
Chemical Composition and Cytotoxic Activity of Extracts from Carpesium divaricatum: In Vitro- versus Field-Grown Plants


Abstract
:1. Introduction
2. Results
2.1. In Vitro Culture
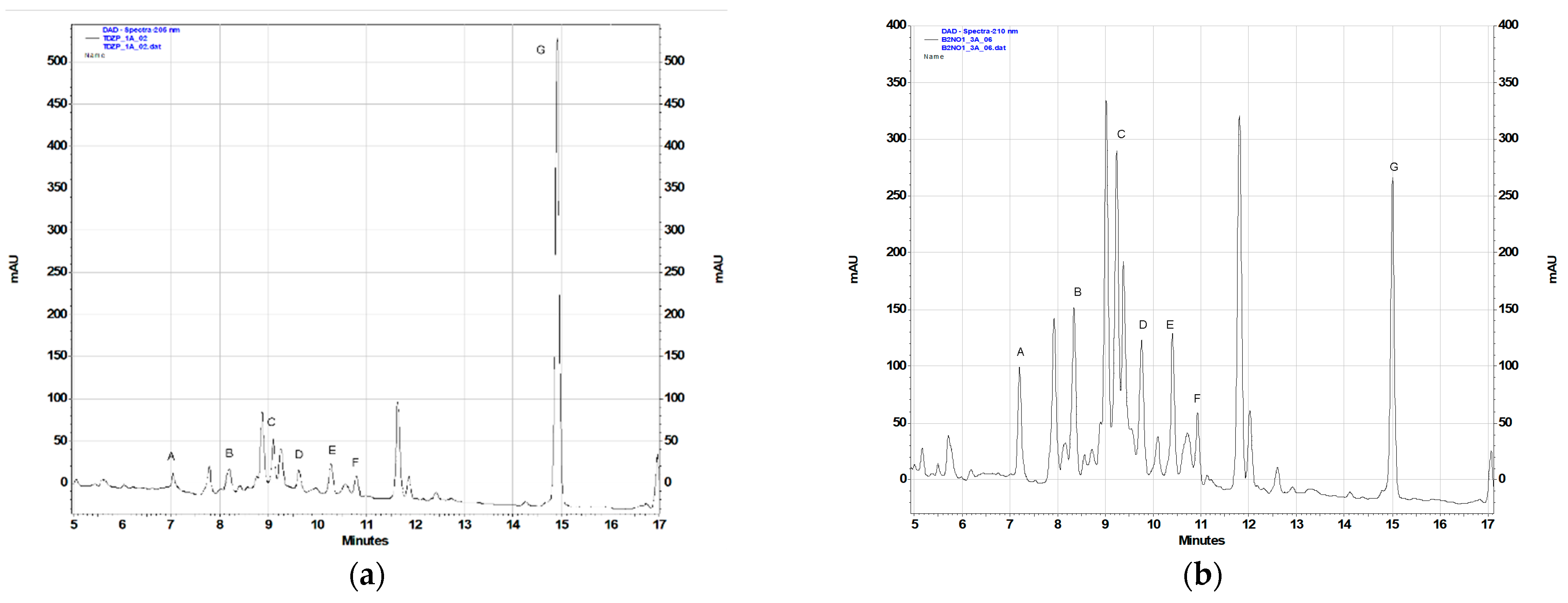
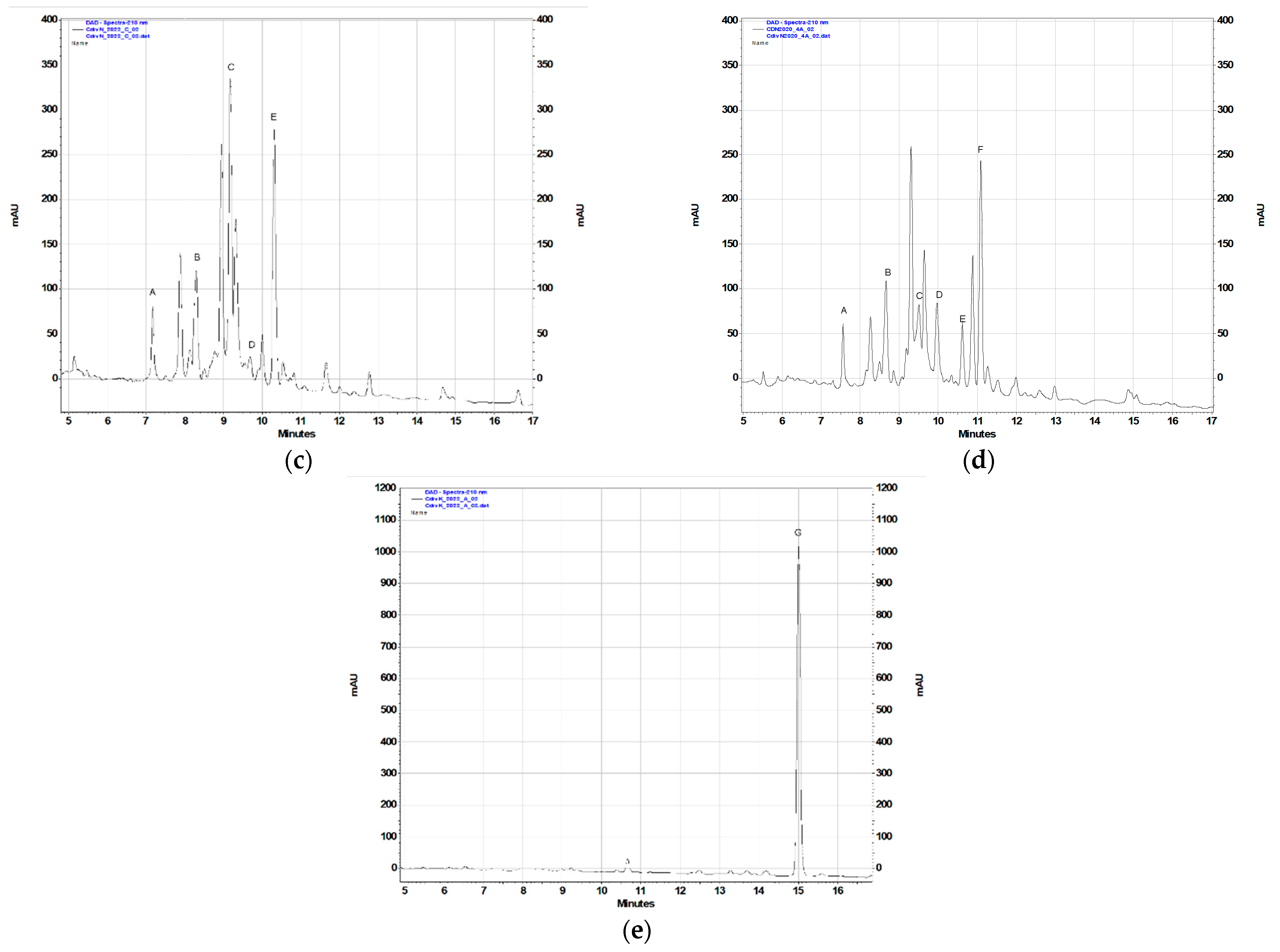
2.2. Phytochemical Analysis
2.3. Cytotoxic Activity of Extracts from In Vitro- and Field-Grown C. divaricatum
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Plant Material
4.2.1. Field-Grown Plants
4.2.2. In Vitro Culture
4.3. Extraction and Isolation of Sesquiterpene Lactones
4.4. RP-HPLC-DAD Analysis of Terpenoids in Plant Extracts
4.4.1. Sample Preparation
4.4.2. Semi-Quantitative Assessment of 10-Isobutyryloxy-8,9-epoxythymyl Isobutyrate and Sesquiterpene Lactone Content
4.5. Cell Culture and Cytotoxicity Assessment
4.5.1. Preparation of Plant Extracts
4.5.2. Assessment of Cytotoxic Activity
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- WFO. Carpesium L. Published on the Internet. 2022. Available online: http://www.worldfloraonline.org/taxon/wfo-4000006771 (accessed on 13 September 2022).
