

Asplenium pseudocapillipes (Aspleniaceae), a New Fern Species from South Korea



Abstract
:1. Introduction
2. Results
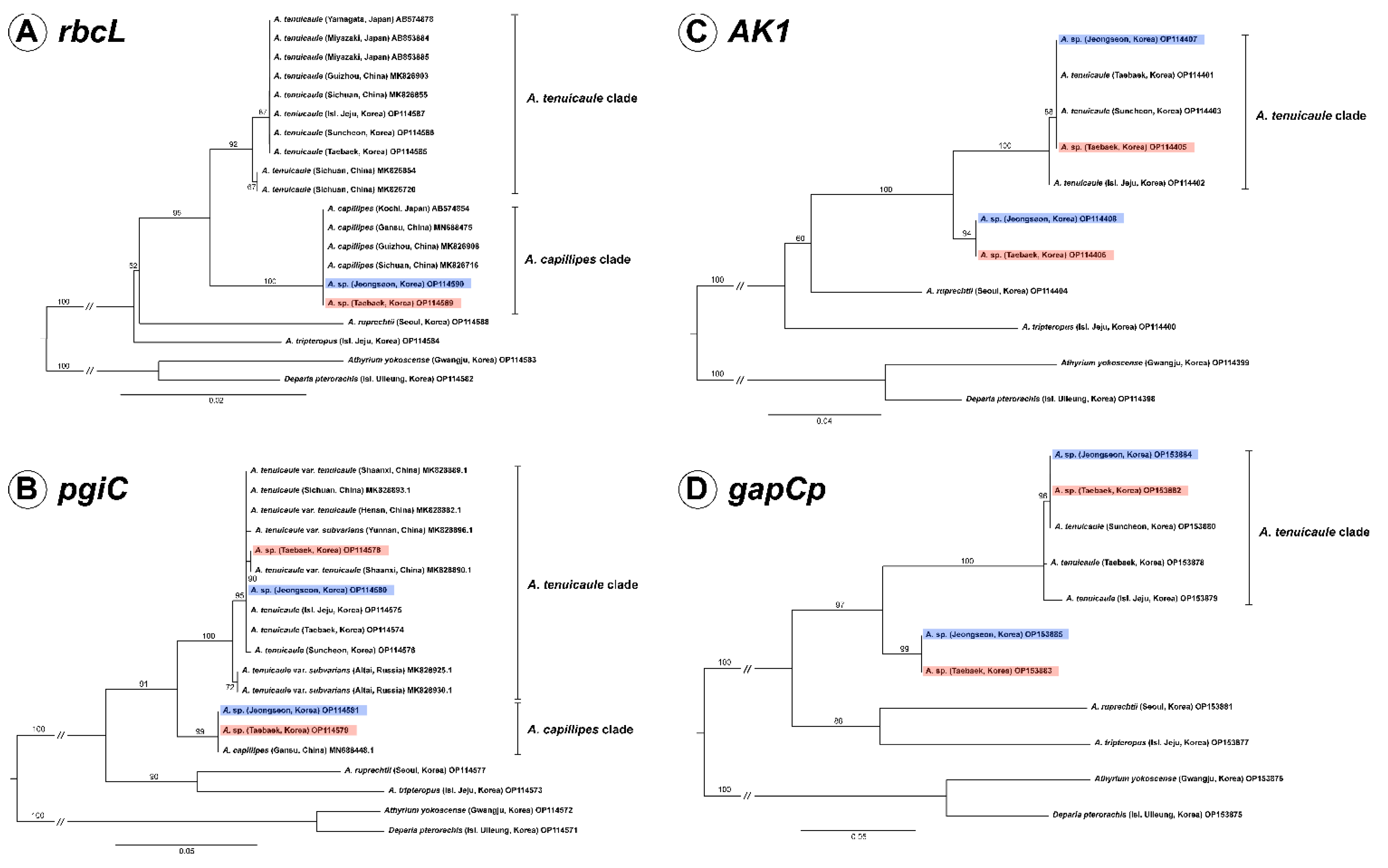
2.1. Chloroplast Phylogeny Based on rbcl
2.2. Nuclear Phylogenies Based on pgiC, AK1, and gapCp
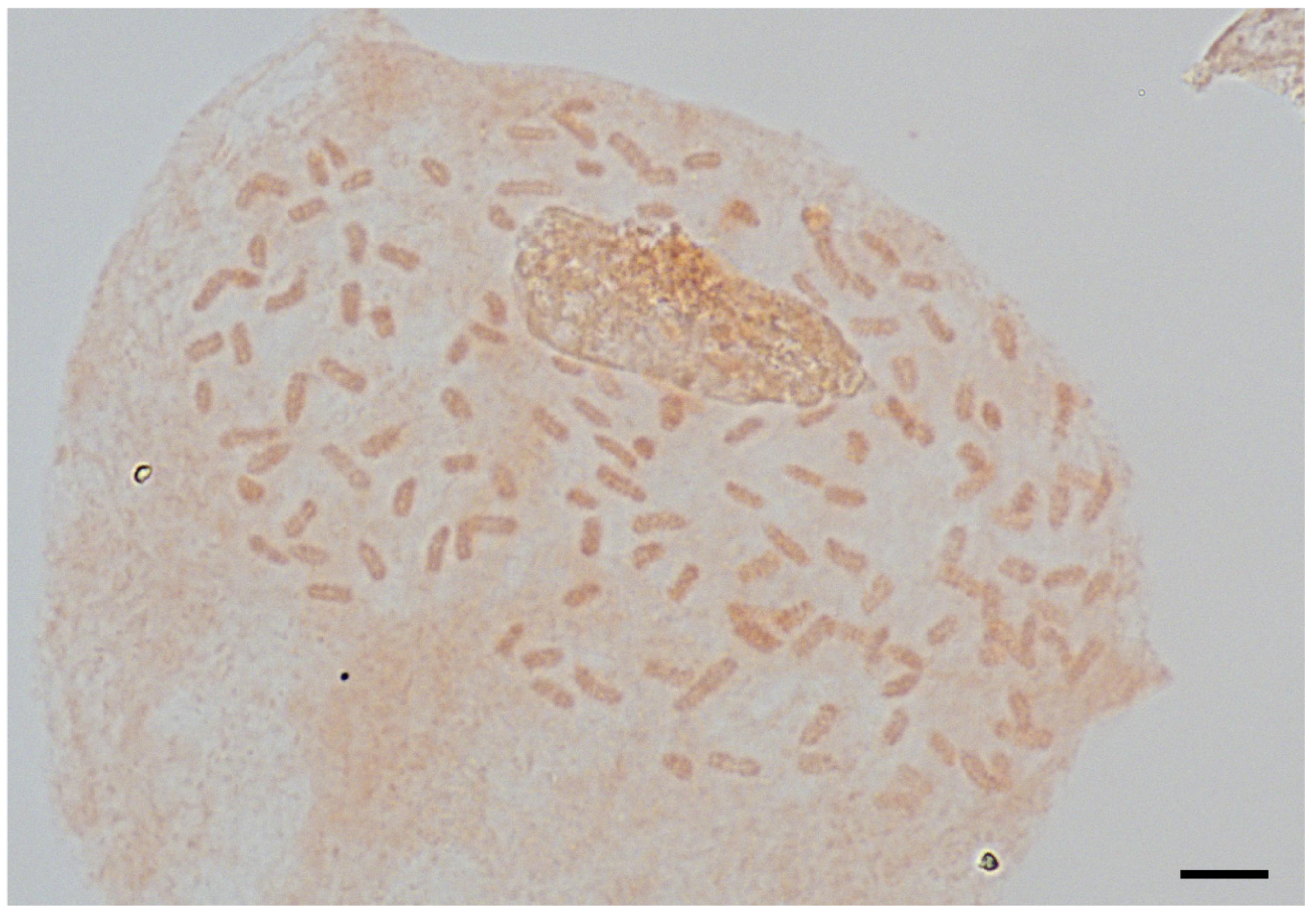
2.3. Ploidy Analysis and Spore Observation
2.4. Taxonomic Treatment
3. Discussion
3.1. Discovering New Allotetraploid in Genus Asplenium
3.2. Reproductive Mode of Asplenium Pseudocapillipes
4. Materials and Methods
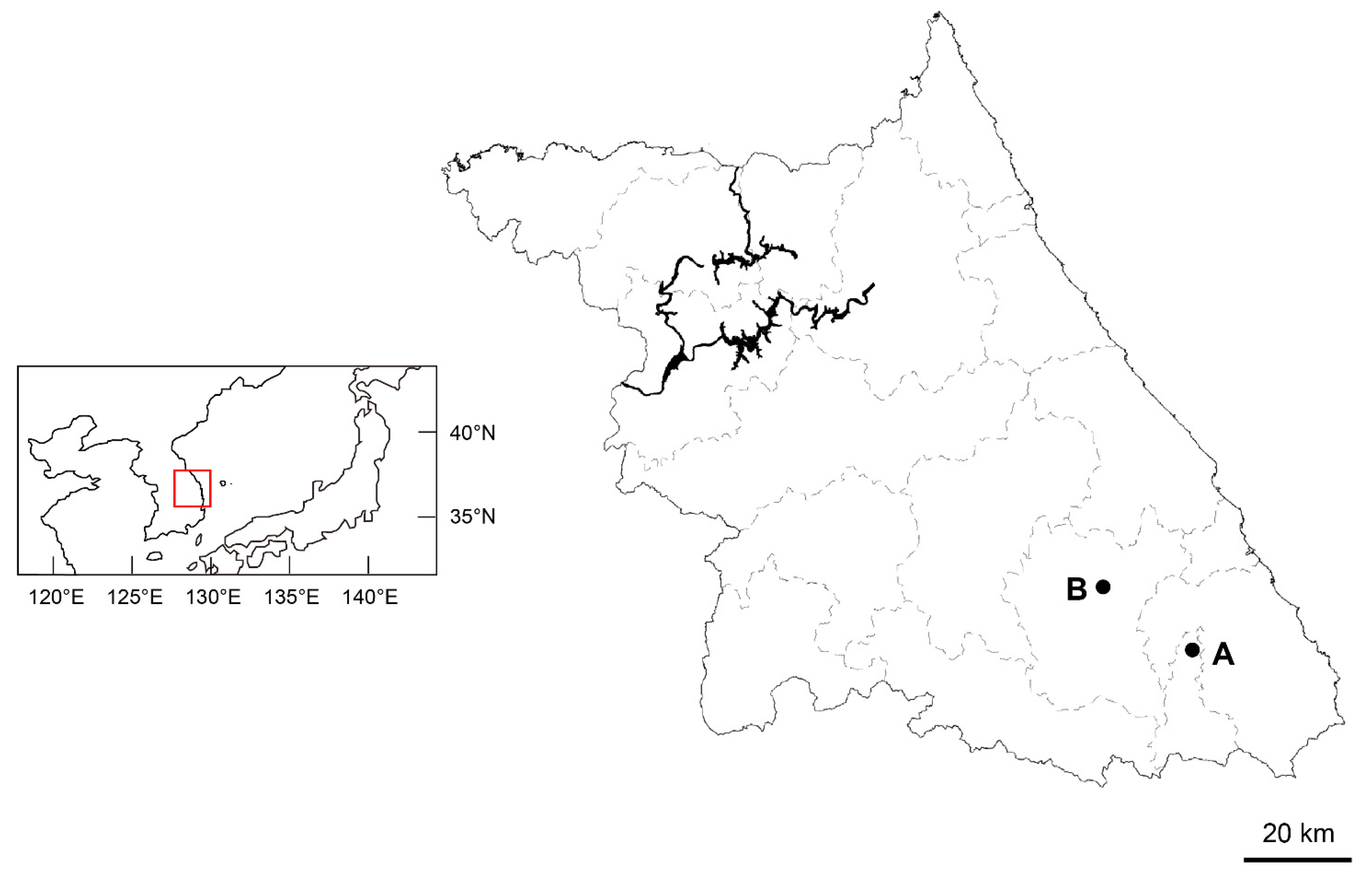
4.1. Plant Materials and Observation of Morphological Characteristics
4.2. Chromosome Counting and Measurement of the Genome Size
4.3. DNA Extraction, PCR Amplification, and Cloning
4.4. Sequence Alignments and Phylogenetic Analyses
4.5. Genome Size Measurement
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Madlung, A. Polyploidy and its effect on evolutionary success: Old questions revisited with new tools. Heredity (Edinburgh) 2013, 110, 99–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otto, S.P. The evolutionary consequences of polyploidy. Cell 2007, 131, 452–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soltis, P.S.; Soltis, D.E. The role of hybridization in plant speciation. Annu. Rev. Plant Biol. 2009, 60, 561–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soltis, P.S.; Marchant, D.B.; Van de Peer, Y.; Soltis, D.E. Polyploidy and genome evolution in plants. Curr. Opin. Genet. Dev. 2015, 35, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Stebbins , G.L., Jr. Types of polyploids: Their classification and significance. Adv. Genet. 1947, 1, 403–429. [Google Scholar]
