

Actinomycete Potential as Biocontrol Agent of Phytopathogenic Fungi: Mechanisms, Source, and Applications
 , , and
, , and
Abstract
:1. Introduction
2. General Characteristics of Actinomycetes
3. Actinomycetes as Biocontrol Agents
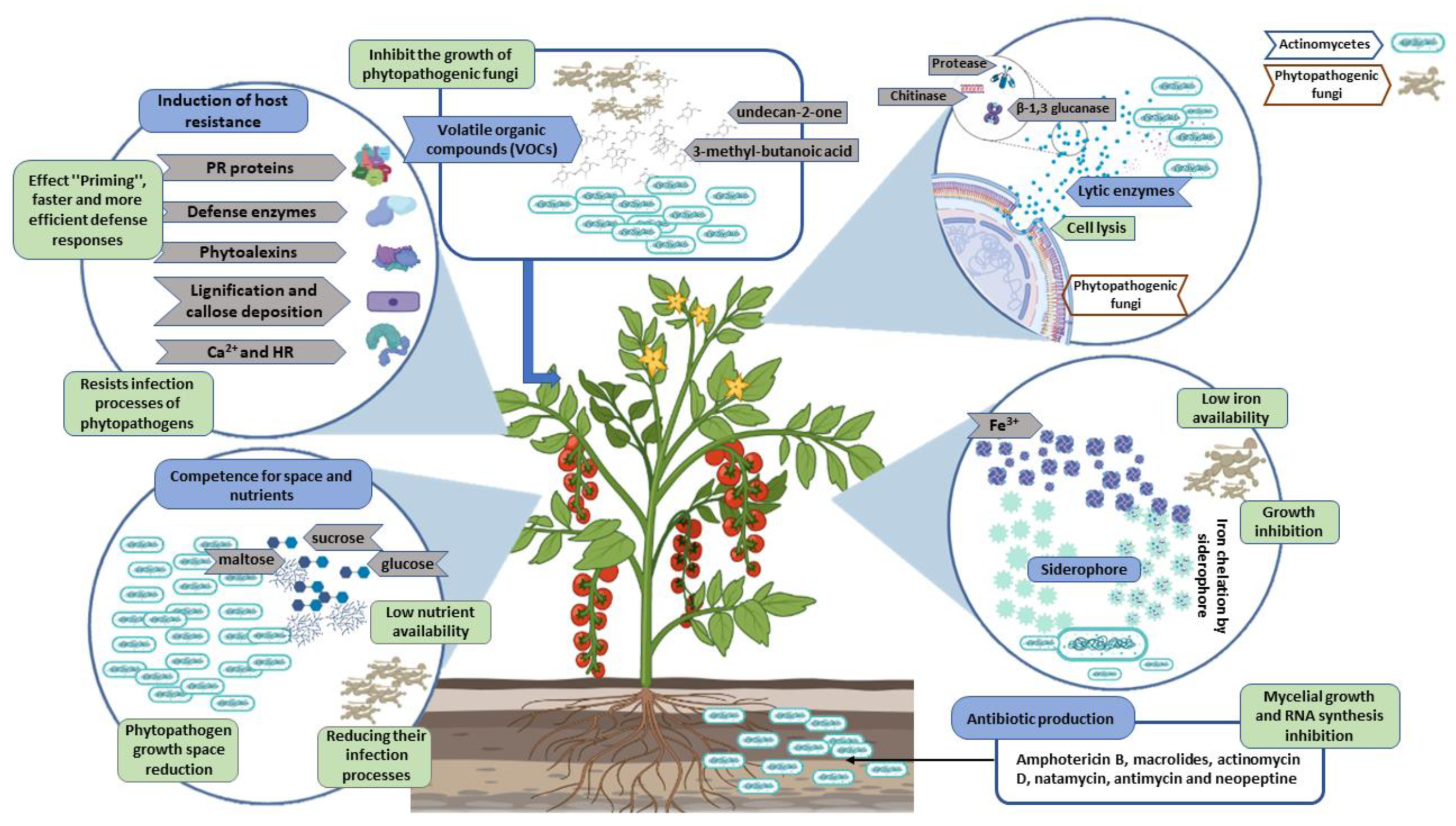
4. Main Actinomycete Antagonistic Mechanisms to Phytopathogenic Fungi
4.1. Competence for Space and Nutrients
4.2. Antibiotic Production
4.3. Siderophore Production
4.4. Lytic Enzyme Production
4.5. Volatile Organic Compounds (VOCs)
4.6. Induction of Host Resistance
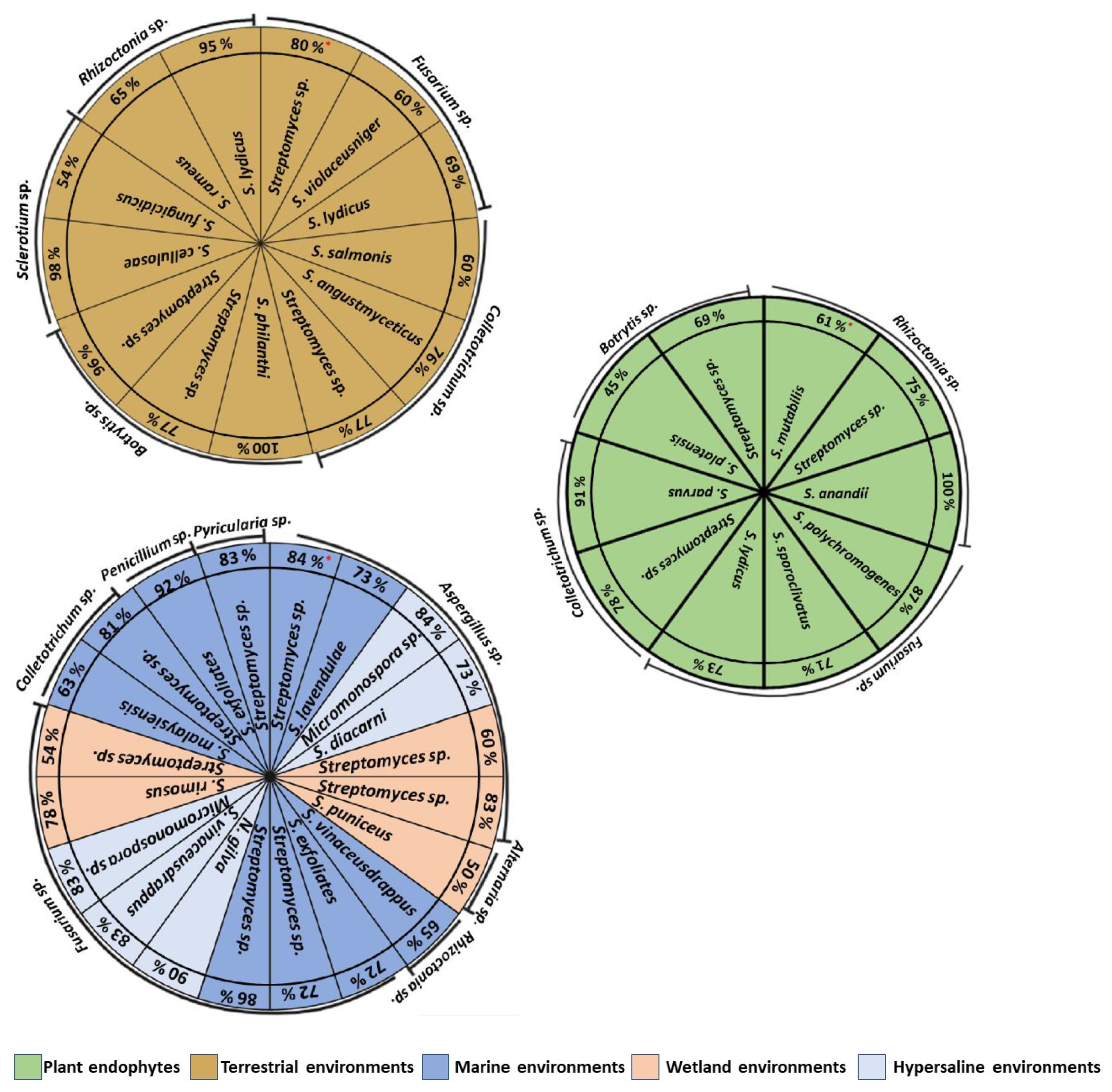
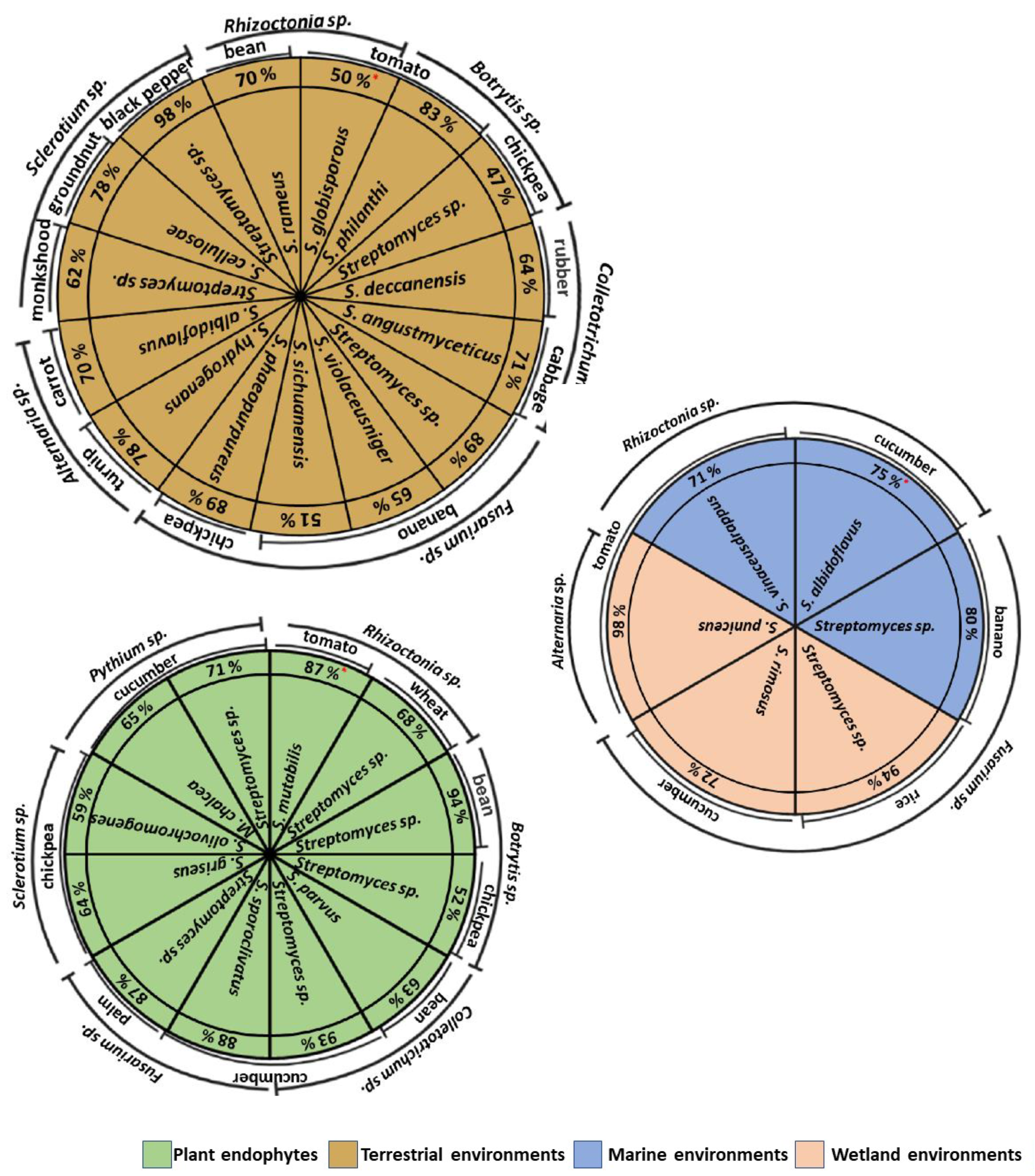
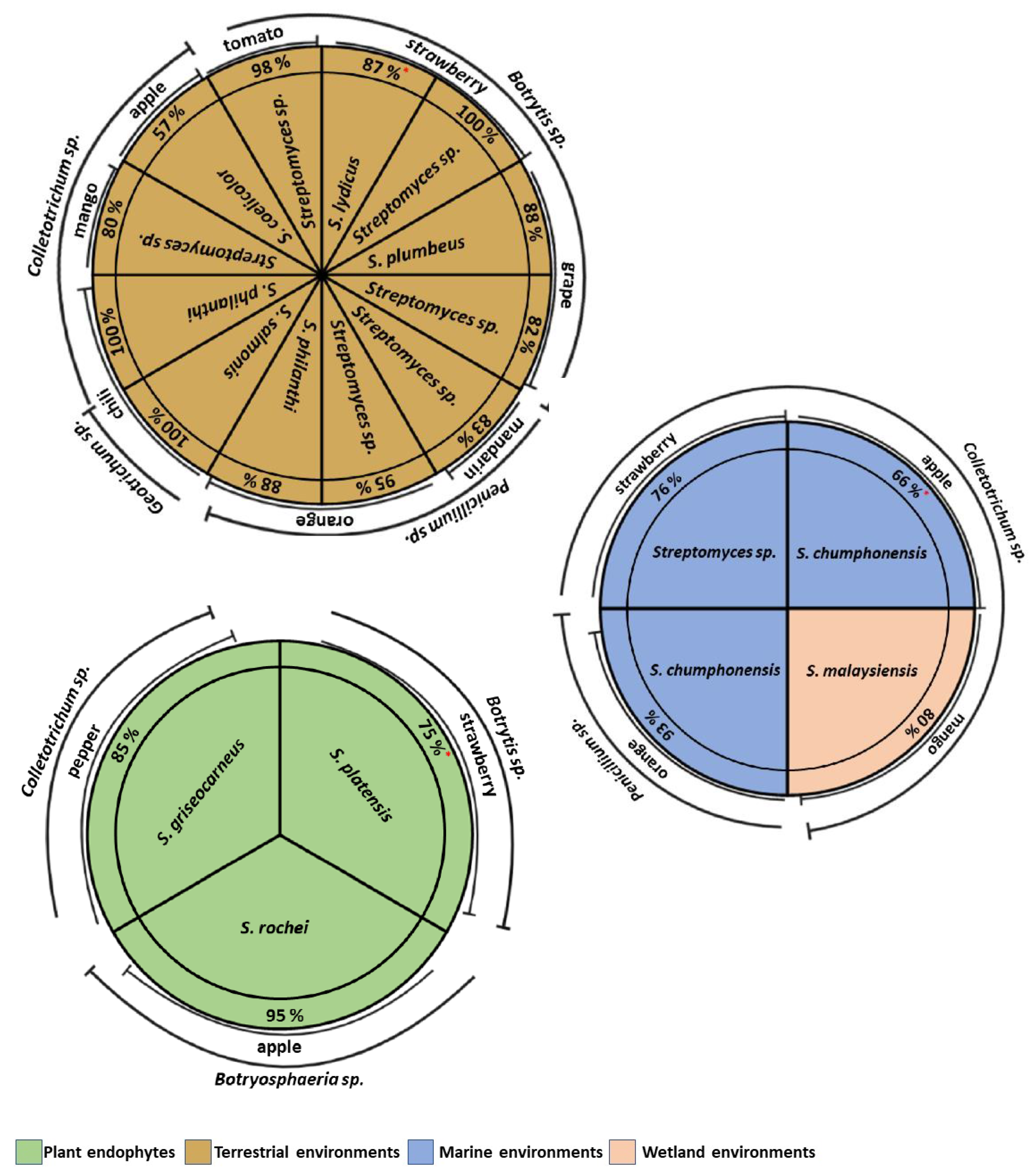
5. Actinomycete Isolation from Different Environments
6. Antifungal In Vitro Activity of Actinomycetes Isolated in Different Environments
7. Antifungal Activity of Actinomycetes In Vivo Isolated from Different Environments
8. Antifungal Activity of Actinomycetes Isolated in Different Environments in Postharvest Fruit

9. Commercial Products Based on Actinomycetes
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alblooshi, A.A.; Purayil, G.P.; Saeed, E.E.; Ramadan, G.A.; Tariq, S.; Altaee, A.S.; El-Tarabily, K.A.; AbuQamar, S.F. Biocontrol potential of endophytic actinobacteria against Fusarium solani, the causal agent of sudden decline syndrome on date palm in the UAE. J. Fungi 2022, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Torres-Rodriguez, J.A.; Reyes-Pérez, J.J.; Castellanos, T.; Angulo, C.; Quiñones-Aguilar, E.E.; Hernandez-Montiel, L.G. A biopolymer with antimicrobial properties and plant resistance inducer against phytopathogens: Chitosan. Not. Bot. Horti Agrobot. Cluj-Napoca 2021, 49, 12231. [Google Scholar] [CrossRef]
- Xiao, L.; Niu, H.-J.; Qu, T.-L.; Zhang, X.-F.; Du, F.-Y. Streptomyces sp. FX13 inhibits fungicide-resistant Botrytis cinerea in vitro and in vivo by producing oligomycin A. Pestic. Biochem. Physiol. 2021, 175, 104834. [Google Scholar] [CrossRef] [PubMed]
- Boukaew, S.; Yossan, S.; Cheirsilp, B.; Prasertsan, P. Impact of environmental factors on Streptomyces spp. metabolites against Botrytis cinerea. J. Basic Microbiol. 2022, 62, 611–622. [Google Scholar] [CrossRef]
- Jagannathan, S.V.; Manemann, E.M.; Rowe, S.E.; Callender, M.C.; Soto, W. Marine actinomycetes, new sources of biotechnological products. Mar. Drugs 2021, 19, 365. [Google Scholar] [CrossRef]
- Tatar, D. Isolation, phylogenetic analysis and antimicrobial activity of halophilic actinomycetes from different saline environments located near Çorum province. Biologia 2021, 76, 773–780. [Google Scholar] [CrossRef]
- Maua, J.O.; Mbuvi, M.T.E.; Matiku, P.; Munguti, S.; Mateche, E.; Owili, M. The difficult choice-to conserve the living filters or utilizing the full potential of wetlands: Insights from the Yala swamp, Kenya. Environ. Chall. 2022, 6, 100427. [Google Scholar] [CrossRef]
- Fadhilah, Q.G.; Santoso, I.; Maryanto, A.E.; Abdullah, S.; Yasman, Y. Evaluation of the antifungal activity of marine actinomycetes isolates against the phytopathogenic fungi Colletotrichum siamense KA: A preliminary study for new antifungal compound discovery. Pharmacia 2021, 68, 837–843. [Google Scholar] [CrossRef]
- Xia, X.; Liu, J.; Huang, L.; Zhang, X.; Deng, Y.; Li, F.; Liu, Z.; Huang, R. Molecular details of actinomycin D-Treated MRSA revealed via high-dimensional data. Mar. Drugs 2022, 20, 114. [Google Scholar] [CrossRef]
- Meena, L.I.; Rajeswari, E.; Ahiladevi, P.; Kamalakannan, A.; Kalaiselvi, T. Antifungal potential of Streptomyces rameus GgS 48 against mungbean root rot [Rhizoctonia bataticola (Taub.) Butler]. J. Biosci. 2022, 47, 10. [Google Scholar] [CrossRef] [PubMed]
- Gebily, D.A.S.; Ghanem, G.A.M.; Ragab, M.M.; Ali, A.M.; Soliman, N.E.K.; Abd El-Moity, T.H. Characterization and potential antifungal activities of three Streptomyces spp. as biocontrol agents against Sclerotinia sclerotiorum (Lib.) de Bary infecting green bean. Egypt. J. Biol. Pest Control 2021, 31, 33. [Google Scholar] [CrossRef]
- Gong, Y.; Liu, J.-Q.; Xu, M.-J.; Zhang, C.-M.; Gao, J.; Li, C.-G.; Xing, K.; Qin, S. Antifungal volatile organic compounds from Streptomyces setonii WY228 control black spot disease of sweet potato. Appl. Environ. Microbiol. 2022, 88, e02317-21. [Google Scholar] [CrossRef] [PubMed]