- Flora of China @ eFloras 2008. Carpesium Linnaeus. Published on the Internet. Available online: http://www.efloras.org/florataxon.aspx?flora_id=2&taxon_id=105695 (accessed on 13 September 2022).
- Zhang, J.-P.; Wang, G.-W.; Tian, X.-H.; Yang, Y.-X.; Liu, Q.-X.; Chen, L.-P.; Li, H.-L.; Zhang, W.-D. The genus Carpesium: A review of its ethnopharmacology, phytochemistry and pharmacology. J. Ethnopharmacol. 2015, 163, 173–191. [Google Scholar] [CrossRef] [PubMed]
- Shi, N.-N.; Hou, C.-C.; Liu, Y.; Li, K.-Y.; Mi, S.-D.; Tong, B.-L.; Zhang, M.-L. Chemical constituents of plants from the genus Carpesium. Heterocycl. Comm. 2022, 28, 95–123. [Google Scholar] [CrossRef]
- Gao, X.; Lin, C.-J.; Jia, Z.-J. Cytotoxic germacranolides and acyclic diterpenoids from the seeds of Carpesium triste. J. Nat. Prod. 2007, 70, 830–834. [Google Scholar] [CrossRef]
- Xu, D.-D.; Yan, Y.; Jiang, C.-X.; Liang, J.-J.; Li, H.-F.; Wu, Q.-X.; Zhu, Y. Sesquiterpenes and diterpenes with cytotoxic activities from the aerial parts of Carpesium humile. Fitoterapia 2018, 128, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Zhu, N.-L.; Tang, C.; Xu, C.; Ke, C.-Q.; Lin, G.; Jenis, J.; Yao, S.; Liu, H.; Ye, Y. Cytotoxic germacrane-type sesquiterpene lactones from the whole plant of Carpesium lipskyi. J. Nat. Prod. 2019, 82, 919–927. [Google Scholar] [CrossRef]
- Zhang, T.; Zhang, Q.-B.; Fu, L.; Li, L.-Y.; Ma, L.-Y.; Si, J.-G.; Zhang, H.-W.; Wei, J.-H.; Yu, S.-S.; Zou, Z.-M. New antiproliferative germacranolides from Carpesium divaricatum. RSC Adv. 2019, 9, 11493–11502. [Google Scholar] [CrossRef]
- Chen, X.; Song, J.; Yuan, D.; Rao, Q.; Jiang, K.; Feng, S.; Zhu, G.; Yan, C.; Li, Y.; Zhu, J. Incaspitolide A extracted from Carpesium cernuum induces apoptosis in vitro via the PI3K/AKT pathway in benign prostatic hyperplasia. Biosci. Rep. 2021, 41, BSR20210477. [Google Scholar] [CrossRef]
- Huang, Y.; Mao, J.; Zhang, L.; Guo, H.; Yan, C.; Chen, M. Antiprostate cancer activity of ineupatolide isolated from Carpesium cernuum L. Biomed Res. Int. 2021, 5515961. [Google Scholar] [CrossRef]
- Huang, Y.; Mao, J.; Zhang, L.; Guo, H.; Yan, C.; Chen, M. Incaspitolide A isolated from Carpesium cernuum L. inhibits the growth of prostate cancer cells and induces apoptosis via regulation of the PI3K/Akt/xIAP pathway. Oncol. Lett. 2021, 21, 477. [Google Scholar] [CrossRef]
- Kłeczek, N.; Malarz, J.; Gierlikowska, B.; Skalniak, Ł.; Galanty, A.; Kiss, A.K.; Stojakowska, A. Germacranolides from Carpesium divaricatum: Some New Data on Cytotoxic and Anti-Inflammatory Activity. Molecules 2021, 26, 4644. [Google Scholar] [CrossRef]
- Zhao, J.; Thi, L.T.; Park, Y.G.; Jeong, B.R. Light quality affects growth and physiology of Carpesium triste Maxim. cultured in vitro. Agriculture 2020, 10, 258. [Google Scholar] [CrossRef]
