
Color Variation and Secondary Metabolites’ Footprint in a Taxonomic Complex of Phyteuma sp. (Campanulaceae)



Abstract
:1. Introduction
2. Results
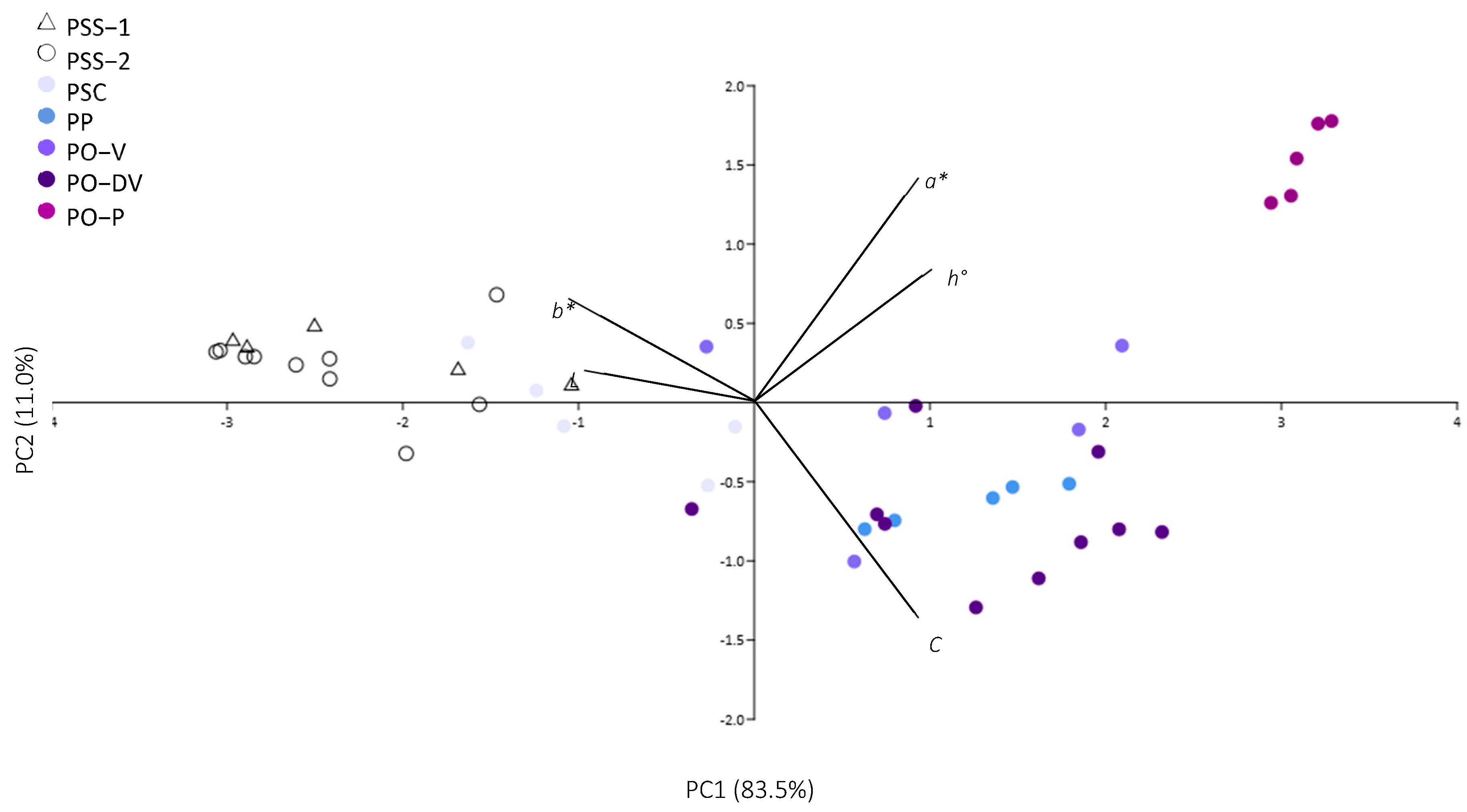
2.1. Analysis of Color
2.1.1. Color Variation in Natural Populations
- P. spicatum ssp. spicatum (abbreviated further as PSS) showed white-greenish flowers. Some populations were growing alone (PSS-1), while others were growing in sympatry with P. spicatum ssp. caeruleum (PSS-2);
- P. spicatum ssp. caeruleum (PSC) showed white flowers with a very light violet tone;
- P. ovatum (PO) shows a typical violet color, although a wider variety of colors were observed on the field. The individuals with violet flowers were separated into two subgroups: one with typical violet flowers (PO-V) and the other with dark violet flowers (PO-DV). Some individuals showed purple flowers with a high presence of reddish tones (PO-P);
- P. persicifolium (PP) showed typical blue color.
