
Real-Time Collection of Conidia Released from Living Single Colonies of Podosphaera aphanis on Strawberry Leaves under Natural Conditions with Electrostatic Techniques

 and
and
Abstract
:1. Introduction
2. Results
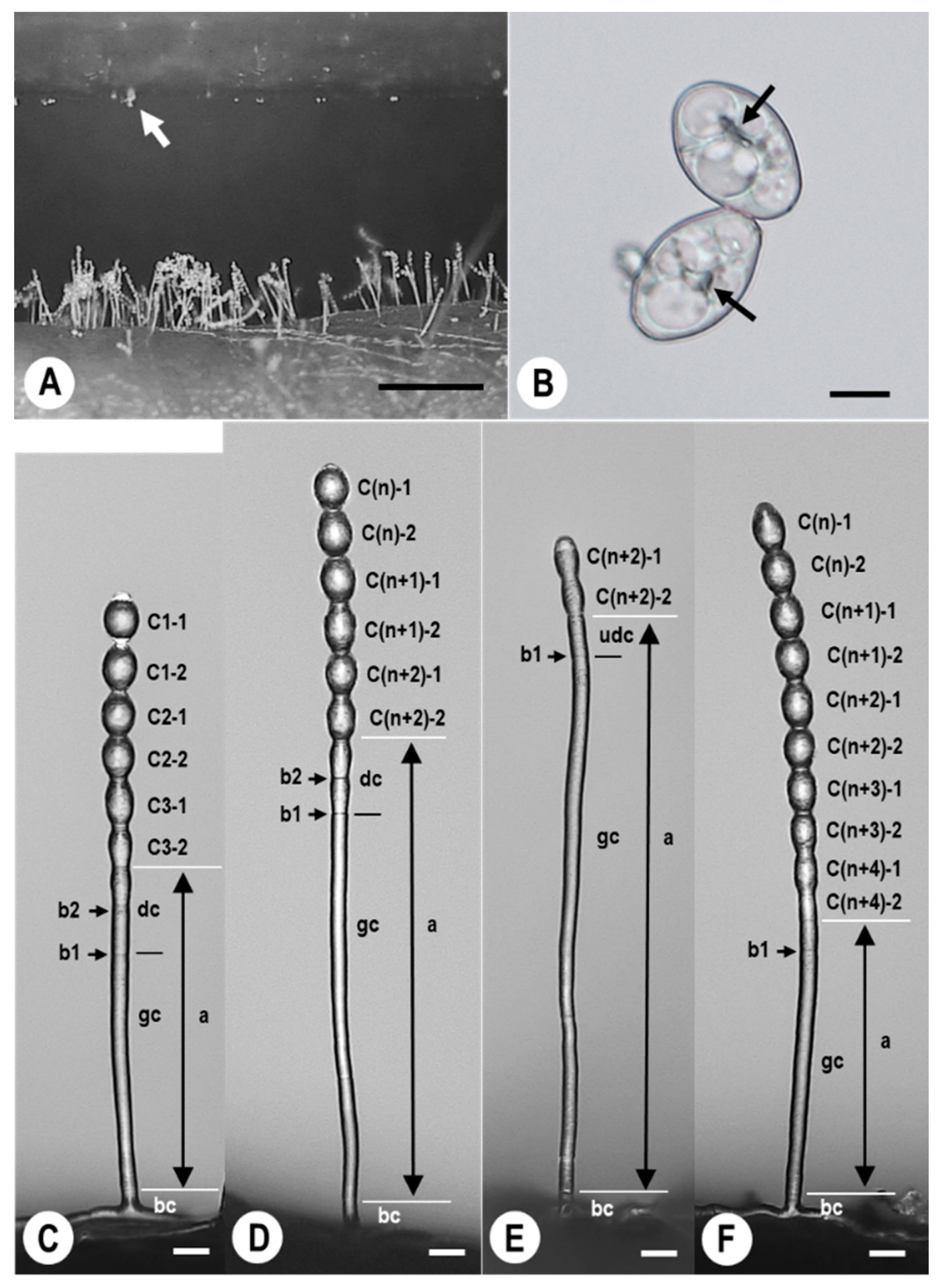
2.1. Morphological Observation of KSP-7N Conidia and Conidiophores Developed under Static Electricity
2.2. Germination Rates of KSP-7N Conidia Electrostatically Collected on Insulator Films
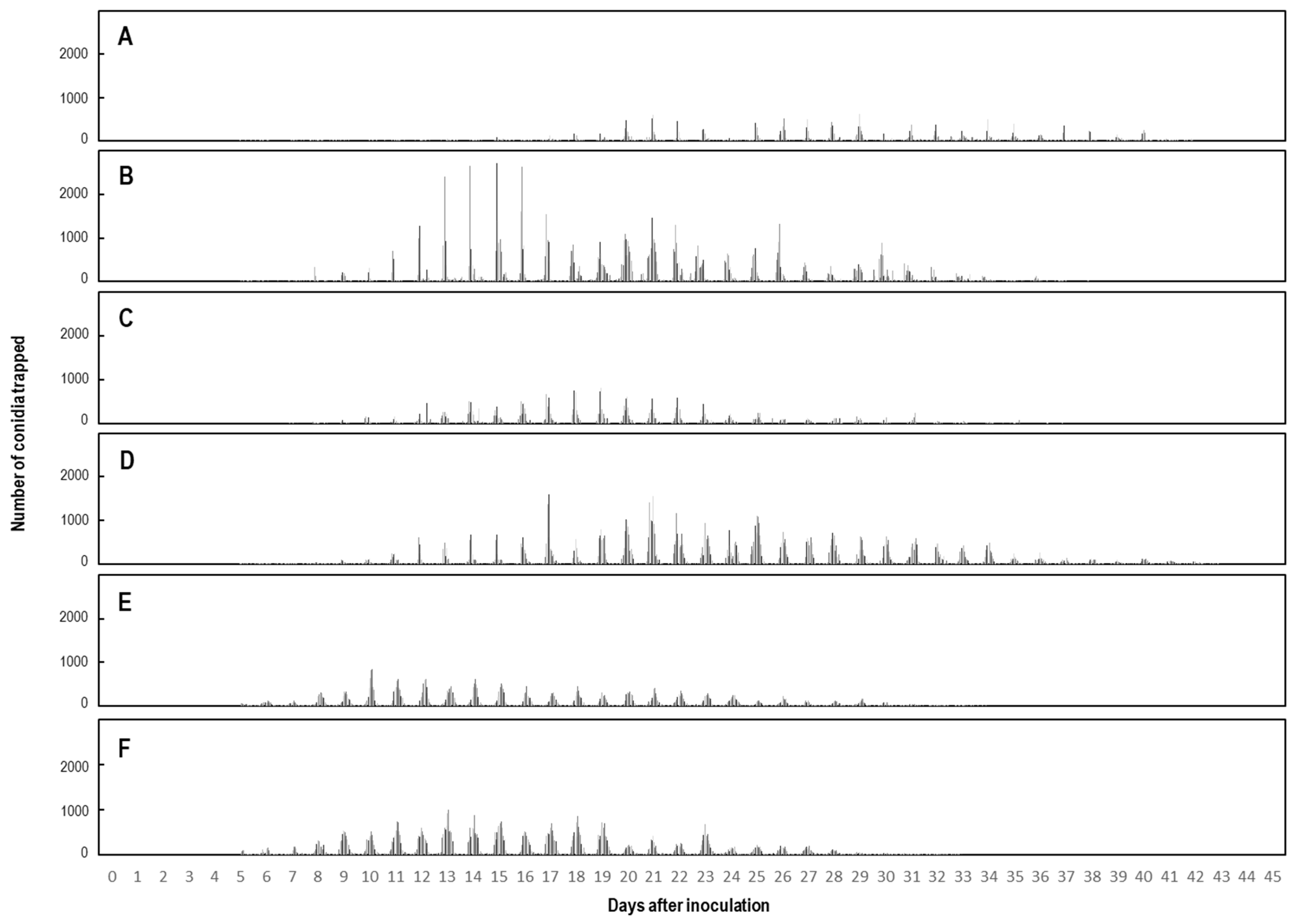
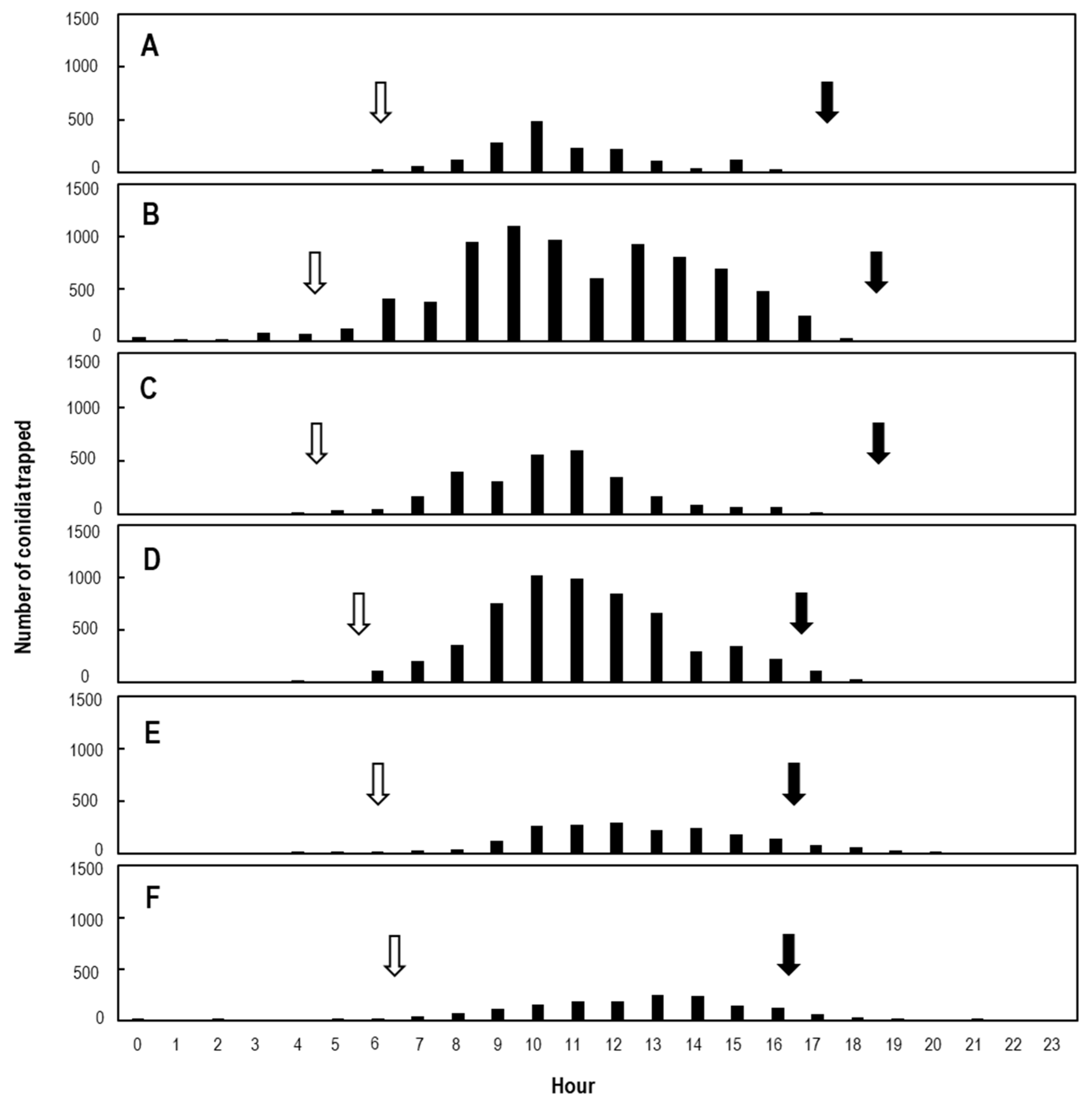
2.3. Total Number of Progeny Conidia Electrostatically Collected from Single KSP-7N Colonies under Greenhouse Conditions
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Cultivation
4.2. Fungal Materials, Inoculation and Incubation
4.3. Electrostatic Spore Collector
4.4. Electrostatically Activated Insulator Film
4.5. Continuous Collection of Progeny Conidia Dispersed from Single KSP-7N Colonies
4.6. Germination Rates of Conidia Collected with an Electrostatic Spore Collector
4.7. Observation of Single KSP-7N Colonies with Digital and Light Microscopes
4.8. Statistical Analysis with Tukey’s Test
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Braun, U.; Takamatsu, S. Phylogeny of Erysiphe, Microsphaera, Uncinula (Erysipheae) and Cystotheca, Podosphaera, Sphaerotheca (Cystotheceae) inferred from rDNA ITS sequences–some taxonomic consequences. Schlechtendalia 2000, 4, 1–33. [Google Scholar]
- Braun, U.; Cook, R.T.A. Taxonomic Manual of the Erysiphales (Powdery Mildews); CBS-KNAW Fungal Biodiversity Centre: Utrecht, The Netherlands, 2012. [Google Scholar]
- Peries, O.S. Studies on strawberry mildew, caused by Sphaerotheca macularis (Wallr. ex Fries) Jaczewski. I. Biology of the fungus. Ann. Appl. Biol. 1962, 50, 211–224. [Google Scholar] [CrossRef]
- Peries, O.S. Studies on strawberry mildew, caused by Sphaerotheca macularis (Wallr. ex Fries) Jaczewski. II. Host-parasite relationships on the foliage of strawberry varieties. Ann. Appl. Biol. 1962, 50, 225–233. [Google Scholar] [CrossRef]
- Jhooty, J.S.; McKeen, W.E. Studies on powdery mildew of strawberry caused by Sphaerotheca macularis. Phytopathology 1965, 55, 281–285. [Google Scholar]
