growth and maturity stages. Bull. Natl. Res. Cent. 2021, 45, 117. [Google Scholar] [CrossRef]
- Piper, C. Soil and Plant Analysis; The University of Adelaide: Adelaide, Australia, 1950; Volume 190, p. 4. [Google Scholar]
- Black, C.A. Method of soil analysis part 2. Chem. Microbiol. Prop. 1965, 9, 1387–1388. [Google Scholar]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2012. [Google Scholar]
- Yan, W.; Kang, M.S. GGE Biplot Analysis: A Graphical Tool for Breeders, Geneticists, and Agronomists; CRC Press: Boca Raton, FL, USA, 2002. [Google Scholar]
- Charrad, M.; Ghazzali, N.; Boiteau, V.; Niknafs, A. NbClust: An R package for determining the relevant number of clusters in a data set. J. Stat. Softw. 2014, 61, 1–36. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| E | Mean | EPC1 | Index | Class | 1 | 2 | 3 | 4 | 5 |
|---|---|---|---|---|---|---|---|---|---|
| 0N | 19.13842 | −2.35395 | −4.575242 | Unfavorable | IR22 | GIZA14 | C22 | MTU1010 | SK2058 |
| LN | 21.94194 | −1.39131 | −1.771727 | Unfavorable | IR22 | MTU1010 | Yabani Lulu | SK2035 | IR66 |
| MN | 24.93612 | 0.054521 | 1.222455 | favorable | SK2046 | IR22 | MTU1010 | Egyptian Yasmin | Giza178 |
| HN | 28.83818 | 3.690741 | 5.124515 | favorable | SK2058 | MTU1010 | Reiho | WAB 880 SG 73 | Giza178 |
| No. | Genotype | Type | Parentage | Origin |
|---|---|---|---|---|
| 1 | Sabieny | J | Selection from Introductions | EGYPT |
| 2 | Nabatat Asmar | J | Selection from Agami M1 | EGYPT |
| 3 | Giza 159 | J | Giza14/Agami M.1 | EGYPT |
| 4 | Yabani LuLu | J | Selection from Introductions | EGYPT |
| 5 | GZ 5830-59-10-2 | J | GZ4120/Suweon349 | EGYPT |
| 6 | Giza 14 | J | Yabani Pearl/Iraki16 | EGYPT |
| 7 | GZ 7718-13-3-1-3 | J | Sakha101/HR4856-1-1-2 | EGYPT |
| 8 | Nahda | J | Selection from Introductions | EGYPT |
| 9 | GZ 6214-4-1-1-1 | J | GZ4122-23-4-2/IRI396 | EGYPT |
| 10 | Sakha 101 | J | Giza176/Milyang79 | EGYPT |
| 11 | IR 68373-R-R-B-22-2-2 | T.J. | JINMIBYEO/YR14987-91 | IRRI |
| 12 | Giza 182 | I | Giza181/IR39422//Giza181 | EGYPT |
| 13 | GZ 7922-B-44-1 | J | Giza177/IDSA | EGYPT |
| 14 | Pusa Basmati 1 | I | India selection | INDIA |
| 15 | Sakha 104 | J | GZ4096/GZ4100 | EGYPT |
| 16 | IR 28 | I | IR8333-6-2-1///IR1561-149-1//IR24*4/O. NIVARA | IRRI |
| 17 | GZ 6522-15-1-1-3 | J | GZ5581/GZ4316 | EGYPT |
| 18 | IR 64 | I | IR5657-33-2-1/IR2061-4665-1-5-5 | IRRI |
| 19 | GZ 7718-13-3-2-2 | J | Sakha101/HR4856-1-1-2 | EGYPT |
| 20 | Giza 177 | J | Giza171/Yamji No.1//PI NO.4 | EGYPT |
| 21 | IR 66 | I | IR13240-108-2-2-3/IR9129-209-2-2-2-1 | IRRI |
| 22 | GZ 6910-28-1-3-1 | J | Sakha101/GZ24316(MUT) | EGYPT |
| 23 | IR 70 | I | IR19660-73-4/IR54//IR9828-36-3 | IRRI |
| 24 | Agami M.1 | J | Selection from cultivated varieties | EGYPT |
| 25 | Sakha 103 | J | GZ4120/Suweon349 | EGYPT |
| 26 | Arabi | I/J | Java3/Yabani Montkhab 3 | EGYPT |
| 27 | Milyang 63 | I/J | TONGIL/IR946-33-2-2-2//YR675-131-2 | KOREA |
| 28 | Yen Geng 135 | J | Chinese selection | CHINA |