2.1.2. Numeric Analysis of Color
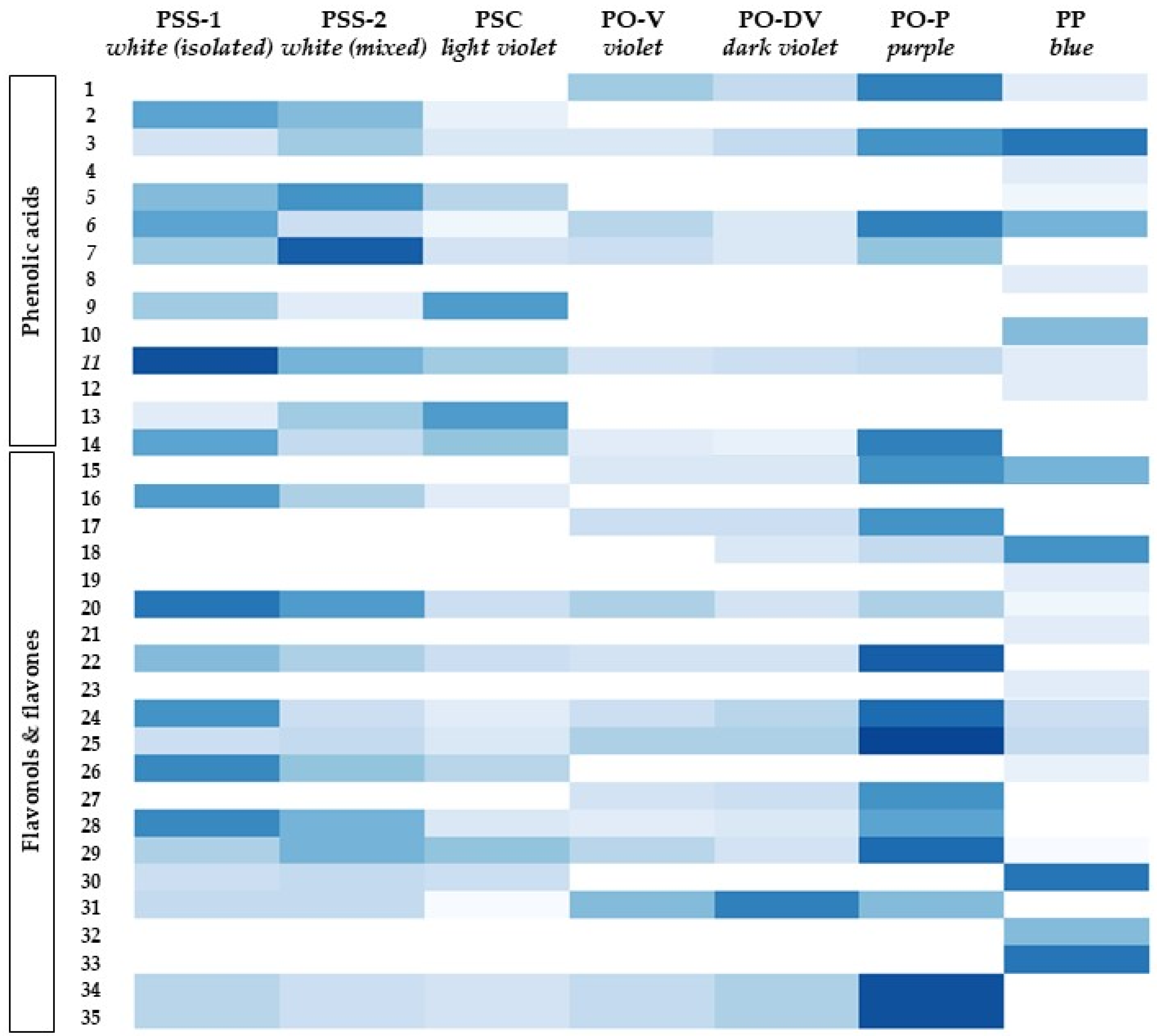
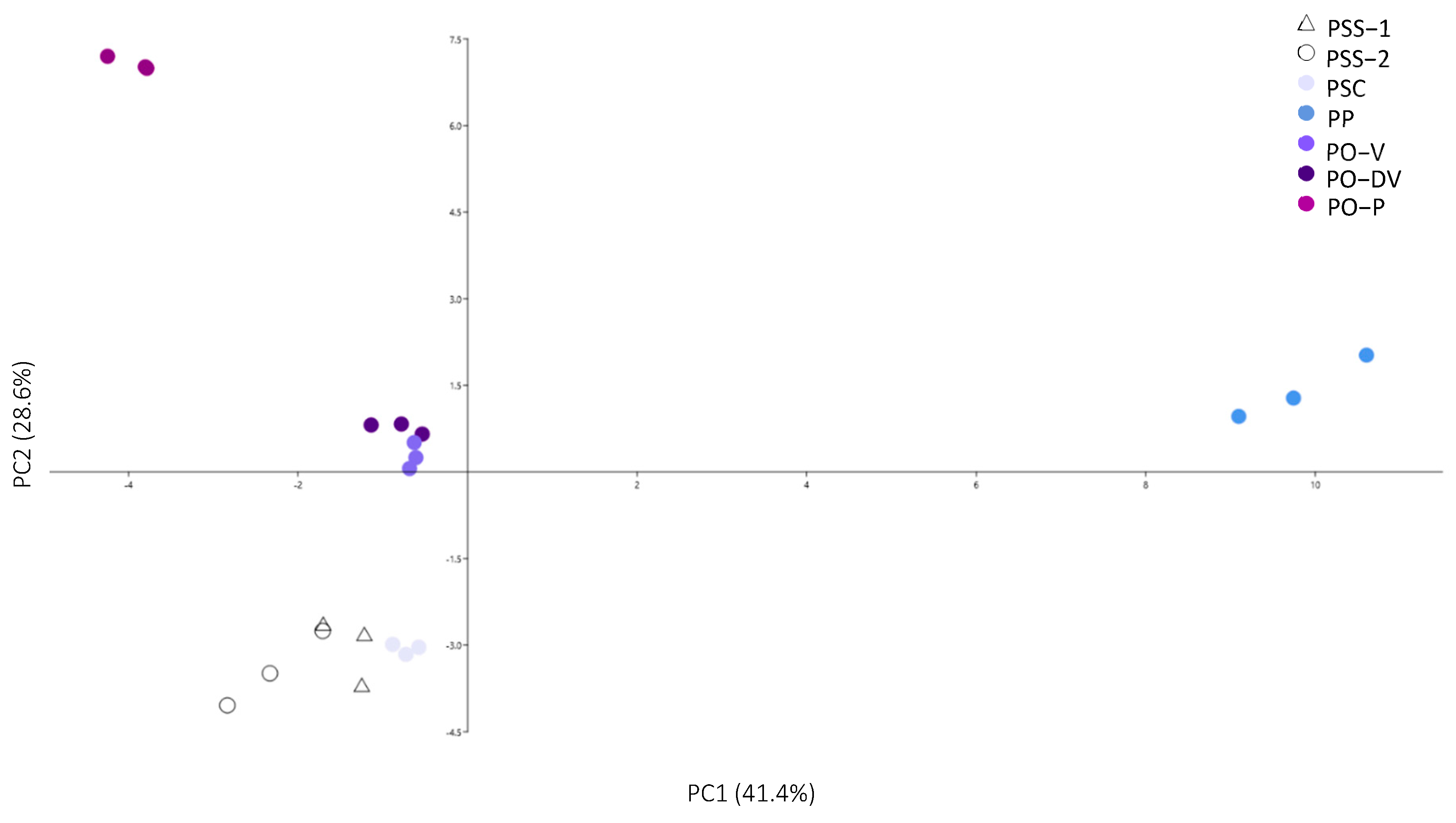
2.2. Phenolic Compounds Identification and Content
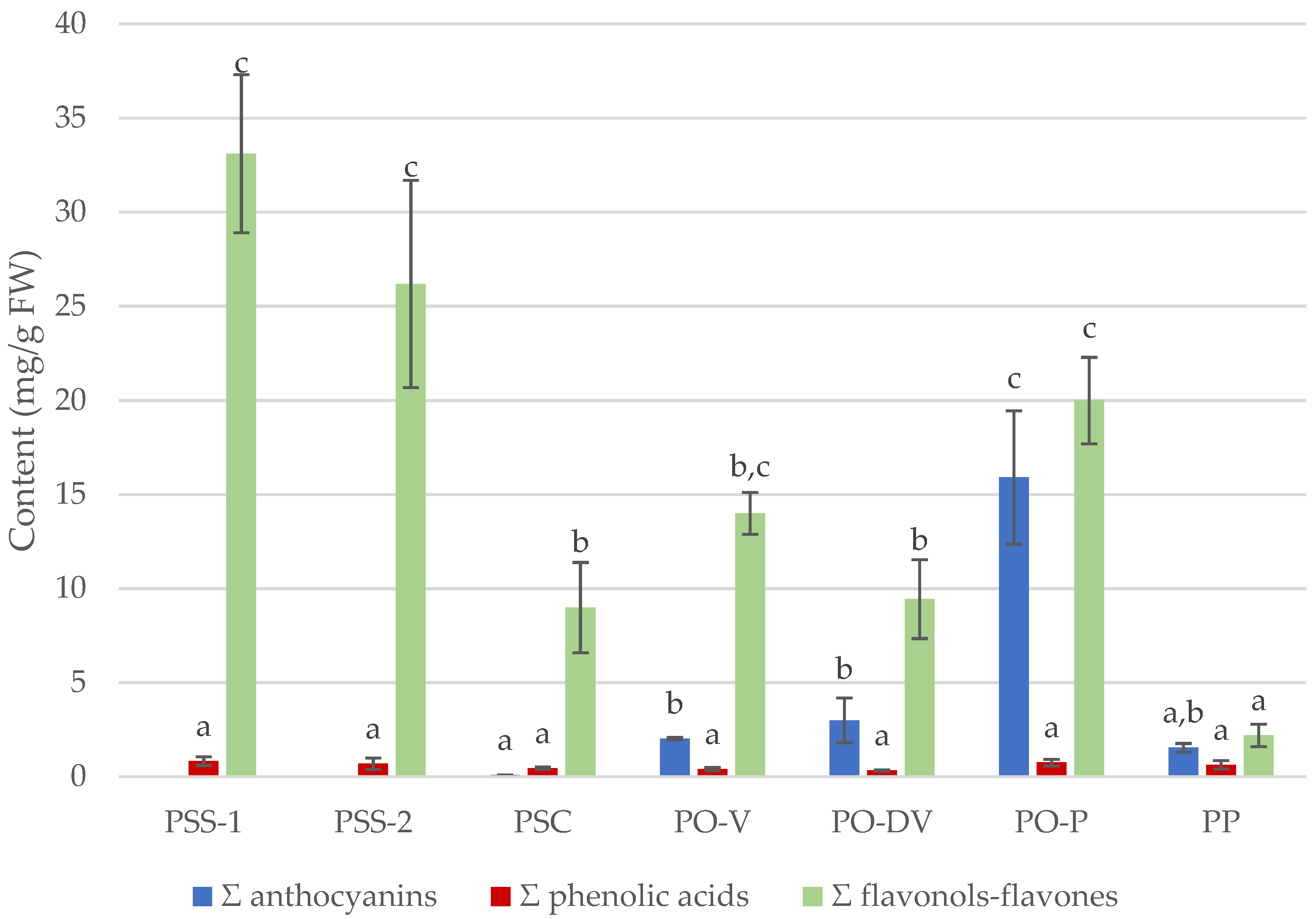
2.2.1. Anthocyanins
2.2.2. Other Phenolic Compounds
2.2.3. Total Contents of Phenolic Compounds
3. Discussion
3.1. Numeric Analysis of Color
3.2. Phenolic Compounds Identification and Content
3.2.1. Anthocyanins
3.2.2. Other Phenolic Compounds
4. Materials and Methods
4.1. Plant Material
4.2. Color Evaluation
4.3. Extraction of Phenolic Compounds
4.4. Analysis with HPLC-MS
4.5. Chemicals
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Damboldt, J.; Phyteuma, L. Flora Europaea; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar]
- Schulz, R. Monographie der Gattung Phyteuma; Schneck, J., Ed.; J. Shneck: Geisenheim am Rhein, Germany, 1904. [Google Scholar]
- Martinčič, A.; Wraber, T.; Jogan, N.; Podobnik, A.; Turk, B.; Vreš, B.; Ravnik, V.; Frajman, B.; Strgulc-Krajšek, S.; Trčak, B.; et al. Mala Flora Slovenije, 4th ed.; Tehniška založba Slovenije: Ljubljana, Slovenia, 2007. [Google Scholar]
- Schneeweiss, G.M.; Pachschwöll, C.; Tribsch, A.; Schönswetter, P.; Barfuss, M.H.J.; Esfeld, K.; Weiss-Schneeweiss, H.; Thiv, M. Molecular Phylogenetic Analyses Identify Alpine Differentiation and Dysploid Chromosome Number Changes as Major Forces for the Evolution of the European Endemic Phyteuma (Campanulaceae). Mol. Phylogenet. Evol. 2013, 69, 634–652. [Google Scholar] [CrossRef] [PubMed]
- Huber, R. Biosystematische Untersuchungen an Phyteuma Spicatum Und Phyteuma Ovatum: Inaugural-Dissertation zur Erlangung der Philosophischen Doktorwürde Vorgelegt der Philosophischen Fakultät II der Universität Zürich. Ph.D. Thesis, University of Zurich, Zurich, Switzerland, 1988. [Google Scholar]
- Zhou, Y.; Yin, M.; Abbas, F.; Sun, Y.; Gao, T.; Yan, F.; Li, X.; Yu, Y.; Yue, Y.; Yu, R.; et al. Classification and Association Analysis of Gerbera (Gerbera hybrida) Flower Color Traits. Front. Plant Sci. 2021, 12, 779288. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Yang, S.; Liu, X.; Cui, J.; Chang, Z.; Ge, H. The Diversity of Petal Colors and Its Relation to Anthocyanins in Rose Germplasms. Acta Hortic. Sin. 2017, 44, 1125–1134. [Google Scholar]
- Cui, H.; Zhang, Y.; Shi, X.; Gong, F.; Xiong, X.; Kang, X.; Xing, G.; Li, S. The Numerical Classification and Grading Standards of Daylily (Hemerocallis) Flower Color. PLoS ONE 2019, 14, e0216460. [Google Scholar] [CrossRef]