- Spencer, D.M. Powdery mildew of strawberries. In The Powdery Mildews; Spencer, D.M., Ed.; Academic Press: New York, NY, USA, 1978; pp. 355–358. [Google Scholar]
- Maas, J.L. Compendium of Strawberry Diseases, 2nd ed.; APS Press: St. Paul, MN, USA, 1998. [Google Scholar]
- Nelson, M.D.; Gubler, W.D.; Shaw, D.V. Inheritance of powdery mildew resistance in greenhouse-grown versus field-grown California strawberry progenies. Phytopathology 1995, 85, 421–424. [Google Scholar] [CrossRef]
- Xiao, C.L.; Chandler, C.K.; Price, J.F.; Duval, J.R.; Mertely, J.C.; Legard, D.E. Comparison of epidemics of Botrytis fruit rot and powdery mildew of strawberry in large plastic tunnel and field production systems. Plant Dis. 2001, 85, 901–909. [Google Scholar] [CrossRef] [Green Version]
- Carisse, O.; Morissette-Thomas, V.; van der Heyden, H. Lagged association between powdery mildew leaf severity, airborne inoculum, weather, and crop losses in strawberry. Phytopathology 2013, 103, 811–821. [Google Scholar] [CrossRef] [Green Version]
- Tanigawa, M.; Nakano, T.; Hagihara, T.; Okayama, K.; Sezaki, S. Relationship between the control effect of fungicides on powdery mildew (Sphaerotheca humuli) and their deposits on strawberry leaves. J. Pesticide Sci. 1993, 18, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Okayama, K.; Nakano, T.; Matsutani, S.; Sugimura, T. A simple and reliable method for evaluating the effectiveness of fungicides for control of powdery mildew (Sphaerotheca macularis) on strawberry. Ann. Phytopathol. Soc. Jpn. 1995, 61, 536–540. [Google Scholar] [CrossRef]
- Nakazawa, Y.; Uchida, K. First record of cleistothecial stage of powdery mildew fungus on strawberry in Japan. Ann. Phytopathol. Soc. Jpn. 1998, 64, 121–124. [Google Scholar] [CrossRef]
- Iwasaki, S.; Asano, S.; Yoshida, K.; Kitamura, S.; Taira, A.; Honda, Y.; Suzuki, T.; Takikawa, Y.; Kakutani, K.; Matsuda, Y.; et al. Analysis of conidiogenesis and lifelong conidial production from single conidiophores of Podosphaera aphanis on strawberry leaves using digital microscopic and electrostatic techniques. Australas. Plant Pathol. 2021, 50, 571–587. [Google Scholar] [CrossRef]