- Kaari, M.; Joseph, J.; Manikkam, R.; Sreenivasan, A.; Venugopal, G.; Alexander, B.; Krishnan, S. Anti-Biofilm activity and biocontrol potential of streptomyces cultures against Ralstonia solanacearum on tomato plants. Indian J. Microbiol. 2022, 62, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Mitra, D.; Mondal, R.; Khoshru, B.; Senapati, A.; Radha, T.K.; Mahakur, B.; Uniyal, N.; Myo, E.M.; Boutaj, H.; Sierra, B.E.G.; et al. Actinobacteria-enhanced plant growth, nutrient acquisition, and crop protection: Advances in soil, plant, and microbial multifactorial interactions. Pedosphere 2022, 32, 149–170. [Google Scholar] [CrossRef]
- van Bergeijk, D.A.; Terlouw, B.R.; Medema, M.H.; van Wezel, G.P. Ecology and genomics of actinobacteria: New concepts for natural product discovery. Nat. Rev. Microbiol. 2020, 18, 546–558. [Google Scholar] [CrossRef]
- Cheng, Z.; McCann, S.; Faraone, N.; Clarke, J.-A.; Hudson, E.A.; Cloonan, K.; Hillier, N.K.; Tahlan, K. Production of plant-associated volatiles by select model and industrially important Streptomyces spp. Microorganisms 2020, 8, 1767. [Google Scholar] [CrossRef] [PubMed]
- Vurukonda, S.S.K.P.; Giovanardi, D.; Stefani, E. Plant growth promoting and biocontrol activity of Streptomyces spp. as endophytes. Int. J. Mol. Sci. 2018, 19, 952. [Google Scholar] [CrossRef] [Green Version]
- Gomes, E.d.B.; Dias, L.R.L.; Miranda, R.d.C.M. Actinomycetes bioactive compounds: Biological control of fungi and phytopathogenic insect. Afr. J. Biotechnol. 2018, 17, 552–559. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Jing, T.; Zhou, D.; Zhang, M.; Qi, D.; Zang, X.; Zhao, Y.; Li, K.; Tang, W.; Chen, Y.; et al. Biocontrol efficacy and possible mechanism of Streptomyces sp. H4 against postharvest anthracnose caused by Colletotrichum fragariae on strawberry fruit. Postharvest Biol. Technol. 2021, 175, 111401. [Google Scholar] [CrossRef]
- Zhou, D.; Jing, T.; Chen, Y.; Yun, T.; Qi, D.; Zang, X.; Zhang, M.; Wei, Y.; Li, K.; Zhao, Y.; et al. Biocontrol potential of a newly isolated Streptomyces sp. HSL-9B from mangrove forest on postharvest anthracnose of mango fruit caused by Colletotrichum gloeosporioides. Food Control 2022, 135, 108836. [Google Scholar] [CrossRef]
- Zou, N.; Zhou, D.; Chen, Y.; Lin, P.; Chen, Y.; Wang, W.; Xie, J.; Wang, M. A novel antifungal actinomycete Streptomyces sp. strain H3-2 effectively controls banana Fusarium wilt. Front. Microbiol. 2021, 12, 706647. [Google Scholar] [CrossRef]
- Tiwari, K.; Gupta, R.K. Diversity and isolation of rare actinomycetes: An overview. Crit. Rev. Microbiol. 2013, 39, 256–294. [Google Scholar] [CrossRef] [PubMed]
- Javoreková, S.; Kovácsová, S.; Medo, J.; Maková, J.; Petrová, J.; Hleba, L.; Košťálová, D.; Cinkocki, R. Soil amended with organic fertilizers as a source of actinomycetes with high potential as biocontrol agents. J. Microbiol. Biotechnol. Food Sci. 2021, 8, 1352–1359. [Google Scholar] [CrossRef]
- Ascencion, L.C.; Liang, W.J.; Yen, T.B. Control of Rhizoctonia solani damping-off disease after soil amendment with dry tissues of Brassica results from increase in Actinomycetes population. Biol. Control 2015, 82, 21–30. [Google Scholar] [CrossRef]
- Qi, D.; Zou, L.; Zhou, D.; Zhang, M.; Wei, Y.; Li, K.; Zhao, Y.; Zhang, L.; Xie, J. Biocontrol potential and antifungal mechanism of a novel Streptomyces sichuanensis against Fusarium oxysporum f. sp. cubense tropical race 4 in vitro and in vivo. Appl. Microbiol. Biotechnol. 2022, 106, 1633–1649. [Google Scholar] [CrossRef]
- Cabrera, R.; García-López, H.; Aguirre-von-Wobeser, E.; Orozco-Avitia, J.A.; Gutiérrez-Saldaña, A.H. Amycolatopsis BX17: An actinobacterial strain isolated from soil of a traditional milpa agroecosystem with potential biocontrol against Fusarium graminearum. Biol. Control 2020, 147, 104285. [Google Scholar] [CrossRef]
- Ramlawi, S.; Abusharkh, S.; Carroll, A.; McMullin, D.R.; Avis, T.J. Biological and chemical characterization of antimicrobial activity in Arthrobacter spp. isolated from disease-suppressive compost. J. Basic Microbiol. 2021, 61, 745–756. [Google Scholar] [CrossRef]
- Allali, K.; Goudjal, Y.; Zamoum, M.; Bouznada, K.; Sabaou, N.; Zitouni, A. Nocardiopsis dassonvillei strain MB22 from the Algerian Sahara promotes wheat seedlings growth and potentially controls the common root rot pathogen Bipolaris sorokiniana. J. Plant Pathol. 2019, 101, 1115–1125. [Google Scholar] [CrossRef]