- Anderberg, A.A.; Eldenäs, P.; Bayer, R.J.; Englund, M. Evolutionary relationships in the Asteraceae tribe Inuleae (incl. Plucheeae) evidenced by DNA sequences of ndhF; with notes on the systematic positions of some aberrant genera. Org. Divers. Evol. 2005, 5, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Englund, M.; Pornpongrungrueng, P.; Gustafsson, M.H.G.; Anderberg, A.A. Phylogenetic relationships and generic delimitation in Inuleae subtribe Inulinae (Asteraceae) based on ITS and cpDNA sequence data. Cladistics 2009, 25, 319–352. [Google Scholar] [CrossRef] [PubMed]
- Stojakowska, A.; Malarz, J.; Kisiel, W. Terpenoids from a multiple shoot culture of Telekia speciosa. Acta Soc. Bot. Pol. 2011, 80, 253–256. [Google Scholar] [CrossRef] [Green Version]
- Wajs-Bonikowska, A.; Stojakowska, A.; Kalemba, D. Chemical Composition of Essential Oils from a Multiple Shoot Culture of Telekia speciosa and Different Plant Organs. Nat. Prod. Commun. 2012, 7, 625–628. [Google Scholar] [CrossRef] [Green Version]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Kim, D.K.; Lee, K.R.; Zee, O.P. Sesquiterpene lactones from Carpesium divaricatum. Phytochemistry 1997, 46, 1245–1247. [Google Scholar] [CrossRef]
- Goswami, A.C.; Baruah, R.N.; Sharma, R.P.; Baruah, J.N.; Kulanthaivel, P.; Herz, W. Germacranolides from Inula cappa. Phytochemistry 1984, 23, 367–372. [Google Scholar] [CrossRef]
- Zhang, T.; Si, J.-G.; Zhang, Q.-B.; Chen, J.-H.; Ding, G.; Zhang, H.-W.; Jia, H.-M.; Zou, Z.-M. Three new highly oxygenated germacranolides from Carpesium divaricatum and their cytotoxic activity. Molecules 2018, 23, 1078. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Chen, J.-H.; Si, J.-G.; Ding, G.; Zhang, Q.-B.; Zhang, H.-W.; Jia, H.-M.; Zou, Z.-M. Isolation, structure elucidation, and absolute configuration of germecrane isomers from Carpesium divaricatum. Sci. Rep. 2018, 8, 12418. [Google Scholar] [CrossRef]
- Stojakowska, A.; Michalska, K.; Malarz, J. Simultaneous quantification of eudesmanolides and thymol derivatives from tissues of Inula helenium and I. royleana by reversed-phase high-performance liquid chromatography. Phytochem. Anal. 2006, 17, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Zee, O.P.; Kim, D.K.; Lee, K.R. Thymol derivatives from Carpesium divaricatum. Arch. Pharm. Res. 1998, 21, 618–620. [Google Scholar] [CrossRef] [PubMed]
- Stojakowska, A.; Malarz, J.; Kisiel, W. Quantitative analysis of sesquiterpene lactones and thymol derivatives in extracts from Telekia speciosa. Phytochem. Lett. 2015, 11, 378–383. [Google Scholar] [CrossRef]
- Stojakowska, A.; Malarz, J. In vitro propagation of Inula royleana DC. Acta Soc. Bot. Pol. 2004, 73, 5–8. [Google Scholar] [CrossRef] [Green Version]
- Trejgell, A.; Kamińska, M.; Lisowska, K.; Tretyn, A. Micropropagation of Inula germanica L. from the seedlings explants. Not. Bot. Horti Agrobot. Cluj Napoca 2018, 46, 52–57. [Google Scholar] [CrossRef] [Green Version]