- Abe, H.; Miura, H.; Motonaga, Y. Quantitative Classification of Camellia japonica and Camellia rusticana (Theaceae) Based on Leaf and Flower Morphology. Plant Divers 2021, 43, 216–224. [Google Scholar] [CrossRef]
- Gawenda-Kempczyńska, D.; Olech, M.; Balcerek, M.; Nowak, R.; Załuski, T.; Załuski, D. Phenolic Acids as Chemotaxonomic Markers Able to Differentiate the Euphrasia Species. Phytochemistry 2022, 203, 113342. [Google Scholar] [CrossRef]
- Tatsuzawa, F. Flower Colors and Flavonoids in the Cultivars of Lobelia erinus L. (Campanulaceae). Dye Pigment 2020, 180, 108500. [Google Scholar] [CrossRef]
- Pandey, A.; Misra, P.; Bhambhani, S.; Bhatia, C.; Trivedi, P.K. Expression of Arabidopsis MYB Transcription Factor, AtMYB111, in Tobacco Requires Light to Modulate Flavonol Content. Sci. Rep. 2014, 4, 5018. [Google Scholar] [CrossRef] [Green Version]
- Eiro, M.J.; Heinonen, M. Anthocyanin Color Behavior and Stability during Storage: Effect of Intermolecular Copigmentation. J. Agric. Food Chem. 2002, 50, 7461–7466. [Google Scholar] [CrossRef]
- Castañeda-Ovando, A.; de Lourdes Pacheco-Hernández, M.; Páez-Hernández, M.E.; Rodríguez, J.A.; Galán-Vidal, C.A. Chemical Studies of Anthocyanins: A Review. Food Chem. 2009, 113, 859–871. [Google Scholar] [CrossRef]
- Klisurova, D.; Petrova, I.; Ognyanov, M.; Georgiev, Y.; Kratchanova, M.; Denev, P. Co-Pigmentation of Black Chokeberry (Aronia melanocarpa) Anthocyanins with Phenolic Co-Pigments and Herbal Extracts. Food Chem. 2019, 279, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, F.; Ji, C.; Liu, D.; Liu, X.; Wang, R.; Li, W. Chemical Constituents of the Pericarp of Toona sinensis and Their Chemotaxonomic Significance. Biochem. Syst. Ecol. 2022, 104, 104458. [Google Scholar] [CrossRef]
- Zhang, Y.; Ma, J.-T.; Feng, T.; Liu, J.-K.; He, J. Chemical Constituents from Twigs and Leaves of Actinidia chinensis Planch and Their Chemotaxonomic Significance. Biochem. Syst. Ecol. 2022, 104, 104461. [Google Scholar] [CrossRef]
- Gu, W.; Xie, R.; Chen, J.; Yuan, C.; Huang, L.; Yi, P.; Hao, X. Two New Secondary Metabolites from Oreocharis auricula and Their Chemotaxonomic Significance. Biochem. Syst. Ecol. 2022, 104, 104477. [Google Scholar] [CrossRef]
- Janković, I.B.; Drobac, M.M.; Lakušić, D. V Compounds of the Methanolic Leaf Extract as Chemotaxonomic Markers for the Campanula pyramidalis Complex (Campanulaceae). Acta Bot. Croat. 2014, 73, 481–490. [Google Scholar] [CrossRef] [Green Version]
- Abbet, C.; Neuburger, M.; Wagner, T.; Quitschau, M.; Hamburger, M.; Potterat, O. Phyteumosides A and B: New Saponins with Unique Triterpenoid Aglycons from Phyteuma orbiculare L. Org. Lett. 2011, 13, 1354–1357. [Google Scholar] [CrossRef] [PubMed]