- Wood, T.E.; Takebayashi, N.; Barker, M.S.; Mayrose, I.; Greenspoon, P.B.; Rieseberg, L.H. The frequency of polyploid speciation in vascular plants. Proc. Natl. Acad. Sci. USA 2009, 106, 13875–13879. [Google Scholar] [CrossRef] [Green Version]
- Linder, C.R.; Rieseberg, L.H. Reconstructing patterns of reticulate evolution in plants. Am. J. Bot. 2004, 91, 1700–1708. [Google Scholar] [CrossRef] [Green Version]
- Barrington, D.S.; Haufler, C.H.; Werth, C.R. Hybridization, Reticulation, and Species Concepts in the Ferns. Am. Fern J. 1989, 79, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.; Ebihara, A.; Lu, S.; Liu, H.; Schneider, H. Integrated taxonomy of the Asplenium normale complex (Aspleniaceae) in China and adjacent areas. J. Plant Res. 2018, 131, 573–587. [Google Scholar] [CrossRef]
- Dyer, R.J.; Savolainen, V.; Schneider, H. Apomixis and reticulate evolution in the Asplenium monanthes fern complex. Ann. Bot. 2012, 110, 1515–1529. [Google Scholar] [CrossRef] [Green Version]
- Ebihara, A.; Ishikawa, H.; Matsumoto, S.; Lin, S.J.; Iwatsuki, K.; Takamiya, M.; Watano, Y.; Ito, M. Nuclear DNA, chloroplast DNA, and ploidy analysis clarified biological complexity of the Vandenboschia radicans complex (Hymenophyllaceae) in Japan and adjacent areas. Am. J. Bot. 2005, 92, 1535–1547. [Google Scholar] [CrossRef] [PubMed]
- Hori, K.; Tono, A.; Fujimoto, K.; Kato, J.; Ebihara, A.; Watano, Y.; Murakami, N. Reticulate evolution in the apogamous Dryopteris varia complex (Dryopteridaceae, subg. Erythrovariae, sect. Variae) and its related sexual species in Japan. J. Plant Res. 2014, 127, 661–684. [Google Scholar] [PubMed]
- Hori, K. Hybrid origin of some species in the Deparia okuboana complex (Athyriaceae, Polypodiidae) verified with DNA analysis. Hikobia 2018, 17, 315–320. [Google Scholar]