- Woerdenbag, H.J.; Lüers, J.F.J.; van Uden, W.; Pras, N.; Malingré, T.M.; Alfermann, A.W. Production of the new antimalarial drug artemisinin in shoot cultures of Artemisia annua L. Plant Cell Tissue Organ Cult. 1993, 32, 247–257. [Google Scholar] [CrossRef]
- Stojakowska, A.; Kisiel, W. Production of parthenolide in organ cultures of feverfew. Plant Cell Tissue Organ Cult. 1997, 47, 159–162. [Google Scholar] [CrossRef]
- Brown, A.M.G.; Lowe, K.C.; Davey, M.R.; Power, J.B. Feverfew (Tanacetum parthenium L.): Tissue culture and parthenolide synthesis. Plant Sci. 1996, 116, 223–232. [Google Scholar] [CrossRef]
- Malarz, J.; Stojakowska, A.; Dohnal, B.; Kisiel, W. Helenalin acetate in in vitro propagated plants of Arnica montana. Planta Med. 1993, 59, 51–53. [Google Scholar] [CrossRef]
- Kayani, W.K.; Kiani, B.H.; Dilshad, E.; Mirza, B. Biotechnological approaches for artemisinin production in Artemisia. World J. Microbiol. Biotechnol. 2018, 34, 54. [Google Scholar] [CrossRef]
- Poulson, M.E.; Thai, T. Effect of high light intensity on photoinhibition, oxyradicals and artemisinin content in Artemisia annua L. Photosynthetica 2015, 53, 403–409. [Google Scholar] [CrossRef]
- Štampar, M.; Breznik, B.; Filipič, M.; Žegura, B. Characterization of in vitro 3D cell model developed from human hepatocellular carcinoma (HepG2) Cell Line. Cells 2020, 9, 2557. [Google Scholar] [CrossRef] [PubMed]
- Nikolaou, N.; Green, C.J.; Gunn, P.J.; Hodson, L.; Tomlinson, J.W. Optimizing human hepatocyte models for metabolic phenotype and function: Effects of treatment with dimethyl sulfoxide (DMSO). Physiol. Rep. 2016, 4, e12944. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.K.; Baek, N.I.; Choi, S.U.; Lee, C.O.; Lee, K.R.; Zee, O.P. Four new cytotoxic germacranolides from Carpesium divaricatum. J. Nat. Prod. 1997, 60, 1199–1202. [Google Scholar] [CrossRef]
- Zhang, T.; Si, J.-G.; Zhang, Q.-B.; Ding, G.; Zou, Z.-M. New highly oxygenated germacranolides from Carpesium divaricatum and their cytotoxic activity. Sci. Rep. 2016, 6, 27237. [Google Scholar] [CrossRef]
- Dürr, L.; Hell, T.; Dobrzyński, M.; Mattei, A.; John, A.; Augsburger, N.; Bradanini, G.; Reinhardt, J.K.; Rossberg, F.; Drobnjakovic, M.; et al. High-content screening pipeline for natural products targeting oncogenic signaling in melanoma. J. Nat. Prod. 2022, 85, 1006–1017. [Google Scholar] [CrossRef]
- Fadel, H.; Sifaoui, I.; Lόpez-Arencibia, A.; Reyes-Batlle, M.; Jiménez, I.A.; Lorenzo-Morales, J.; Ghedadba, N.; Benayache, S.; Piñero, J.E.; Bazzocchi, I.L. Antioxidant and leishmanicidal evaluation of Pulicaria inuloides root extracts: A bioguided fractionation. Pathogens 2019, 8, 201. [Google Scholar] [CrossRef] [Green Version]