- Konishi, T.; Tada, A.; Shoji, J.; Kasai, R.; Tanaka, O. The Structures of Platycodin A and C, Monoacetylated Saponins of the Roots of Platycodon Grandiflorum A. DC. Chem. Pharm. Bull. Tokyo 1978, 26, 668–670. [Google Scholar] [CrossRef] [Green Version]
- Abbet, C.; Slacanin, I.; Hamburger, M.; Potterat, O. Comprehensive Analysis of Phyteuma orbiculare L., a Wild Alpine Food Plant. Food Chem. 2013, 136, 595–603. [Google Scholar] [CrossRef]
- Brandt, K.; Ishimaru, K. Campanula (Bellflower) Species: In Vitro Culture, Micropropagation, and the Production of Anthocyanins, Polyacetylenes, and Other Secondary Metabolites. In Medicinal and Aromatic Plants X; Springer: Berlin/Heidelberg, Germany, 1998; pp. 45–66. [Google Scholar]
- Justesen, H.; Andersen, A.S.; Brandt, K. Accumulation of Anthocyanins and Flavones during Bud and Flower Development in Campanula isophylla Moretti. Ann. Bot. 1997, 79, 355–360. [Google Scholar] [CrossRef] [Green Version]
- Saito, N.; Tatsuzawa, F.; Yazaki, Y.; Shigihara, A.; Honda, T. 7-Polyacylated Delphinidin 3, 7-Diglucosides from the Blue Flowers of Leschenaultia cv. Violet Lena. Phytochemistry 2007, 68, 673–679. [Google Scholar] [CrossRef]
- Gao, L.; Yang, H.; Liu, H.; Yang, J.; Hu, Y. Extensive Transcriptome Changes Underlying the Flower Color Intensity Variation in Paeonia ostii. Front. Plant Sci. 2016, 6, 1205. [Google Scholar] [CrossRef] [PubMed]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and Anthocyanins: Colored Pigments as Food, Pharmaceutical Ingredients, and the Potential Health Benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [Green Version]
- Polatschek, A. Cytotaxonomische Beiträge Zur Flora Der Ostalpenländer, II. Osterr. Bot. Z. 1966, 113, 101–147. [Google Scholar] [CrossRef]
- Tison, J.M.; de Foucault, B. Flora Gallica; Biotope: Mèze, France, 2014. [Google Scholar]
- Brunerye, L. Note Sur Les Phyteuma Du Groupe Spicatum s. 1. de La Flore de France. Bull. Société Bot. Cent.-Ouest NS 1989, 20, 13–21. [Google Scholar]
- Eggenberg, S.; Bornand, C.; Juillerat, P.; Jutzi, M.; Möhl, A.; Nyffeler, R.; Santiago, H. Flora Helvetica. In Guide d’Excursions; Haupt: Bern, Switzerland, 2018. [Google Scholar]
- de Bolòs, O.; Bonada, J.V. Flora Dels Països Catalans, 1st ed.; Institut d’Estudis Catalans: Barcino, Spain, 1984; Volume 2, ISBN 8472266206. [Google Scholar]
- Liu, Y.; Tikunov, Y.; Schouten, R.E.; Marcelis, L.F.M.; Visser, R.G.F.; Bovy, A. Anthocyanin Biosynthesis and Degradation Mechanisms in Solanaceous Vegetables: A Review. Front. Chem. 2018, 6, 52. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Wang, N.; Liu, H.; Li, Z.; Lu, L.; Wang, C. The Bioactive Compounds and Biological Functions of Asparagus officinalis L.—A Review. J. Funct. Foods 2020, 65, 103727. [Google Scholar] [CrossRef]