| 29 | IR 73689-31-1 | T.J. | SR18977-TB-4/JINMIBYEO | IRRI |
| 30 | Giza 178 | I/J | Giza175/Milyang49 | EGYPT |
| 31 | WAB 450-1-B-P-91-HB | I | --- | Africa Rice |
| 32 | BG 304 | I | --- | SRILANKA |
| 33 | MTU 1010 | I | --- | INDIA |
| 34 | IR 68353-35-3-3-2-2-1-2 | T.J. | CHEOLWEON49/KYWHA9 | IRRI |
| 35 | Giza 175 | I/J | (IR28/IR1541)/(Giza180/Giza14) | EGYPT |
| 36 | WAB 880 SG 73 | I | --- | Africa Rice |
| 37 | E 7034 | J | EWAN NO.5/857 | CHINA |
| 38 | SKC 23808-28-5-2-1-1 | I/J | 98-Y-116/Sakha102 | IRRI |
| 39 | IET 1444 | I | TN1/CO.29 | INDIA |
| 40 | Black Rice | J | Jingo9601 | China |
| 41 | IR 7421-35-1-1-2 | T.J. | IR2035-290-2-1-1/MASINO | IRRI |
| 42 | GZ 6903-3-4-2-1 | J | Sakha101/Suweon313 | EGYPT |
| 43 | SKC 23822-304-3-1-1-1 | I/J | M202/Giza177 | IRRI |
| 44 | Taikeng Yu 1420 | J | C253///J692130/BL6//TAINUNG67/IR4547-2-1-2 | TAIWAN |
| 45 | Egyptian Yasmine | I | IR262-43-8-11/KDML105 | EGYPT |
| 46 | IR 67075-2B-5-2 | I | IR10198-66-2//GZ2175/CSR1 | IRRI |
| 47 | IR 74 | I | IR19661-131-1-2/IR15795-199-3-3 | IRRI |
| 48 | Reiho | J | HOYOKU/AYANISHKI | JAPAN |
| 49 | C 22 | I | TJRERMAS/BPI76//PALAWAN/AZUCENA | IRRI |
| 50 | Yun Lu No. 48 | J | LUYIN NO.7/YUNANJINGDAO-38 | CHINA |
| 51 | IR 22 | I | PETA/DEE GEO WOO GEN//TADUKAN | IRRI |
| 52 | SK2034 | I | IR69625A/Giza178R | EGYPT |
| 53 | SK2046 | I | IR69625A/Giza181R | EGYPT |
| 54 | SK2035 | I | IR70368A/Giza178R | EGYPT |
| 55 | SK2058 | I | IR69625A/Giza182R | EGYPT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdelrahman, M.; Alharbi, K.; El-Denary, M.E.; Abd El-Megeed, T.; Naeem, E.-S.; Monir, S.; Al-Shaye, N.A.; Ammar, M.H.; Attia, K.; Dora, S.A.; et al. Detection of Superior Rice Genotypes and Yield Stability under Different Nitrogen Levels Using AMMI Model and Stability Statistics. Plants 2022, 11, 2775. https://doi.org/10.3390/plants11202775
Abdelrahman M, Alharbi K, El-Denary ME, Abd El-Megeed T, Naeem E-S, Monir S, Al-Shaye NA, Ammar MH, Attia K, Dora SA, et al. Detection of Superior Rice Genotypes and Yield Stability under Different Nitrogen Levels Using AMMI Model and Stability Statistics. Plants. 2022; 11(20):2775. https://doi.org/10.3390/plants11202775
Chicago/Turabian StyleAbdelrahman, Mohamed, Khadiga Alharbi, Medhat E. El-Denary, Taher Abd El-Megeed, El-Sayed Naeem, Samah Monir, Najla A. Al-Shaye, Megahed H. Ammar, Kotb Attia, Said A. Dora, and et al. 2022. "Detection of Superior Rice Genotypes and Yield Stability under Different Nitrogen Levels Using AMMI Model and Stability Statistics" Plants 11, no. 20: 2775. https://doi.org/10.3390/plants11202775
APA StyleAbdelrahman, M., Alharbi, K., El-Denary, M. E., Abd El-Megeed, T., Naeem, E. -S., Monir, S., Al-Shaye, N. A., Ammar, M. H., Attia, K., Dora, S. A., & Draz, A. -S. E. (2022). Detection of Superior Rice Genotypes and Yield Stability under Different Nitrogen Levels Using AMMI Model and Stability Statistics. Plants, 11(20), 2775. https://doi.org/10.3390/plants11202775