- Talavera-Alemán, A.; Rodríguez-García, G.; López, Y.; García-Gutiérrez, H.A.; Torres-Valencia, J.; Del Río, R.E.; Cerda-García-Rojas, C.M.; Joseph-Nathan, P.; Gómez-Hurtado, M.A. Systematic evaluation of thymol derivatives possessing stereogenic or prostereogenic centers. Phytochem. Rev. 2015, 15, 251–277. [Google Scholar] [CrossRef]
- Sołtys, A.; Galanty, A.; Zagrodzki, P.; Grabowska, K.; Malarz, J.; Podolak, I. Sorbus intermedia (EHRH.) PERS. fruits as a novel source of biologically active triterpenoids–Comparative studies of ursolic acid derivatives with cytotoxic potential. Biomed. Pharmacother. 2022, 154, 113592. [Google Scholar] [CrossRef]
- Grabowska, K.; Amanowicz, K.; Paśko, P.; Podolak, I.; Galanty, A. Optimization of the extraction procedure for the phenolic-rich Glechoma hederacea L. herb and evaluation of its cytotoxic and antioxidant potential. Plants 2022, 11, 2217. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Culture Medium | Light Conditions | Regeneration of Shoots (%) | Number of Shoots per Explant | Final Dry Weight per Explant (g) 3 |
|---|---|---|---|---|
| MS + 0.5 μM TDZ; solidified medium 1 | continuous illumination; cool white light; 40 μmol m−2 s−1 | 100 | 3.36 ± 1.50 1,a | 0.131 ± 0.023 b |
| MS + 2.0 μM BA + 0.1 μM NAA; solidified medium 1 | continuous illumination; cool white light; 40 μmol m−2 s−1 | 96.8 | 3.73 ± 1.43 1,a | 0.067 ± 0.014 a |
| MS + 4.44 μM BA + 0.54 μM NAA; solidified medium 1 | continuous illumination; cool white light; 40 μmol m−2 s−1 | 94.5 | 3.49 ± 1.22 1,a | 0.081 ± 0.017 a |
| MS + 0.5 μM TDZ; liquid medium 2 | photoperiod 16/8 (light/dark); cool white light; 20 μmol m−2 s−1 | 100 | 3.78 ± 1.37 2,a | 0.216 ± 0.042 c |
| MS + 1.0 μM TDZ; liquid medium 2 | photoperiod 16/8 (light/dark); cool white light; 20 μmol m−2 s−1 | 100 | 4.07 ± 1.61 2,a | 0.269 ± 0.028 c |
| Plant Extract | Contents of the Terpenoids (% DW) 1 | ||||||
|---|---|---|---|---|---|---|---|
| A | B | C | D | E | F | G | |
| 05TDZL 2 | 0.016 ± 0.002 a | 0.026 ± 0.002 a | 0.036 ± 0.004 a | 0.021 ± 0.003 a | 0.027 ± 0.004 a | 0.018 ± 0.003 a | 0.064 ± 0.008 a |
| 05TDZS | 0.034 ± 0.003 a,b | 0.049 ± 0.006 b | 0.018 ± 0.001 b | 0.083 ± 0.004 b | 0.040 ± 0.005 a | 0.094 ± 0.016 b | 0.040 ± 0.001 b |
| 20B01NS | 0.051 ± 0.004 b,c | 0.114 ± 0.002 c | 0.138 ± 0.001 c | 0.081 ± 0.002 b | 0.104 ± 0.002 b | 0.034 ± 0.001 a | 0.043 ± 0.004 b |
| 44B05NS | 0.077 ± 0.010 d | 0.039 ± 0.001 b | 0.023 ± 0.001 b | 0.139 ± 0.019 c | 0.079 ± 0.002 c | 0.214 ± 0.021 c | 0.028 ± 0.004 b |
| CdN2020 | 0.047 ± 0.001 b | 0.079 ± 0.006 d | 0.031 ± 0.001 a | 0.070 ± 0.002 b | 0.049 ± 0.001 a | 0.179 ± 0.007 c | nd |
| CdN2022 | 0.060 ± 0.007 c,d | 0.089 ± 0.006 d | 0.171 ± 0.004 d | 0.018 ± 0.001 a | 0.201 ± 0.013 d | nd | nd |
| CdK2022 | nd | nd | nd | nd | nd | nd | 0.118 ± 0.008 c |