- Ebrahimi-Zarandi, M.; Bonjar, G.H.S.; Riseh, R.S.; El-Shetehy, M.; Saadoun, I.; Barka, E.A. Exploring two Streptomyces species to control Rhizoctonia solani in tomato. Agronomy 2021, 11, 1384. [Google Scholar] [CrossRef]
- Lahlali, R.; Ezrari, S.; Radouane, N.; Kenfaoui, J.; Esmaeel, Q.; El Hamss, H.; Belabess, Z.; Ait Barka, E. Biological control of plant pathogens: A global perspective. Microorganisms 2022, 10, 596. [Google Scholar] [CrossRef]
- Carmona-Hernandez, S.; Reyes-Pérez, J.J.; Chiquito-Contreras, R.G.; Rincon-Enriquez, G.; Cerdan-Cabrera, C.R.; Hernandez-Montiel, L.G. Biocontrol of postharvest fruit fungal diseases by bacterial antagonists: A review. Agronomy 2019, 9, 121. [Google Scholar] [CrossRef] [Green Version]
- Dutta, S.; Lee, Y.H. High-throughput identification of genes influencing the competitive ability to obtain nutrients and performance of biocontrol in Pseudomonas putida JBC17. Sci. Rep. 2022, 8, 14336. [Google Scholar] [CrossRef] [PubMed]
- Farda, B.; Djebaili, R.; Vaccarelli, I.; Del Gallo, M.; Pellegrini, M. Actinomycetes from caves: An overview of their diversity, biotechnological properties, and insights for their use in soil environments. Microorganisms 2022, 10, 453. [Google Scholar] [CrossRef] [PubMed]
- Köhl, J.; Kolnaar, R.; Ravensberg, W.J. Mode of action of microbial biological control agents against plant diseases: Relevance beyond efficacy. Front. Plant Sci. 2019, 10, 845. [Google Scholar] [CrossRef] [Green Version]
- Palla, M.S.; Guntuku, G.S.; Muthyala, M.K.K.; Pingali, S.; Sahu, P.K. Isolation and molecular characterization of antifungal metabolite producing actinomycete from mangrove soil. Beni-Suef Univ. J. Basic Appl. Sci. 2018, 7, 250–256. [Google Scholar] [CrossRef]
- Wang, H.; Tian, R.; Tian, Q.; Yan, X.; Huang, L.; Ji, Z. Investigation on the antifungal ingredients of Saccharothrix yanglingensis Hhs. 015, an antagonistic endophytic actinomycete isolated from cucumber plant. Molecules 2019, 24, 3686. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Lu, Y.; Chen, H.; Wu, C.; Zhang, H.; Chen, L.; Chen, X. Antifungal peptides produced by actinomycetes and their biological activities against plant diseases. J. Antibiot. 2020, 73, 265–282. [Google Scholar] [CrossRef]
- Polyak, Y.M.; Sukharevich, V.I. Detection and regulation of antagonistic properties of the soil actinomycete Streptomyces sp. 89. Biol. Bull. 2021, 48, 626–634. [Google Scholar] [CrossRef]
- Vázquez-Laslop, N.; Mankin, A.S. How macrolide antibiotics work. Trends Biochem. Sci. 2018, 43, 668–684. [Google Scholar] [CrossRef]
- Hartmann, D.O.; Shimizu, K.; Rothkegel, M.; Petkovic, M.; Ferraz, R.; Petrovski, Ž.; Branco, L.C.; Lopes, J.N.C.; Pereira, C.S. Tailoring amphotericin B as an ionic liquid: An upfront strategy to potentiate the biological activity of antifungal drugs. RSC Adv. 2021, 11, 14441–14452. [Google Scholar] [CrossRef]
- Young, D.H.; Wang, N.X.; Meyer, S.T.; Avila-Adame, C. Characterization of the mechanism of action of the fungicide fenpicoxamid and its metabolite UK-2A. Pest Manag. Sci. 2018, 74, 489–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meena, M.; Prajapati, P.; Ravichandran, C.; Sehrawat, R. Natamycin: A natural preservative for food applications-a review. Food Sci. Biotechnol. 2021, 30, 1481–1496. [Google Scholar] [CrossRef]
- Sharma, S.; Fulke, A.B.; Chaubey, A. Bioprospection of marine actinomycetes: Recent advances, challenges and future perspectives. Acta Oceanol. Sin. 2019, 38, 1–17. [Google Scholar] [CrossRef]
- Ghosh, S.K.; Bera, T.; Chakrabarty, A.M. Microbial siderophore–A boon to agricultural sciences. Biol. Control 2020, 144, 104214. [Google Scholar] [CrossRef]
- Roskova, Z.; Skarohlid, R.; McGachy, L. Siderophores: An alternative bioremediation strategy? Sci. Total Environ. 2022, 819, 153144. [Google Scholar] [CrossRef] [PubMed]
- Retamal-Morales, G.; Senges, C.H.R.; Stapf, M.; Olguín, A.; Modak, B.; Bandow, J.E.; Tischler, D.; Schlömann, M.; Levicán, G. Isolation and characterization of arsenic-binding siderophores from Rhodococcus erythropolis S43: Role of heterobactin B and other heterobactin variants. Appl. Microbiol. Biotechnol. 2021, 105, 1731–1744. [Google Scholar] [CrossRef]