- Dixon, R.A.; Achnine, L.; Kota, P.; Liu, C.-J.; Reddy, M.S.S.; Wang, L. The Phenylpropanoid Pathway and Plant Defence—A Genomics Perspective. Mol. Plant Pathol. 2002, 3, 371–390. [Google Scholar] [CrossRef]
- Meléndez-Martínez, A.J.; Benítez, A.; Corell, M.; Hernanz, D.; Mapelli-Brahm, P.; Stinco, C.; Coyago-Cruz, E. Screening for Innovative Sources of Carotenoids and Phenolic Antioxidants among Flowers. Foods 2021, 10, 2625. [Google Scholar] [CrossRef]
- Saito, N.; Tatsuzawa, F.; Honda, T. Light Absorption Spectral Patterns of Intact Garden Flowers in Relation to the Flower Colors and Anthocyanin Pigments. Heterocycles 2015, 90, 41–84. [Google Scholar]
- Mikulic-Petkovsek, M.; Slatnar, A.; Schmitzer, V.; Stampar, F.; Veberic, R.; Koron, D. Chemical Profile of Black Currant Fruit Modified by Different Degree of Infection with Black Currant Leaf Spot. Sci. Hortic. 2013, 150, 399–409. [Google Scholar] [CrossRef]
- Ruiz, A.; Hermosín-Gutiérrez, I.; Vergara, C.; von Baer, D.; Zapata, M.; Hitschfeld, A.; Obando, L.; Mardones, C. Anthocyanin Profiles in South Patagonian Wild Berries by HPLC-DAD-ESI-MS/MS. Food Res. Int. 2013, 51, 706–713. [Google Scholar] [CrossRef]
- Said, R.B.; Hamed, A.I.; Mahelel, U.A.; Al-Ayed, A.S.; Kowalczyk, M.; Moldoch, J.; Oleszek, W.; Stochmal, A. Tentative Characterization of Polyphenolic Compounds in the Male Flowers of Phoenix Dactylifera by Liquid Chromatography Coupled with Mass Spectrometry and DFT. Int. J. Mol. Sci. 2017, 18, 512. [Google Scholar] [CrossRef] [PubMed]
- di Lecce, G.; Martínez-Huélamo, M.; Tulipani, S.; Vallverdú-Queralt, A.; Lamuela-Raventós, R.M. Setup of a UHPLC–QqQ-MS Method for the Analysis of Phenolic Compounds in Cherry Tomatoes, Tomato Sauce, and Tomato Juice. J. Agric. Food Chem. 2013, 61, 8373–8380. [Google Scholar] [CrossRef]
- Díaz-de-Cerio, E.; Gómez-Caravaca, A.M.; Verardo, V.; Fernández-Gutiérrez, A.; Segura-Carretero, A. Determination of Guava (Psidium guajava L.) Leaf Phenolic Compounds Using HPLC-DAD-QTOF-MS. J. Funct. Foods 2016, 22, 376–388. [Google Scholar] [CrossRef]
- Gómez-Romero, M.; Zurek, G.; Schneider, B.; Baessmann, C.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Automated Identification of Phenolics in Plant-Derived Foods by Using Library Search Approach. Food Chem. 2011, 124, 379–386. [Google Scholar] [CrossRef]