- Liang, S.; Zhang, X.; Wei, R. Integrative taxonomy resolved species delimitation in a fern complex: A case study of the Asplenium coenobiale complex. Biodivers. Sci. 2019, 27, 1205–1220. [Google Scholar]
- Liang, S.Q.; Viane, R.L.; Zhang, X.C.; Wei, R. Exploring the reticulate evolution in the Asplenium pekinense complex and the A. varians complex (Aspleniaceae). J. Syst. Evol. 2021, 59, 125–140. [Google Scholar] [CrossRef]
- Ohlsen, D.J.; Perrie, L.R.; Shepherd, L.D.; Brownsey, P.J.; Bayly, M.J. Investigation of species boundaries and relationships in the Asplenium paleaceum complex (Aspleniaceae) using AFLP fingerprinting and chloroplast and nuclear DNA sequences. Aust. Syst. Bot. 2015, 27, 378–394. [Google Scholar] [CrossRef] [Green Version]
- Ebihara, A.; Matsumoto, S.; Ito, M. Taxonomy of the reticulate Vandenboschia radicans complex (Hymenophyllaceae) in Japan. Acta Phytotaxon. Geobot. 2009, 60, 26–40. [Google Scholar]
- Lin, Y.X.; Viane, R. Aspleniaceae; Wu, Z.Y., Raven, P.H., Hong, D.Y., Eds.; Flora of China Vol. 2–3 (Pteridophytes); Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 2013; pp. 267–316. [Google Scholar]
- PPGI. A community-derived classification for extant lycophytes and ferns. J. Syst. Evol. 2016, 54, 563–603. [Google Scholar] [CrossRef]
- Schneider, H.; Liu, H.M.; Chang, Y.F.; Ohlsen, D.; Perrie, L.R.; Shepherd, L.; Kessler, M.; Karger, D.N.; Hennequin, S.; Marquardt, J. Neo-and Paleopolyploidy contribute to the species diversity of Asplenium—The most species-rich genus of ferns. J. Syst. Evol. 2017, 55, 353–364. [Google Scholar] [CrossRef] [Green Version]
- Trewick, S.; Morgan-Richards, M.; Russell, S.; Henderson, S.; Rumsey, F.; Pinter, I.; Barrett, J.; Gibby, M.; Vogel, J. Polyploidy, phylogeography and Pleistocene refugia of the rockfern Asplenium ceterach: Evidence from chloroplast DNA. Mol. Ecol. 2002, 11, 2003–2012. [Google Scholar] [CrossRef]