- Wu, Q.; Deering, R.W.; Zhang, G.; Wang, B.; Li, X.; Sun, J.; Chen, J.; Zhang, H.; Rowley, D.C.; Wang, H. Albisporachelin, a new hydroxamate type siderophore from the deep ocean sediment-derived actinomycete Amycolatopsis albispora WP1T. Mar. Drugs 2018, 16, 199. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Montiel, L.G.; Rivas-García, T.; Romero-Bastidas, M.; Chiquito-Contreras, C.J.; Ruiz-Espinoza, F.H.; Chiquito-Contreras, R.G. Antagonistic potential of bacteria and marine yeasts for the control of phytopathogenic fungi. Rev. Mex. Cienc. Agríc. 2018, 9, 4311–4321. [Google Scholar] [CrossRef] [Green Version]
- Mottola, A.; Ramírez-Zavala, B.; Hünniger, K.; Kurzai, O.; Morschhäuser, J. The zinc cluster transcription factor Czf1 regulates cell wall architecture and integrity in Candida albicans. Mol. Microbiol. 2021, 116, 483–497. [Google Scholar] [CrossRef]
- Dimkić, I.; Janakiev, T.; Petrović, M.; Degrassi, G.; Fira, D. Plant-associated Bacillus and Pseudomonas antimicrobial activities in plant disease suppression via biological control mechanisms-A review. Physiol. Mol. Plant Pathol. 2022, 117, 101754. [Google Scholar] [CrossRef]
- Jadhav, H.P.; Shaikh, S.S.; Sayyed, R.Z. Role of hydrolytic enzymes of rhizoflora in biocontrol of fungal phytopathogens. An overview. In Rhizotrophs: Plant Growth Promotion to Bioremediation; Mehnaz, S., Ed.; Microorganisms for Sustainability; Springer: Singapore, 2017; pp. 183–203. ISBN 978-981-10-4862-3. [Google Scholar]
- del Rosario Cappellari, L.; Chiappero, J.; Banchio, E. Invisible signals from the underground: A practical method to investigate the effect of microbial volatile organic compounds emitted by rhizobacteria on plant growth. Biochem. Mol. Biol. Educ. 2019, 47, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Ayed, A.; Kalai-Grami, L.; Ben Slimene, I.; Chaouachi, M.; Mankai, H.; Karkouch, I.; Djebali, N.; Elkahoui, S.; Tabbene, O.; Limam, F. Antifungal activity of volatile organic compounds from Streptomyces sp. strain S97 against Botrytis cinerea. Biocontrol Sci. Technol. 2021, 31, 1330–1348. [Google Scholar] [CrossRef]
- Widada, J.; Damayanti, E.; Alhakim, M.R.; Yuwono, T.; Mustofa, M. Two strains of airborne Nocardiopsis alba producing different volatile organic compounds (VOCs) as biofungicide for Ganoderma boninense. FEMS Microbiol. Lett. 2021, 368, fnab138. [Google Scholar] [CrossRef]
- Alipour Kafi, S.; Karimi, E.; Akhlaghi Motlagh, M.; Amini, Z.; Mohammadi, A.; Sadeghi, A. Isolation and identification of Amycolatopsis sp. strain 1119 with potential to improve cucumber fruit yield and induce plant defense responses in commercial greenhouse. Plant Soil 2021, 468, 125–145. [Google Scholar] [CrossRef]
- Yu, Y.; Gui, Y.; Li, Z.; Jiang, C.; Guo, J.; Niu, D. Induced systemic resistance for improving plant immunity by beneficial microbes. Plants 2022, 11, 386. [Google Scholar] [CrossRef] [PubMed]
- Vilasinee, S.; Toanuna, C.; McGovern, R.J.; Nalumpang, S. Expression of pathogenesis-related (PR) genes in tomato against Fusarium wilt by challenge inoculation with Streptomyces NSP3. Int. J. Agric. Technol. 2019, 15, 157–170. [Google Scholar]
- Bizuneh, G.K. The chemical diversity and biological activities of phytoalexins. Tradit. Med. 2021, 21, 31–43. [Google Scholar] [CrossRef]
- Vijayabharathi, R.; Gopalakrishnan, S.; Sathya, A.; Vasanth Kumar, M.; Srinivas, V.; Mamta, S. Streptomyces sp. as plant growth-promoters and host-plant resistance inducers against Botrytis cinerea in chickpea. Biocontrol Sci. Technol. 2018, 28, 1140–1163. [Google Scholar] [CrossRef]
- Li, Y.; He, F.; Guo, Q.; Feng, Z.; Zhang, M.; Ji, C.; Xue, Q.; Lai, H. Compositional and functional comparison on the rhizosphere microbial community between healthy and Sclerotium rolfsii-infected monkshood (Aconitum carmichaelii) revealed the biocontrol potential of healthy monkshood rhizosphere microorganisms. Biol. Control 2022, 165, 104790. [Google Scholar] [CrossRef]
- Ameen, F.; AlNadhari, S.; Al-Homaidan, A.A. Marine microorganisms as an untapped source of bioactive compounds. Saudi J. Biol. Sci. 2021, 28, 224–231. [Google Scholar] [CrossRef]
- Vijayabharathi, R.; Gopalakrishnan, S.; Sathya, A.; Srinivas, V.; Sharma, M. Deciphering the tri-dimensional effect of endophytic Streptomyces sp. on chickpea for plant growth promotion, helper effect with Mesorhizobium ciceri and host-plant resistance induction against Botrytis cinerea. Microb. Pathog. 2018, 122, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Jing, T.; Zhou, D.; Zhang, M.; Yun, T.; Qi, D.; Wei, Y.; Chen, Y.; Zang, X.; Wang, W.; Xie, J. Newly isolated Streptomyces sp. JBS5-6 as a potential biocontrol agent to control banana Fusarium wilt: Genome sequencing and secondary metabolite cluster profiles. Front. Microbiol. 2020, 11, 3036. [Google Scholar] [CrossRef]