- Mustafa, A.M.; Angeloni, S.; Abouelenein, D.; Acquaticci, L.; Xiao, J.; Sagratini, G.; Maggi, F.; Vittori, S.; Caprioli, G. A New HPLC-MS/MS Method for the Simultaneous Determination of 36 Polyphenols in Blueberry, Strawberry and Their Commercial Products and Determination of Antioxidant Activity. Food Chem. 2022, 367, 130743. [Google Scholar] [CrossRef]
- Sobral, F.; Calhelha, R.C.; Barros, L.; Dueñas, M.; Tomás, A.; Santos-Buelga, C.; Vilas-Boas, M.; Ferreira, I.C.F.R. Flavonoid Composition and Antitumor Activity of Bee Bread Collected in Northeast Portugal. Molecules 2017, 22, 248. [Google Scholar] [CrossRef] [Green Version]
- Verardo, V.; Arráez-Román, D.; Segura-Carretero, A.; Marconi, E.; Fernández-Gutiérrez, A.; Caboni, M.F. Identification of Buckwheat Phenolic Compounds by Reverse Phase High Performance Liquid Chromatography–Electrospray Ionization-Time of Flight-Mass Spectrometry (RP-HPLC–ESI-TOF-MS). J. Cereal Sci. 2010, 52, 170–176. [Google Scholar] [CrossRef]
- Xie, L.; Lin, Q.; Guo, K.; Tong, C.; Shi, S.; Shi, F. HPLC-DAD-QTOF-MS/MS Based Comprehensive Metabolomic Profiling of Phenolic Compounds in Kalimeris Indica Anti-Inflammatory Fractions. Ind. Crops Prod. 2019, 140, 111636. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Flower Color | L* | a | b | C* | h° |
|---|---|---|---|---|---|---|
| PSS-1 | white | 58.3 ± 5.3 e | −0.7 ± 1.3 a | −34.2 ± 2.7 c | 24.2 ± 2.7 d | 268.5 ± 3.0 c |
| PSS-2 | white | 51.4 ± 2.9 d | −0.5 ± 1.7 a | −33.4 ± 4.9 c | 23.4 ± 4.9 d | 269.5 ± 3.9 c |
| PSC | light violet | 45.8 ± 2.2 c | 1.6 ± 0.8 b | −41.8 ± 4.9 b | 31.7 ± 4.9 c | 275.6 ± 4.1 c |
| PO-V | violet | 37.2 ± 3.3 b | 4.2 ± 1.7 c | −51.0 ± 4.5 a | 39.6 ± 4.5 b | 299.4 ± 20.2 b |
| PO-DV | dark violet | 29.9 ± 6.4 a | 2.6 ± 1.3 b | −53.5 ± 4.5 a | 42.7 ± 4.2 a,b | 298.0 ± 15.8 >b |
| PO-P | purple | 24.9 ± 2.9 a | 10.9 ± 0.9 d | −55.8 ± 0.6 a | 38.3 ± 0.7 b | 339.1 ± 4.1 a |
| PP | blue | 42.2 ± 1.2 b,c | 3.0 ± 0.4 b,c | −55.3 ± 1.8 a | 44.3 ± 1.3 a | 304.8 ± 13.4 b |
| Peak | RT (min) | ʎmax | [M + H]+ (m/z) | MS2 (m/z) | MS3 (m/z) | Tentative Identification |
|---|---|---|---|---|---|---|
| 1 | 8.9 | 527 | 611 | 303(100), 465(16) | Delphinidin-3-rutinoside | |
| 2 | 10.2 | 517 | 595 | 287(100), 449(19) | Cyanidin-3-rutinoside | |
| 3 | 12.1 | 528 | 463 | 301(100) | Peonidin-3-glucoside | |
| 4 | 14.7 | 535 | 1175.6 | 867(100), 611(44), 465(5) | [465] 303(100); [611] 303(100), 465(17) | Delphinidin rutinoside der. |
| 5 | 16.7 | 523, 533 | 727 | 317(100), 479(71) | Petunidin-3-rutinoside der. | |
| 6 | 18.6 | 530, 523 | 787.5 | 479(100), 299(29) | [479] 299(100); [299] 271(100), 255(24), 243(13) | Pelargonidin-3-rutinoside der. |