- Braun, U. A monograph of the Erysiphales (powdery mildews). Beih. Nova Hedwig. 1987, 89, 1–700. [Google Scholar]
- Takamatsu, S.; Niinomi, S.; Harada, M.; Havrylenko, M. Molecular phylogenetic analyses reveal a close evolutionary relationship between Podosphaera (Erysiphales: Erysiphaceae) and its rosaceous hosts. Persoonia 2010, 24, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Kanto, T.; Matsuura, K.; Ogawa, T.; Yamada, M.; Ishiwata, M.; Usami, T.; Amemiya, Y. A new UV-B lighting system controls powdery mildew of strawberry. Acta Hortic. 2014, 1049, 655–660. [Google Scholar] [CrossRef]
- Janisiewicz, W.J.; Takeda, F.; Jurick, W.; Nichols, B.; Wolford, S.; Glenn, D.M. A novel approach to control gray mold, anthracnose, and powdery mildew on strawberry using low-dose UV-C irradiation. Phytopathology 2015, 105, S4.64. [Google Scholar]
- Freeman, J.A.; Pepin, H.S. A systemic fungicide (fungicide 1991) for the control of gray mold and powdery mildew in strawberries and raspberries. Can. Plant Dis. Surv. 1967, 47, 104–107. [Google Scholar]
- Hukkanen, A.T.; Kokko, H.I.; Buchala, A.J.; McDougall, G.J.; Stewart, D.; Kärenlampi, S.O.; Karjalainen, R.O. Benzothiadiazole induces the accumulation of phenolics and improves resistance to powdery mildew in strawberries. J. Agric. Food Chem. 2007, 55, 1862–1870. [Google Scholar] [CrossRef] [Green Version]
- Miller, T.C.; Gubler, W.D.; Laemmlen, F.F.; Geng, S.; Rizzo, D.M. Potential for using Lecanicillium lecanii for suppression of strawberry powdery mildew. Biocont. Sci. Technol. 2004, 14, 215–220. [Google Scholar] [CrossRef]
- Davik, J.; Honne, B.I. Genetic variance and breeding values for resistance to a wind-borne disease [Sphaerotheca macularis (Wallr. ex Fr.)] in strawberry (Fragaria × ananassa Duch.) estimated by exploring mixed and spatial models and pedigree information. Theor. Appl. Genet. 2005, 111, 256–264. [Google Scholar] [CrossRef]
- De Cal, A.; Redondo, C.; Sztejnberg, A.; Melgarejo, P. Biocontrol of powdery mildew by Penicillium oxalicum in open-field nurseries of strawberries. Biol. Control. 2008, 47, 103–107. [Google Scholar] [CrossRef]