- Jepsen, T.; Jensen, B.; Jørgensen, N.O.G. Volatiles produced by Streptomyces spp. delay rot in apples caused by Colletotrichum acutatum. Curr. Res. Microb. Sci. 2022, 3, 100121. [Google Scholar] [CrossRef]
- Buatong, J.; Rukachaisirikul, V.; Sangkanu, S.; Surup, F.; Phongpaichit, S. Antifungal metabolites from marine-derived Streptomyces sp. AMA49 against Pyricularia oryzae. J. Pure Appl. Microbiol. 2019, 13, 653–665. [Google Scholar] [CrossRef] [Green Version]
- Uba, B.O.; Okoye, E.B.; Anyaeji, O.J.; Ogbonnaya, O.C. Antagonistic potentials of actinomycetes isolated from coastal area of Niger Delta against Citrus sinensis (Sweet Orange) and Lycopersicum esculentum (Tomato) fungal pathogens. Res. Rev. A J. Biotech. 2019, 9, 4–15. [Google Scholar]
- Torres-Rodriguez, J.; Reyes-Pérez, J.; Castellanos, T.; Angulo, C.; Quiñones-Aguilar, E.; Hernandez-Montiel, L. Identication and morphological characterization of marine actinomycetes as biocontrol agents of Fusarium solani in tomato. Rev. Fac. Agron. Univ. Zulia 2022, 39, e223915. [Google Scholar] [CrossRef]
- Wan, M.; Li, G.; Zhang, J.; Jiang, D.; Huang, H.C. Effect of volatile substances of Streptomyces platensis F-1 on control of plant fungal diseases. Biol. Control 2008, 46, 552–559. [Google Scholar] [CrossRef]
- Shimizu, M.; Yazawa, S.; Ushijima, Y. A promising strain of endophytic Streptomyces sp. for biological control of cucumber anthracnose. J. Gen. Plant Pathol. 2009, 75, 27–36. [Google Scholar] [CrossRef]
- Gholami, M.; Khakvar, R.; AliasgarZad, N. Application of endophytic bacteria for controlling anthracnose disease (Colletotrichum lindemuthianum) on bean plants. Arch. Phytopathol. Plant Prot. 2013, 46, 1831–1838. [Google Scholar] [CrossRef]
- Goudjal, Y.; Toumatia, O.; Yekkour, A.; Sabaou, N.; Mathieu, F.; Zitouni, A. Biocontrol of Rhizoctonia solani damping-off and promotion of tomato plant growth by endophytic actinomycetes isolated from native plants of Algerian Sahara. Microbiol. Res. 2014, 169, 59–65. [Google Scholar] [CrossRef]
- Malviya, N.; Yandigeri, M.S.; Yadav, A.K.; Solanki, M.K.; Arora, D.K. Isolation and characterization of novel alkali-halophilic actinomycetes from the Chilika brackish water lake, India. Ann. Microbiol. 2014, 64, 1829–1838. [Google Scholar] [CrossRef]
- Yandigeri, M.S.; Malviya, N.; Solanki, M.K.; Shrivastava, P.; Sivakumar, G. Chitinolytic Streptomyces vinaceusdrappus S5MW2 isolated from Chilika lake, India enhances plant growth and biocontrol efficacy through chitin supplementation against Rhizoctonia solani. World J. Microbiol. Biotechnol. 2015, 31, 1217–1225. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Ma, Z.; Xu, X.; Yu, X. Isolation and identification of biocontrol agent Streptomyces rimosus M527 against Fusarium oxysporum f. sp. cucumerinum. J. Basic Microbiol. 2016, 56, 929–933. [Google Scholar] [CrossRef]
- Mingma, R.; Duangmal, K. Characterization, antifungal activity and plant growth promoting potential of endophytic actinomycetes isolated from rice (Oryza sativa L.). Chiang Mai J. Sci. 2018, 45, 2652–2665. [Google Scholar]
- Adlin Jenifer, J.S.C.; Michaelbabu, M.; Eswaramoorthy Thirumalaikumar, C.L.; Jeraldin Nisha, S.R.; Uma, G.; Citarasu, T. Antimicrobial potential of haloalkaliphilic Nocardiopsis sp. AJ1 isolated from solar salterns in India. J. Basic Microbiol. 2019, 59, 288–301. [Google Scholar] [CrossRef]
- Hao, L.; Zheng, X.; Wang, Y.; Li, S.; Shang, C.; Xu, Y. Inhibition of tomato early blight disease by culture extracts of a Streptomyces puniceus isolate from mangrove soil. Phytopathology 2019, 109, 1149–1156. [Google Scholar] [CrossRef]
- Wonglom, P.; Suwannarach, N.; Lumyong, S.; Ito, S.; Matsui, K.; Sunpapao, A. Streptomyces angustmyceticus NR8-2 as a potential microorganism for the biological control of leaf spots of Brassica rapa subsp. pekinensis caused by Colletotrichum sp. and Curvularia lunata. Biol. Control 2019, 138, 104046. [Google Scholar] [CrossRef]
- Benhadj, M.; Metrouh, R.; Menasria, T.; Gacemi-Kirane, D.; Slim, F.Z.; Ranque, S. Broad-spectrum antimicrobial activity of wetland-derived Streptomyces sp. ActiF450. Excli J. 2020, 19, 360–371. [Google Scholar] [CrossRef]