| 7 | 22.6 | 542 | 551 | 303(100) | Delphinidin hexoside der. |
| Population | Color | D3R | DRd | DHd | C3R | Po3G | PtRd | PlRd |
|---|---|---|---|---|---|---|---|---|
| PSS-1 | white | - | - | - | - | - | - | - |
| PSS-2 | white | - | - | - | - | traces | - | - |
| PSC | light violet | 5.8 ± 1.6 a | - | - | 1.9 ± 0.6 a | 2.2 ± 0.9 a | - | - |
| PO-V | violet | 72.8 ± 41.7 b | - | - | 128.3 ± 52.5 a | - | 1.4 ± 0.7 a | - |
| PO-DV | dark-violet | 180.7 ± 80.1 a | - | - | 106.6 ± 58.6 a | - | 12.2 ± 8.7 b | - |
| PO-P | purple | 58.5 ± 21.3 b | - | - | 1511.3 ± 42.3 b | 1.2 ± 0.1 a | - | 19.3 ± 1.2 b |
| PP | blue | 1.4 ± 0.1 c | 13.7 ± 3.0 a | traces | 136.4 ± 33.8 a | - | - | 2.2 ± 1.9 a |
| Peak | RT (min) | ʎmax | [M − H]− (m/z) | MS2 (m/z) | MS3 (m/z) | MS4 (m/z) | Tentative Identification |
|---|---|---|---|---|---|---|---|
| Phenolic acids | |||||||
| 1 | 8.03 | 279, 304 | 325 | 163(100) | p-Coumaric acid hexoside der. | ||
| 2 | 8.5 | 261 | 299 | 137(100), 179(63) | p-Hydroxybenzoic acid hexose | ||
| 3 | 9.04 | 312,279 | 325 | 163(100) | p-Coumaric acid der. | ||
| 4 | 9.4 | 306 | 353 | 191(100), 179(46) | Neochlorogenic acid | ||
| 5 | 9.9 | 322,297 | 353 | 191(100), 179(46) | Caffeoylquinic acid der. | ||
| 6 | 12.5 | 310 | 337 | 163(100), 173(4), 119(4), 191(6) | Coumaroylquinic acid der. 1 | ||
| 7 | 13.0 | 318 | 353 | 191(100), 179(6) | Cryptoclorogenic acid | ||
| 8 | 13.7 | 322,248 | 367 | 193(100), 134(5), 173(4) | Feruloylquinic acid der. 1 | ||
| 9 | 14.0 | 327,252 | 355 | 193(100), 175(30) | Ferulic acid hexoside | ||
| 10 | 16.2 | 267, 311 | 677 | 502(100), 503(93), 323(13) | [502] 240(100), 191(74), 163(58), 173(31); [323] 179(100) | [163] 119(100), 163; [179] 135(100) | p-coumaric-caffeoylquinic acid der. |
| 11 | 16.5 | 311 | 337 | 191(100), 163(6), 173(6) | Coumaroylquinic acid der. 2 | ||
| 12 | 18 | 273 | 705 | 531(100), 357(21) | [531] 357(100), 269(42), 313(16) | [357] 313(100), 193(13), 163(12) | Feruloylquinic acid der. 2 |
| 13 | 18.0 | 271 | 367 | 193(100), 173(8) | Ferulic acid der. | ||
| 14 | 18.2 | 304 | 337 | 191(100), 163(6) | Coumaroylquinic acid der. 4 | ||
| Flavonols & flavones | |||||||
| 15 | 10.41 | 374 | 593 | 285(100), 284(29) | Kaempferol-3-O-rutinoside | ||
| 16 | 15.6 | 377 | 755 | 593(100) | [593] 285(100) | Luteolin-7-rutinoside glucoside | |
| 17 | 17.3 | 350 | 755 | 593(100), 300(48), 301(23), 271(9) | Quercetin hexoside dirhamnoside | ||
| 18 | 19.0 | 353 | 609 | 300(100), 301(27), 271(15), 179(4) | Quercetin-3-rutinoside 1 | ||
| 19 | 19.5 | 355 | 635 | 284(100), 285(30), 255(28) | Kaempferol der. 2 | ||