- Pertot, I.; Zasso, R.; Amsalem, L.; Baldessari, M.; Angeli, G.; Elad, Y. Integrating biocontrol agents in strawberry powdery mildew control strategies in high tunnel growing systems. Crop Prot. 2008, 27, 622–631. [Google Scholar] [CrossRef]
- Cockerton, H.M.; Vickerstaff, R.J.; Karlström, A.; Wilson, F.; Sobczyk, M.; He, J.Q.; Sargent, D.J.; Passey, A.J.; McLeary, K.J.; Pakozdi, K.; et al. Identification of powdery mildew resistance QTL in strawberry (Fragaria × ananassa). Theor. Appl. Genet. 2018, 131, 1995–2007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aylor, D.E. The role of intermittent wind in the dispersal of fungal pathogens. Annu. Rev. Phytopathol. 1990, 28, 73–92. [Google Scholar] [CrossRef]
- Brown, J.K.M.; Hovmøller, M.S. Aerial dispersal of pathogens on the global and continental scales and its impact on plant disease. Science 2002, 297, 537–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nonomura, T.; Matsuda, Y.; Yamashita, S.; Akahoshi, H.; Takikawa, Y.; Kakutani, K.; Toyoda, H. Natural woody plant, Mallotus japonicus, as an ecological partner to transfer different pathotypic conidia of Oidium neolycopersici to greenhouse tomatoes. Plant Protect. Sci. 2013, 49, S33–S40. [Google Scholar] [CrossRef]
- Oichi, W.; Matsuda, Y.; Sameshima, T.; Nonomura, T.; Kakutani, K.; Nishimura, H.; Kusakari, S.; Toyoda, H. Consecutive monitoring for conidiogenesis by Oidium neolycopersici on tomato leaves with a high-fidelity digital microscope. J. Gen. Plant Pathol. 2004, 70, 329–332. [Google Scholar] [CrossRef]
- Oichi, W.; Matsuda, Y.; Nonomura, T.; Toyoda, H.; Xu, L.; Kusakari, S. Formation of conidial pseudochains by tomato powdery mildew Oidium neolycopersici. Plant Dis. 2006, 90, 915–919. [Google Scholar] [CrossRef] [Green Version]
- Moriura, N.; Matsuda, Y.; Oichi, W.; Nakashima, S.; Hirai, T.; Sameshima, T.; Nonomura, T.; Kakutani, K.; Kusakari, S.; Higashi, K.; et al. Consecutive monitoring of lifelong production of conidia by individual conidiophores of Blumeria graminis f. sp. hordei on barley leaves by digital microscopic techniques with electrostatic micromanipulation. Mycol. Res. 2006, 110, 18–27. [Google Scholar] [CrossRef]