- Cao, P.; Li, C.; Wang, H.; Yu, Z.; Xu, X.; Wang, X.; Zhao, J.; Xiang, W. Community structures and antifungal activity of root-associated endophytic actinobacteria in healthy and diseased cucumber plants and Streptomyces sp. HAAG3-15 as a promising biocontrol agent. Microorganisms 2020, 8, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghadhban, W.I.; Abd Burghal, A. Identification of antibiotic producing actinomycetes isolated from sediment in Basra, Iraq. Int. J. Pharm. Sci. 2020, 11, b92–b98. [Google Scholar] [CrossRef]
- Peng, C.; An, D.; Ding, W.-X.; Zhu, Y.-X.; Ye, L.; Li, J. Fungichromin production by Streptomyces sp. WP-1, an endophyte from Pinus dabeshanensis, and its antifungal activity against Fusarium oxysporum. Appl. Microbiol. Biotechnol. 2020, 104, 10437–10449. [Google Scholar] [CrossRef]
- Abo-Zaid, G.; Abdelkhalek, A.; Matar, S.; Darwish, M.; Abdel-Gayed, M. Application of bio-friendly formulations of chitinase-producing Streptomyces cellulosae Actino 48 for controlling peanut soil-borne diseases caused by Sclerotium rolfsii. J. Fungi 2021, 7, 167. [Google Scholar] [CrossRef]
- Boukaew, S.; Cheirsilp, B.; Prasertsan, P.; Yossan, S. Antifungal effect of volatile organic compounds produced by Streptomyces salmonis PSRDC-09 against anthracnose pathogen Colletotrichum gloeosporioides PSU-03 in postharvest chili fruit. J. Appl. Microbiol. 2021, 131, 1452–1463. [Google Scholar] [CrossRef]
- Li, X.; Li, K.; Zhou, D.; Zhang, M.; Qi, D.; Jing, T.; Zang, X.; Qi, C.; Wang, W.; Xie, J. Biological control of banana wilt disease caused by Fusarium oxyspoum f. sp. Cubense using Streptomyces sp. H4. Biol. Control 2021, 155, 104524. [Google Scholar] [CrossRef]
- Ziedan, E.S.H. A review of the efficacy of biofumigation agents in the control of soil-borne plant diseases. J. Plant Prot. Res. 2022, 62, 1–11. [Google Scholar] [CrossRef]
- El-Tarabily, K.A.; Nassar, A.H.; Hardy, G.S.J.; Sivasithamparam, K. Plant growth promotion and biological control of Pythium aphanidermatum, a pathogen of cucumber, by endophytic actinomycetes. J. Appl. Microbiol. 2009, 107, 1765–1766. [Google Scholar] [CrossRef]
- Costa, F.G.; Zucchi, T.D.; Melo, I.S.D. Biological control of phytopathogenic fungi by endophytic actinomycetes isolated from maize (Zea mays L.). Braz. Arch. Biol. Technol. 2013, 56, 948–955. [Google Scholar] [CrossRef]
- Singh, S.P.; Gupta, R.; Gaur, R.; Srivastava, A.K. Antagonistic actinomycetes mediated resistance in Solanum lycopersicon Mill. against Rhizoctonia solani Kühn. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2017, 87, 789–798. [Google Scholar] [CrossRef]
- Anusha, B.G.; Gopalakrishnan, S.; Naik, M.K.; Sharma, M. Evaluation of Streptomyces spp. and Bacillus spp. for biocontrol of Fusarium wilt in chickpea (Cicer arietinum L.). Arch. Phytopathol. Plant Prot. 2019, 52, 417–442. [Google Scholar] [CrossRef]
- Barnett, S.J.; Ballard, R.A.; Franco, C.M. Field assessment of microbial inoculants to control Rhizoctonia root rot on wheat. Biol. Control 2019, 132, 152–160. [Google Scholar] [CrossRef] [Green Version]
- Pathom-aree, W.; Kreawsa, S.; Kamjam, M.; Tokuyama, S.; Yoosathaporn, S.; Lumyong, S. Potential of selected mangrove Streptomyces as plant growth promoter and rice bakanae disease control agent. Chiang Mai J. Sci. 2019, 46, 261–276. [Google Scholar]
- Zhang, J.; Gao, Z.; Li, N.; Zhang, X. The antifungal activity of biocontrol actinomycetes G-1 against Alternaria radicina. J. Henan Agric. Sci. 2019, 48, 99–104. [Google Scholar]
- El-Shatoury, S.A.; Ameen, F.; Moussa, H.; Wahid, O.A.; Dewedar, A.; AlNadhari, S. Biocontrol of chocolate spot disease (Botrytis cinerea) in faba bean using endophytic actinomycetes Streptomyces: A field study to compare application techniques. PeerJ 2020, 8, e8582. [Google Scholar] [CrossRef] [Green Version]
- Gu, L.; Zhang, K.; Zhang, N.; Li, X.; Liu, Z. Control of the rubber anthracnose fungus Colletotrichum gloeosporioides using culture filtrate extract from Streptomyces deccanensis QY-3. Antonie Van Leeuwenhoek 2020, 113, 1573–1585. [Google Scholar] [CrossRef]
- Saeed, I.; Khan, S.H.; Rasheed, A.; Jahangir, M.M.; Jabbar, A.; Shaheen, H.M.F.; Din, W.U.; Mazhar, K. Assessment of antagonistic potential of bacteria as biocontrol agent against Alternaria leaf spot of turnip. Pak. J. Phytopathol. 2021, 33, 401–409. [Google Scholar] [CrossRef]