- Chang, Y.; Li, J.; Lu, S.; Schneider, H. Species diversity and reticulate evolution in the Asplenium normale complex (Aspleniaceae) in China and adjacent areas. Taxon 2013, 62, 673–687. [Google Scholar] [CrossRef]
- Yatabe, Y.; Shinohara, W.; Matsumoto, S.; Murakami, N. Patterns of hybrid formation among cryptic species of bird-nest fern, Asplenium nidus complex (Aspleniaceae), in West Malesia. Bot. J. Linn. Soc. 2009, 160, 42–63. [Google Scholar] [CrossRef] [Green Version]
- Ichihara, Y. Ploidy level and reproductive mode of Asplenium capillipes (Aspleniaceae). J. Phytogeogr. Taxon. 2021, 69, 249–253. [Google Scholar] [CrossRef]
- Fraser-Jenkins, C.R. Taxonomic Revision of Three Hundred Indian Subcontinental Pteridophytes: With a Revised Census List; a New Picture of Fern-Taxonomy and Nomenclature in the Indian Subcontinent; Bishen Singh Mahendra Pal Singh: Dehradun, India, 2008. [Google Scholar]
- Ebihara, A. The Standard of Ferns and Lycophytes in Japan, Vol. 1; Gakken Publishers: Tokyo, Japan, 2016. [Google Scholar]
- BIR, S. Biosystematic studies on the asplenioid and athyrioid genera from the Himalayas: Results of chromosomal analysis on population basis. Indian Fern J. 1994, 11, 82–88. [Google Scholar]
- Walker, T. The cytogenetics of ferns. In The Experimental Biology of Ferns; Dyer, A.F., Ed.; Academic Press: London, UK; New York, NY, USA, 1979; pp. 87–132. [Google Scholar]
- Döpp, W. Die Apogamie bei Aspidium remotum Al. Br. Planta 1932, 17, 86–152. [Google Scholar] [CrossRef]
- Manton, I. Problems of Cytology and Evolution in the Pteridophyta; Cambridge University Press: London, UK, 1950. [Google Scholar]
- Braithwaite, A. A new type of apogamy in ferns. New Phytol. 1964, 63, 293–305. [Google Scholar] [CrossRef]