- Takikawa, Y.; Nonomura, T.; Miyamoto, S.; Okamoto, N.; Murakami, T.; Matsuda, Y.; Kakutani, K.; Kusakari, S.; Toyoda, H. Digital microscopic analysis of conidiogenesis of powdery mildew pathogens isolated from melon leaves. Phytoparasitica 2015, 43, 517–530. [Google Scholar] [CrossRef]
- Matsuda, Y.; Ikeda, H.; Moriura, N.; Tanaka, N.; Shimizu, K.; Oichi, W.; Nonomura, T.; Kakutani, K.; Kusakari, S.; Higashi, K.; et al. A new spore precipitator with polarized dielectric insulators for physical control of tomato powdery mildew. Phytopathology 2006, 96, 967–974. [Google Scholar] [CrossRef] [Green Version]
- Moriura, N.; Matsuda, Y.; Oichi, W.; Nakashima, S.; Hirai, T.; Nonomura, T.; Kakutani, K.; Kusakari, S.; Higashi, K.; Toyoda, H. An apparatus for collecting total conidia of Blumeria graminis f. sp. hordei from leaf colonies using electrostatic attraction. Plant Pathol. 2006, 55, 367–374. [Google Scholar] [CrossRef]
- Nonomura, T.; Matsuda, Y.; Xu, L.; Kakutani, K.; Takikawa, Y.; Toyoda, H. Collection of highly germinative pseudochain conidia of Oidium neolycopersici from conidiophores by electrostatic attraction. Mycol. Res. 2009, 113, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Nakamura, R.; Takagi, N.; Takikawa, Y.; Kakutani, K.; Matsuda, Y.; Matsui, K.; Nonomura, T. Quantitative analysis of the lifelong production of conidia released from single colonies of Podosphaera xanthii on melon leaves using electrostatic techniques. Australas. Plant Pathol. 2019, 48, 297–307. [Google Scholar] [CrossRef]
- Salmon, E.S. The strawberry mildew (Sphaerotheca humuli (DC.) Burr.). J. R. Hortic. Soc. 1900, 25, 132–138. [Google Scholar]
- Karajeh, M.R.; Al-Rawashdeh, Z.B.; Al-Dein, E.; Al-Ramamneh, M. Occurrence and control of strawberry powdery mildew in Al-Shoubak/Jordan. Jordan J. Agric. Sci. 2012, 8, 380–390. [Google Scholar]
- Calis, O.; Cekic, C.; Soylu, S.; Tor, M. Identification of new resistance sources from diploid wild strawberry against powdery mildew pathogen. Pak. J. Agri. Sci. 2015, 52, 677–683. [Google Scholar]
- de los Santos, B.; Blanco, C.; Porras, M.; Barrau, C.; Romero, F. First confirmation of Sphaerotheca macularis on strawberry plants in southwesthern Spain. Plant Dis. 2002, 86, 1049. [Google Scholar] [CrossRef]