- Yao, X.; Zhang, Z.; Huang, J.; Wei, S.; Sun, X.; Chen, Y.; Liu, H.; Li, S. Candicidin isomer production is essential for biocontrol of cucumber Rhizoctonia rot by Streptomyces albidoflavus W68. Appl. Environ. Microbiol. 2021, 87, e03078-20. [Google Scholar] [CrossRef]
- Shewa, A.G.; Gobena, D.A.; Ali, M.K. Review on postharvest quality and handling of apple. J. Agric. Sci. Food Technol. 2022, 8, 028–032. [Google Scholar] [CrossRef]
- Li, Y.; Xia, M.; He, P.; Yang, Q.; Wu, Y.; He, P.; Ahmed, A.; Li, X.; Wang, Y.; Munir, S.; et al. Developing Penicillium digitatum management strategies on post-harvest citrus fruits with metabolic components and colonization of Bacillus subtilis L1-21. J. Fungi 2022, 8, 80. [Google Scholar] [CrossRef]
- Sarkar, G.; Suthindhiran, K. Diversity and biotechnological potential of marine actinomycetes from India. Indian J. Microbiol. 2022, Vol., 1–19. [Google Scholar] [CrossRef]
- Hu, X.; Cheng, B.; Du, D.; Huang, Z.; Pu, Z.; Chen, G.; Peng, A.; Lu, L. Isolation and identification of a marine actinomycete strain and its control efficacy against citrus green and blue moulds. Biotechnol. Biotechnol. Equip. 2019, 33, 719–729. [Google Scholar] [CrossRef]
- Najmeh, S.; Hosein, S.B.G.; Sareh, S.; Bonjar, L.S. Biological control of citrus green mould, Penicillium digitatum, by antifungal activities of Streptomyces isolates from agricultural soils. Afr. J. Microbiol. Res. 2014, 8, 1501–1509. [Google Scholar] [CrossRef] [Green Version]
- Yong, D.; Wang, C.; Li, G.; Li, B. Control efficiency of endophytic actinomycetes A-1 against apple fruit ring rot and its influence on the activity of defense-related enzymes. Acta Phytophylacica Sin. 2014, 41, 335–341. [Google Scholar]
- Lyu, A.; Liu, H.; Che, H.; Yang, L.; Zhang, J.; Wu, M.; Chen, W.; Li, G. Reveromycins A and B from Streptomyces sp. 3-10: Antifungal activity against plant pathogenic fungi in vitro and in a strawberry food model system. Front. Microbiol. 2017, 8, 3–10. [Google Scholar] [CrossRef]
- Boukaew, S.; Petlamul, W.; Bunkrongcheap, R.; Chookaew, T.; Kabbua, T.; Thippated, A.; Prasertsan, P. Fumigant activity of volatile compounds of Streptomyces philanthi RM-1-138 and pure chemicals (acetophenone and phenylethyl alcohol) against anthracnose pathogen in postharvest chili fruit. Crop Prot. 2018, 103, 1–8. [Google Scholar] [CrossRef]
- Liotti, R.G.; da Silva Figueiredo, M.I.; Soares, M.A. Streptomyces griseocarneus R132 controls phytopathogens and promotes growth of pepper (Capsicum annuum). Biol. Control 2019, 138, 104065. [Google Scholar] [CrossRef]
- Boukaew, S.; Petlamul, W.; Prasertsan, P. Comparison of the biocontrol efficacy of culture filtrate from Streptomyces philanthi RL-1-178 and acetic acid against Penicillium digitatum, in vitro and in vivo. Eur. J. Plant Pathol. 2020, 158, 939–949. [Google Scholar] [CrossRef]
- Kim, J.D.; Kang, J.E.; Kim, B.S. Postharvest disease control efficacy of the polyene macrolide lucensomycin produced by Streptomyces plumbeus strain CA5 against gray mold on grapes. Postharvest Biol. Technol. 2020, 162, 111115. [Google Scholar] [CrossRef]
- Nguyen, H.T.T.; Park, A.R.; Hwang, I.M.; Kim, J.C. Identification and delineation of action mechanism of antifungal agents: Reveromycin E and its new derivative isolated from Streptomyces sp. JCK-6141. Postharvest Biol. Technol. 2021, 182, 111700. [Google Scholar] [CrossRef]
- Chaurasia, A.; Meena, B.R.; Tripathi, A.N.; Pandey, K.K.; Rai, A.B.; Singh, B. Actinomycetes: An unexplored microorganisms for plant growth promotion and biocontrol in vegetable crops. World J. Microbiol. Biotechnol. 2018, 34, 132. [Google Scholar] [CrossRef]
- Lahdenperä, M.L.; Simon, E.; Uoti, J. Mycostop-a novel biofungicide based on Streptomyces bacteria. In Developments in Agricultural and Managed Forest Ecology; Beemster, A.B.R., Bollen, G.J., Gerlagh, M., Ruissen, M.A., Schippers, B., Tempel, A., Eds.; Elsevier: Amsterdam, The Netherlands, 1991; Volume 23, pp. 258–263. [Google Scholar]
- Tian, X.; Zheng, Y. Evaluation of biological control agents for Fusarium wilt in Hiemalis begonia. Can. J. Plant Pathol. 2013, 35, 363–370. [Google Scholar] [CrossRef]
- Kabaluk, J.T.; Svircev, A.M.; Goettel, M.S.; Woo, S.G. The Use and Regulation of Microbial Pesticides in Representative Jurisdiction Worldwide; IOBC Global: Hong Kong, China, 2010; p. 99. [Google Scholar]