| 20 | 19.9 | 347 | 593 | 285(100) | [285] 285(100), 241(33), 175(24), 199(21), 217(20), 243(20) | Luteolin-7-rutinoside | |
| 21 | 20.1 | 351 | 609 | 301(100), 300(22), 179(2) | Quercetin-3-rutinoside 2 | ||
| 22 | 20.1 | 350 | 1338 | 497(100), 659(89) | [497] 261(100) | Tanghenioside VII | |
| 23 | 20.4 | 330,350 | 623 | 315(100), 300(51) | Isorhamnetin-3-O-rutinoside 1 | ||
| 24 | 20.6 | 351 | 623 | 314(100), 315(96), 299(32) | Isorhamnetin-3-O-rutinoside 2 | ||
| 25 | 21.07 | 351 | 463 | 301(100), 300(24), 179(2) | Quercetin hexoside | ||
| 26 | 21.8 | 352 | 623 | 315(100), 300(17) | Isorhamnetin-3-O-rutinoside 3 | ||
| 27 | 21.9 | 352 | 623 | 315(100), 300(63), 271(5) | Isorhamnetin-3-O-rutinoside 4 | ||
| 28 | 22.0 | 344 | 653 | 611(100), 301(32), 300(29), 271(6) | Quercetin der. 1 | ||
| 29 | 22.3 | 345 | 608 | 300(100), 299(81), 285(25), 284(23) | [300] 285(100), 284(7); [299] 284(100) | [284] 284(100), 256(11) | Kaempferol dihexoside |
| 30 | 22.6 | 353 | 549 | 505(100), 405(45), 345(11) | [505] 301(100), 300(22) | Quercetin malonyl-hexoside der. 1 | |
| 31 | 22.6 | 353 | 1099 | 505(100), 549(25) | Quercetin malonyl-hexoside dimer | ||
| 32 | 23.7 | 350 | 505 | 301(100), 300(63), 179(3) | Quercetin der. 2 | ||
| 33 | 24.6 | 353,355 | 563 | 531(100), 463(76) | [531] 463(100); [463] 301(100), 300(14), 179(1) | Isorhamnetin malonyl-glucoside | |
| 34 | 24.6 | 353,355 | 1127 | 519(100) | 315(100), 300(12) | Isorhamnetin der. 1 | |
| 35 | 24.6 | 353,355 | 519 | 314(100), 315(63) | Isorhamnetin der. 2 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grohar, M.C.; Medic, A.; Ivancic, T.; Veberic, R.; Jogan, J. Color Variation and Secondary Metabolites’ Footprint in a Taxonomic Complex of Phyteuma sp. (Campanulaceae). Plants 2022, 11, 2894. https://doi.org/10.3390/plants11212894
Grohar MC, Medic A, Ivancic T, Veberic R, Jogan J. Color Variation and Secondary Metabolites’ Footprint in a Taxonomic Complex of Phyteuma sp. (Campanulaceae). Plants. 2022; 11(21):2894. https://doi.org/10.3390/plants11212894
Chicago/Turabian StyleGrohar, Mariana Cecilia, Aljaz Medic, Tea Ivancic, Robert Veberic, and Jernej Jogan. 2022. "Color Variation and Secondary Metabolites’ Footprint in a Taxonomic Complex of Phyteuma sp. (Campanulaceae)" Plants 11, no. 21: 2894. https://doi.org/10.3390/plants11212894
APA StyleGrohar, M. C., Medic, A., Ivancic, T., Veberic, R., & Jogan, J. (2022). Color Variation and Secondary Metabolites’ Footprint in a Taxonomic Complex of Phyteuma sp. (Campanulaceae). Plants, 11(21), 2894. https://doi.org/10.3390/plants11212894