- Knobloch, I.W. A Preliminary Review of Spore Number and Apogamy within the Genus Cheilanthes. Am. Fern J. 1966, 56, 163–167. [Google Scholar] [CrossRef]
- Lin, S.-J.; Kato, M.; Iwatsuki, K. Sporogenesis, reproductive mode, and cytotaxonomy of some species of Sphenomeris, Lindsaea, and Tapeinidium (Lindsaeaceae). Am. Fern J. 1990, 80, 97–109. [Google Scholar] [CrossRef]
- Gastony, G.J. Spore morphology in the Dicksoniaceae. I. The genera Cystodium, Thyrsopteris, and Culcita. Am. J. Bot. 1981, 68, 808–819. [Google Scholar] [CrossRef]
- Huang, Y.-M.; Chiou, W.-L.; Lee, P.-H. Morphology of the gametophytes and young sporophytes of Cyatheaceae native to Taiwan. TAIWANIA-TAIPEI- 2001, 46, 274–283. [Google Scholar]
- Regalado, L.; Sánchez, C. Spore morphology as a taxonomic tool in the delimitation of three Asplenium L. species complexes (Aspleniaceae: Pteridophyta) in Cuba. Grana 2002, 41, 107–113. [Google Scholar] [CrossRef]
- Morzenti, V.M. Morphological and Cytological Data on Southeastern United States Species of the Asplenium heterochroum-resiliens Complex. Am. Fern J. 1966, 56, 167–177. [Google Scholar] [CrossRef]
- Mehra, P.N.; Bir, S.S. Cytological Observations on <i>Asplenium cheilosorum</i> Kunze. Cytologia 1960, 25, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Mitui, K. Chromosome studies on Japanese ferns (4). J. Jap. Bot. 1970, 45, 84–90. [Google Scholar]
- De Groot, G.A.; During, H.J.; Maas, J.W.; Schneider, H.; Vogel, J.C.; Erkens, R.H. Use of rbcL and trnL-F as a two-locus DNA barcode for identification of NW-European ferns: An ecological perspective. PLoS ONE 2011, 6, e16371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuettpelz, E.; Pryer, K.M. Fern phylogeny inferred from 400 leptosporangiate species and three plastid genes. Taxon 2007, 56, 1037–1050. [Google Scholar]