- Okuyama, A.; Machiyama, N.; Kinoshita, T.; Tanaka, N. Inhibition by kasugamycin of initiation complex formation on 30S ribosomes. Biochem. Biophys. Res. Commun. 1971, 43, 196–199. [Google Scholar] [CrossRef] [PubMed]
- Copping, L.G.; Duke, S.O. Natural products that have been used commercially as crop protection agents. Pest Manag. Sci. 2007, 63, 524–554. [Google Scholar] [CrossRef] [PubMed]
- Copping, L.G.; Menn, J.J. Biopesticides: A review of their action, applications and efficacy. Pest Manag. Sci. 2000, 56, 651–676. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Actinomycete | Phytopathogen | Host | In Vivo Inhibition | Antagonistic Mechanisms | Reference |
|---|---|---|---|---|---|
| Streptomyces sp. | Colletotrichum fragariae | Strawberry | 100% | Secondary metabolites | [19] |
| S. sampsonii | Sclerotinia sclerotiorum | Green bean | 100% | Secondary metabolites | [11] |
| Streptomyces sp. | Ralstonia solanacearum | Tomato | 97% | Induction of host resistance | [13] |
| S. sichuanensis | Fusarium oxysporum | Banana | 51% | Siderophores | [25] |
| Amycolatopsis sp. | F. graminearum | Maize | 79% | Lytic enzyme | [26] |
| Arthrobacter humicola | A. alternata | Tomato | 31% | Secondary metabolites | [27] |
| Nocardiopsis dassonvillei | Bipolaris sorokiniana | Wheat | 72% | Siderophores and lytic enzyme | [28] |
| S. rameus | R. bataticola | Bean | 70% | Siderophores and lytic enzyme | [10] |
| S. globisporous | R. solani | Tomato | 50% | Induction of host resistance | [29] |
| Commercial Product | Actinomycete | Registered Countries | Phytopathogen Species/Target Disease | Main Effects | Reference |
|---|---|---|---|---|---|
| Mycostop | S. griseovirids | Canada, UE countries, and USA | Alternaria, R. solani, Fusarium, Botrytis, Phytophthora, and Pythium | Space and nutrient competence and produces polyenic antibiotics | [111] |
| Actinovate | S. lydicus | Canada and USA | Pythium, Fusarium, Phytophthora, Rhizoctonia, and Verticillium, powdery and downy mildew, and Botrytis, Alternaria, Geotrichum, and Sclerotinia | Induces resistance in plants and produces extracellular chitinases | [112] |
| Mycocide KIBC | S. colombiensis | South Korea | Powdery mildews, grey mold, and brown patch | Produces enzymes and antibiotics | [113] |
| Safegrow KIBC | S. kasugaensis | South Korea | Sheath blight and large patch | Produces enzymes and antibiotics | [113] |
| Kasugamycin, Kasumin | S. kasugaensis | Ukraine | Leaf spot, scab, and root rot | Inhibit protein biosynthesis | [114] |
| Agrimycin, Paushak, Cuprimicin 17, Astrepto 17 | S. griseus | India, USA, New Zealand, China, Ukraine and Canada | Bacterial rots, Xanthomona, and Pseudomonas | Inhibit protein biosynthesis | [112] |
| Polyoxorim (Endorse, Polyoxin Z and Stopit) | S. cacaoi var. asoensis | UE countries | Sphaerotheca, powdery mildews, Botrytis, Sclerotium, Corynespora, Cochliobolus, Alternaria, sheath blight, and Helminthosporium | Inhibit cell wall biosynthesis and causes abnormal spore germ tube swelling and hypha points | [115] |
| Validacin, Valimun, Dantotsupadanvalida, Mycin Hustler, Valida | S. hygroscopicus | - | Rhizoctonia | Inhibit trehalase in Rhizoctonia | [116] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torres-Rodriguez, J.A.; Reyes-Pérez, J.J.; Quiñones-Aguilar, E.E.; Hernandez-Montiel, L.G. Actinomycete Potential as Biocontrol Agent of Phytopathogenic Fungi: Mechanisms, Source, and Applications. Plants 2022, 11, 3201. https://doi.org/10.3390/plants11233201
Torres-Rodriguez JA, Reyes-Pérez JJ, Quiñones-Aguilar EE, Hernandez-Montiel LG. Actinomycete Potential as Biocontrol Agent of Phytopathogenic Fungi: Mechanisms, Source, and Applications. Plants. 2022; 11(23):3201. https://doi.org/10.3390/plants11233201
Chicago/Turabian StyleTorres-Rodriguez, Juan A., Juan J. Reyes-Pérez, Evangelina E. Quiñones-Aguilar, and Luis G. Hernandez-Montiel. 2022. "Actinomycete Potential as Biocontrol Agent of Phytopathogenic Fungi: Mechanisms, Source, and Applications" Plants 11, no. 23: 3201. https://doi.org/10.3390/plants11233201
APA StyleTorres-Rodriguez, J. A., Reyes-Pérez, J. J., Quiñones-Aguilar, E. E., & Hernandez-Montiel, L. G. (2022). Actinomycete Potential as Biocontrol Agent of Phytopathogenic Fungi: Mechanisms, Source, and Applications. Plants, 11(23), 3201. https://doi.org/10.3390/plants11233201