- Paulus, A.O. Fungal diseases of strawberry. Hort. Sci. 1990, 25, 885–889. [Google Scholar] [CrossRef]
- Jhooty, J.S.; McKeen, W.E. The perfect stage of Sphaerotheca macularis on strawberry plants. Plant Dis. Reptr. 1962, 46, 218–219. [Google Scholar]
- Howard, C.M.; Albregts, E.E. Cleistothecia of Sphaerotheca macularis on strawberry plants in Florida. Plant Dis. 1982, 66, 261–262. [Google Scholar] [CrossRef]
- Farooq, M.; Dodgson, J.; Hall, A.M. Examination of the morphology of Podosphaera aphanis cleistothecia and their role in over wintering of the fungus. Asp. Appl. Biol. 2007, 83, 55–58. [Google Scholar]
- Abiko, K. Specialization of parasitism of Sphaerotheca humuli (DC.) Burr. Bull. Veg. Ornam. Crops Res. Stn. Jpn. Ser. A 1982, 10, 69–74, (In Japanese with English summary). [Google Scholar]
- Harvey, N.; Xu, X.-M. Powdery mildew on raspberry is genetically different from strawberry powdery mildew. J. Plant Pathol. 2010, 92, 775–779. [Google Scholar]
- Nelson, M.D.; Gubler, W.D.; Shaw, D.V. Relative resistance of 47 strawberry cultivars to powdery mildew in California greenhouse and field environments. Plant Dis. 1996, 80, 326–328. [Google Scholar] [CrossRef]
- Suzuki, T.; Iwasaki, S.; Hisazumi, H.; Miyamoto, A.; Ogami, H.; Takikawa, Y.; Kakutani, K.; Matsuda, Y.; Nonomura, T. Inhibitory effects of blue light-emitting diode irradiation on Podosphaera xanthii conidial release and infection of melon seedlings. Agriculture 2022, 12, 198. [Google Scholar] [CrossRef]
- Willocquet, L.; Sombardier, A.; Blancard, D.; Jolivet, J.; Savary, S. Spore dispersal and disease gradients in strawberry powdery mildew. Can. J. Plant Pathol. 2008, 30, 434–441. [Google Scholar] [CrossRef]
- Leach, C.M. An electrostatic theory to explain violent spore liberation by Drechslera turcica and other fungi. Mycologia 1976, 68, 63–86. [Google Scholar] [CrossRef] [PubMed]
- Griffith, W.T. Electrostatic phenomena. In The Physics of Everyday Phenomena: A Conceptual Introduction to Physics; Bruflodt, D., Loehr, B.S., Eds.; McGraw-Hill: New York, NY, USA, 2004; pp. 232–252. [Google Scholar]