- Hori, K.; Okuyama, Y.; Watano, Y.; Murakami, N. Recurrent hybridization without homoeologous chromosome paring in the Dryopteris varia complex (Dryopteridaceae). Chromosome Bot. 2018, 13, 9–24. [Google Scholar] [CrossRef] [Green Version]
- Schuettpelz, E.; Grusz, A.L.; Windham, M.D.; Pryer, K.M. The utility of nuclear gapCp in resolving polyploid fern origins. Syst. Bot. 2008, 33, 621–629. [Google Scholar] [CrossRef]
- Ishikawa, H.; Watano, Y.; Kano, K.; Ito, M.; Kurita, S. Development of primer sets for PCR amplification of the PgiC gene in ferns. J. Plant Res. 2002, 115, 0065–0070. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Misawa, K.; Kuma, K.i.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Valkonen, J.; Watanabe, K.; Pehu, E. Analysis of correlation between nuclear DNA content, chromosome number, and flowering capacity of asymmetric somatic hybrids of diploid Solanum brevidens and (di) haploid S. tuberosum. Jpn. J. Genet. 1994, 69, 525–536. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A. capillipes | A. pseudocapillipes | A. tenuicaule | |
|---|---|---|---|
| Plant height (cm) | 3–8 | 3–9 | 6–15 |
| Stipe (cm) | 0.3–2.5 | 0.5–3 | 1.5–5 |
| Lamina (cm) | 1.5–6 × 0.5–2.5 | 1.5–6 × 0.7–2.5 | 4–9 × 1.2–3 |
| Number of pinnae | 5–7 | 4–9 | 7–10 |
| Spore length (μm) | 38–42 45.7 * | 46–59 | 28–32 29.1 * |
| Ploidy | 2×, 4× | 4× | 2× |
| Number of spores per sporangium | 32 | 32 | 64 |
| Gemma on rachis | Present | Absent | Absent |
| Voucher | Locality | Accession | |||
|---|---|---|---|---|---|
| rbcL | AK1 | gapCp | pgiC | ||
| Asplenium sp. | |||||
| CBNU2021-0084-A | Korea: Gangwon-do, Taebaek-si | OP114589 | OP114405 P114406 | OP153882 P153883 | OP114578 P114579 |