| Plant Extract and Time of Exposure (h) | IC50 (μg/mL) 1 | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Prostate Panel | Skin Panel | Gastrointestinal Panel | |||||||
| PNT2 | Du145 | PC3 | HaCaT | A375 | HTB140 | HT29 | Caco2 | HepG2 | |
| 05TDZL 24 | 27.48 ± 1.12 a | 79.18 ± 2.96 b | 44.94 ± 1.08 c | >100 | 34.96 ± 1.02 a | 37.94 ± 1.24 b | 46.53 ± 1.51 a | 28.51 ± 0.98 b | >100 |
| 05TDZL 48 | 21.42 ± 1.24 a | 54.59 ± 2.06 b | 32.63 ± 1.41 c | >100 | 30.40 ± 0.80 a | 33.89 ± 1.06 b | 42.11 ± 1.03 a | 25.64 ± 0.74 b | >100 |
| 20B01NS 24 | 43.17 ± 1.77 a | 76.24 ± 2.79 b | 26.50 ± 0.77 c | >100 | 42.29 ± 1.33 a | 33.97 ± 1.41 b | 64.78 ± 1.81 a | 26.13 ± 1.04 b | >100 |
| 20B01NS 48 | 30.23 ± 1.79 a | 42.48 ± 1.33 b | 14.07 ± 0.76 c | 94.15 ± 3.38 a | 32.07 ± 1.11 b | 19.87 ± 0.72 c | 52.76 ± 1.92 a | 14.62 ± 1.41 b | >100 |
| CdN2020 24 | 47.04 ± 1.84 a | 45.59 ± 1.77 a | 29.00 ± 1.41 b | >100 | 68.04 ± 2.59 a | 39.06 ± 1.12 b | 47.56 ± 1.00 a | 24.64 ± 1.57 b | >100 |
| CdN2020 48 | 32.89 ± 1.30 a | 24.19 ± 0.95 b | 21.18 ± 1.33 b | >100 | 42.91 ± 1.50 a | 26.12 ± 1.20 b | 21.18 ± 1.33 a | 16.21 ± 1.42 b | 98.27 ± 2.95 |
| CdN2022 24 | 34.88 ± 1.10 a | 74.36 ± 1.91 b | 64.48 ± 1.59 c | 95.05 ± 3.85 a | 49.91 ± 1.58 b | 27.43 ± 1.09 c | 58.45 ± 1.87 a | 37.42 ± 2.29 b | 98.41 ± 3.49 c |
| CdN2022 48 | 29.69 ± 1.28 a | 53.56 ± 1.54 b | 54.04 ± 1.97 b | 75.64 ± 3.06 a | 39.79 ± 2.39 b | 21.92 ± 1.04 c | 49.06 ± 1.79 a | 26.36 ± 2.04 b | 74.93 ± 2.12 c |
| CdK2022 24 | 50.62 ± 1.86 a | 80.61 ± 3.34 b | 43.61 ± 1.98 c | >100 | 74.14 ± 2.27 a | 46.13 ± 1.29 b | 75.00 ± 2.52 a | 53.93 ± 1.97 b | >100 |
| CdK2022 48 | 33.81 ± 1.47 a | 61.28 ± 1.58 b | 34.56 ± 2.28 a | >100 | 68.02 ± 2.32 a | 28.17 ± 1.80 b | 64.84 ± 1.64 a | 32.32 ± 1.47 b | >100 |
| Doxorubicin 24 | 1.38 | 3.18 | >50 | 4.68 | 0.59 | 5.71 | 1.53 | 3.44 | 1.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malarz, J.; Galanty, A.; Stojakowska, A. Chemical Composition and Cytotoxic Activity of Extracts from Carpesium divaricatum: In Vitro- versus Field-Grown Plants. Plants 2022, 11, 2815. https://doi.org/10.3390/plants11212815
Malarz J, Galanty A, Stojakowska A. Chemical Composition and Cytotoxic Activity of Extracts from Carpesium divaricatum: In Vitro- versus Field-Grown Plants. Plants. 2022; 11(21):2815. https://doi.org/10.3390/plants11212815
Chicago/Turabian StyleMalarz, Janusz, Agnieszka Galanty, and Anna Stojakowska. 2022. "Chemical Composition and Cytotoxic Activity of Extracts from Carpesium divaricatum: In Vitro- versus Field-Grown Plants" Plants 11, no. 21: 2815. https://doi.org/10.3390/plants11212815
APA StyleMalarz, J., Galanty, A., & Stojakowska, A. (2022). Chemical Composition and Cytotoxic Activity of Extracts from Carpesium divaricatum: In Vitro- versus Field-Grown Plants. Plants, 11(21), 2815. https://doi.org/10.3390/plants11212815