- Halliday, D.; Resnick, R.; Walker, J. Electric charge. In Fundamentals of Physics; Johnson, S., Ford, E., Eds.; John Wiley & Sons: New York, NY, USA, 2005; pp. 561–579. [Google Scholar]
- Shimizu, T.; Matsuda, Y.; Nonomura, T.; Ikeda, H.; Tamura, N.; Kusakari, S.; Kimbara, J.; Toyoda, H. Dual protection of hydroponic tomatoes from rhizosphere pathogens Ralstonia solanacearum and Fusarium oxysporum f. sp. radicis-lycopersici and airborne conidia of Oidium neolycopersici with an ozone-generative electrostatic spore precipitator. Plant Pathol. 2007, 56, 987–997. [Google Scholar] [CrossRef]
- Suzuki, T.; Nishimura, S.; Yagi, K.; Nakamura, R.; Takikawa, Y.; Matsuda, Y.; Kakutani, K.; Nonomura, T. Effects of light quality on conidiophore formation of the melon powdery mildew pathogen Podosphaera xanthii. Phytoparasitica 2018, 94, 1105–1110. [Google Scholar] [CrossRef]
- Hirata, K. Notes on haustoria, hyphae and conidia of the powdery mildew fungus of barley, Erysiphe graminis f. sp. hordei. Mem. Fac. Agric. Nigata Univ. 1967, 6, 205–259. [Google Scholar]
- Takikawa, Y.; Kakutani, K.; Nonomura, T.; Matsuda, Y.; Toyoda, H. Conidia of Erysiphe trifoliorum attempt penetration twice during a two-step germination process on non-host barley leaves and an artificial hydrophobic surface. Mycoscience 2011, 52, 204–209. [Google Scholar] [CrossRef]
- Sameshima, T.; Kashimoto, K.; Kida, K.; Matsuda, Y.; Nonomura, T.; Kakutani, K.; Nakata, K.; Kusakari, S.; Toyoda, H. Cytological events in tomato leaves inoculated with conidia of Blumeria graminis f. sp. hordei and Oidium neolycopersici KTP-01. J. Gen. Plant Pathol. 2004, 70, 7–10. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Colony * | Colony Area (cm2) | Number of Conidiophores in a Single Colony | Duration of Conidial Secession (day) | Total Conidia Collected |
|---|---|---|---|---|
| A | 0.6 | 574 | 38 | 28,910 |
| B | 1.1 | 1034 | 34 | 103,208 |
| C | 0.6 | 594 | 34 | 34,674 |
| D | 1.0 | 957 | 39 | 98,504 |
| E | 0.5 | 498 | 29 | 56,117 |
| F | 0.7 | 650 | 28 | 83,058 |
| Means | 0.8 | 718 | 34 | 67,412 |
| Date | Times of Sunrise | Times of Sunset | Periods of Daytime | Periods of Night-Time |
|---|---|---|---|---|