| CBNU2020-0171-A | Korea: Gangwon-do, Jeongsun-gun | OP114590 | OP114407 P114408 | OP153884 P153885 | OP114580 P114581 |
| Asplenium tenuicaule | |||||
| CBNU2020-0096-E | Korea: Jeollabuk-do, Suncheon-si | OP114586 | OP114403 | OP153880 | OP114576 |
| CBNU2020-0157-D | Korea: Jeju Island, Jeju-si | OP114587 | OP114402 | OP153879 | OP114575 |
| CBNU2021-0086-D | Korea: Gangwon-do, Taebaek-si | OP114585 | OP114401 | OP153878 | OP114574 |
| Asplenium ruprechtii | |||||
| CBNU2020-0180-A | Korea: Seoul, Gangbuk-gu | OP114588 | OP114404 | OP153881 | OP114577 |
| Asplenium tripteropus | |||||
| CBNU2020-0028-A | Korea: Jeju Island, Jeju-si | OP114584 | OP114400 | OP153877 | OP114573 |
| Athyrium yokoscense(outgroup) | |||||
| CBNU2021-0104-A | Korea: Jeollanam-do, Gwangju-si | OP114583 | OP114399 | OP153876 | OP114572 |
| Deparia pterorachis (outgroup) | |||||
| CBNU2021-0017-A | Korea: Gyeongsangbuk-do, Ulleung Island | OP114582 | OP114398 | OP153875 | OP114571 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.H.; Kim, J.S.; Kim, H.T. Asplenium pseudocapillipes (Aspleniaceae), a New Fern Species from South Korea. Plants 2022, 11, 3089. https://doi.org/10.3390/plants11223089
Park SH, Kim JS, Kim HT. Asplenium pseudocapillipes (Aspleniaceae), a New Fern Species from South Korea. Plants. 2022; 11(22):3089. https://doi.org/10.3390/plants11223089
Chicago/Turabian StylePark, Sang Hee, Jung Sung Kim, and Hyoung Tae Kim. 2022. "Asplenium pseudocapillipes (Aspleniaceae), a New Fern Species from South Korea" Plants 11, no. 22: 3089. https://doi.org/10.3390/plants11223089
APA StylePark, S. H., Kim, J. S., & Kim, H. T. (2022). Asplenium pseudocapillipes (Aspleniaceae), a New Fern Species from South Korea. Plants, 11(22), 3089. https://doi.org/10.3390/plants11223089