| 20 February 2021 | 6:37 AM | 5:44 PM | 11 h 07 min | 12 h 53 min |
| 20 May 2021 | 4:50 AM | 6:56 PM | 14 h 06 min | 9 h 54 min |
| 20 July 2021 | 4:58 AM | 7:08 PM | 14 h 10 min | 9 h 50 min |
| 20 October 2021 | 6:06 AM | 5:16 PM | 11 h 10 min | 12 h 50 min |
| 20 November 2021 | 6:35 AM | 4:49 PM | 10 h 14 min | 13 h 46 min |
| 20 December 2021 | 6:59 AM | 4:49 PM | 9 h 50 min | 14 h 10 min |
| Date | The Number of Conidia Collected from a Single Colony at Periods of Daytime * | The Number of Conidia Collected from a Single Colony at Periods of Night-Time * |
|---|---|---|
| 20 February 2021 | 1764 | 27 |
| 20 May 2021 | 7925 | 189 |
| 20 July 2021 | 2438 | 17 |
| 20 October 2021 | 6106 | 216 |
| 20 November 2021 | 2606 | 383 |
| 20 December 2021 | 2606 | 403 |
| Means ± SD | 3794.8 ± 2579.0 a | 205.8 ± 166.3 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ayabe, S.; Kimura, Y.; Umei, N.; Takikawa, Y.; Kakutani, K.; Matsuda, Y.; Nonomura, T. Real-Time Collection of Conidia Released from Living Single Colonies of Podosphaera aphanis on Strawberry Leaves under Natural Conditions with Electrostatic Techniques. Plants 2022, 11, 3453. https://doi.org/10.3390/plants11243453
Ayabe S, Kimura Y, Umei N, Takikawa Y, Kakutani K, Matsuda Y, Nonomura T. Real-Time Collection of Conidia Released from Living Single Colonies of Podosphaera aphanis on Strawberry Leaves under Natural Conditions with Electrostatic Techniques. Plants. 2022; 11(24):3453. https://doi.org/10.3390/plants11243453
Chicago/Turabian StyleAyabe, Shuka, Yutaka Kimura, Naoki Umei, Yoshihiro Takikawa, Koji Kakutani, Yoshinori Matsuda, and Teruo Nonomura. 2022. "Real-Time Collection of Conidia Released from Living Single Colonies of Podosphaera aphanis on Strawberry Leaves under Natural Conditions with Electrostatic Techniques" Plants 11, no. 24: 3453. https://doi.org/10.3390/plants11243453
APA StyleAyabe, S., Kimura, Y., Umei, N., Takikawa, Y., Kakutani, K., Matsuda, Y., & Nonomura, T. (2022). Real-Time Collection of Conidia Released from Living Single Colonies of Podosphaera aphanis on Strawberry Leaves under Natural Conditions with Electrostatic Techniques. Plants, 11(24), 3453. https://doi.org/10.3390/plants11243453